
Abstract
Background:
Thyroid cancer cell lines have been of great value for the study of thyroid cancer. However, the availability of benign thyroid adenoma cell lines is limited.
Methods:
Cell lines were established from thyroid adenomatous nodules that developed in mice treated with the goitrogen amitrole. Expression of epithelial, mesenchymal, and thyroid markers of these established cell lines was determined, and the effect of lentivirus-transduced overexpression of NKX2-1, a master regulator of thyroid development, on the thyroid marker expression was examined. Signal transduction and cell proliferation were evaluated after treatment with insulin-like growth factor-I (IGF-I) and the selective IGF-I receptor (IGF-IR) inhibitor NVP-ADW742. Xenograft studies were performed to examine tumorigenicity of the cells in mice. Whole-genome sequencing (WGS) was used to comprehensively determine the genetic mutations in the established two cell lines.
Results:
Five mouse thyroid adenomatous nodules-derived cell lines named CAT (cells from amitrole-treated thyroids) were established. Among these, two cell lines, CAT458/458s (CAT458s: a subline of CAT458) and CAT459, were found to be positive for epithelial markers and negative for a mesenchymal marker. NKX2-1-positive CAT459 cells showed higher messenger RNA (mRNA) expression of some thyroid differentiation markers than NKX2-1-negative CAT458s cells, and NKX2-1 overexpression increased and/or induced their expression. IGF-I signaling was transduced in thyrotropin receptor (Tshr)-negative CAT458s and 459 cells, and NVP-ADW742 suppressed their proliferation. No tumors developed in mice after subcutaneous injection of CAT458s or 459 cells. The WGS analysis revealed the presence of missense mutations in the tumor suppressor genes such as Polk (encoding DNA polymerase kappa) and Tgfb1 (encoding transforming growth factor beta 1), while no mutations were found in the prominent thyroid cancer-related genes Braf, Trp53 (encoding p53), and Tert (encoding telomerase reverse transcriptase).
Conclusions:
Two mouse thyroid adenomatous nodule-derived cell lines with different thyroid differentiation marker expression were established. NKX2-1 induced partial differentiation of these cell lines. They lacked tumorigenicity and prominent gene mutations involved in thyroid cancer development, while missense mutations were found in some tumor suppressors as revealed by WGS. The CAT458s and 459 provide a new tool to further clarify the process of thyroid multistep carcinogenesis and differentiation.
Introduction
Many gene mutations have been identified in thyroid carcinogenesis. Among them, the Braf mutation is the most common and is thought to directly transform normal thyroid follicular cells to papillary thyroid cancer (PTC) without an adenoma intermediate step. 1 On the contrary, some types of thyroid cancers are known to arise from thyroid adenomas, leading to malignant transition from the benign adenoma, as proposed in the multistep carcinogenesis model. 1
Human thyroid cancer cell lines, including those derived from PTC, follicular thyroid cancer (FTC), and anaplastic thyroid cancer (ATC), have been extensively used to study thyroid cancer. 2 To investigate normal thyroid functions, the spontaneously immortalized normal rat thyroid cell lines, FRTL-53–5 and PCCL3, 5,6 and normal human thyroid cell lines immortalized with SV40, Nthy-ori 3-1 7 and N-thy-Ts.1, 8 have been used. However, thyroid adenoma cell lines have not been established and characterized. Thyroid adenoma cell lines as model benign thyroid epithelial cells would help unravel the mechanisms of thyroid carcinogenesis and thyroid differentiation.
Amitrole, 3-amino-1, 2, 4-triazole, widely used herbicide for nonfood cropland, was reported to inhibit thyroid peroxidase (TPO) and iodine uptake 9 –11 and induce tumors in rat thyroids. 12,13 In addition, amitrole induces thyroid adenomas, but not thyroid cancer in Nkx2-1 (fl/fl) mice. 14
In the present study, cell lines were established from thyroid adenomatous nodules developed in mice fed amitrole-containing diet. Among the five cell lines established, two cell lines showed expression of epithelial and thyroid markers, suggesting that these cell lines originated from thyroid follicular epithelial cells, while the other three cell lines were considered as mesenchymal cells. The differentiation, proliferation, and tumorigenicity capacity of the two thyroid epithelial cell lines were characterized, and whole-genome sequencing (WGS) was carried out to identify gene mutations in these cell lines. The results suggest that they are consistent with benign thyroid cells with different degrees of thyroid differentiation. These cell lines could provide a novel tool to obtain a more complete understanding of thyroid carcinogenesis and differentiation.
Materials and Methods
Generation of thyroid adenomatous nodule-derived cell lines and cell cultures
Method for the generation of CAT (cells from amitrole-treated thyroids) cell lines and culture conditions for CAT and MLg cells purchased from ATCC are described in the Supplementary Methods.
Cultured cell analyses
The lentiviral vector expression system was used to establish CAT458s and 459 cells stably expressing tetracycline (Tet)-inducible rat NKX2-1 protein, as previously described. 15 Methods for production of recombinant lentivirus, selection of infected cells, induction of NKX2-1, cell proliferation assay, CCK-8 assay, and the reagents used in the study are described in the Supplementary Methods.
Biochemical and molecular biological methods
Total RNA extraction, complementary DNA (cDNA) synthesis, and quantitative reverse transcription–polymerase chain reaction (qRT-PCR), and protein extraction and Western blotting are described in the Supplementary Methods. Primers and antibodies used in this study are listed in Supplementary Tables S1 and S2, respectively.
Animal procedures and cytological/histological analysis
All animal studies were performed in accordance with the Using Animals in Intramural Research Guidelines (National Institutes of Health Animal Research Advisory Committee, National Institutes of Health [NIH]) with approval of the Institutional Animal Care and Use Committee (No. 01-094, LM-077). Subcutaneous xenografting of CAT458s and 459 cells in immunocompromised nude mice and preparation of cells and mouse tissues for cytological and histological analyses were performed as described in the Supplementary Methods.
WGS and validation of gene mutations
Genomic DNA extraction, WGS, and validation were performed as described in the Supplementary Methods. Primers used for validation were designed based on the Ensembl 102 GRCm38 database. 16 The sequences are listed in Supplementary Table S1. Forward primers used for validation were used as sequencing primers unless otherwise mentioned.
In silico amino acid mutation analysis
Predicted mutation impacts on stability were estimated using protein structure- 17 –19 and sequence 20 –23 -based approaches. Various methods 24 –30 were employed to predict the association with a disease. The details are described in the Supplementary Methods.
Database analysis
The mutated gene lists by WGS were analyzed using Database for Annotation, Visualization and Integrated Discovery (DAVID) knowledgebase. 31,32 Protein alignment comparison between mouse and human was performed using Clustal Omega, 33,34 and the mutations in cancer patients were analyzed using the database Catalogue Of Somatic Mutations In Cancer (COSMIC). 35 The details are described in the Supplementary Methods and Supplementary Tables S6–S9.
Statistical analysis
Two-tailed Student's t-tests or Welch's t-tests were used for comparison of two samples. Results were considered significant at p < 0.05.
Results
Establishment of five cell lines from thyroid adenomatous nodules induced by amitrole in mice
Nkx2–1(fl/fl) mice were fed an amitrole-containing diet starting at 5 weeks of age to induce thyroid adenomas, as previously described. 14 Mice developed thyroid tumors within 6–12 months post-initiation of the diet (Fig. 1A, B). The thyroid adenomatous nodules, initially characterized based on gross pathology, were resected, whole tumor tissue was dissociated, and the cells were subjected to continuous culture. The nodules were similar to adenomas found in the thyroids of amitrole-treated Nkx2-1(fl/fl);Tpo-Cre mice 14 (Supplementary Fig. S1). The primary cells that continuously grew after five passages were considered as permanent cell lines, and five CAT lines, CAT411, 413, 427, 458, and 459, were successfully established from five individual tumors in individual mice (Fig. 1A).

CAT458/458s and 459 cells show epithelial morphology and epithelial marker expression. (
CAT411, 413, and 427 cells were spindle-shaped cells (Fig. 1C), as was MLg, a mouse lung fibroblast cell line (Supplementary Fig. S2). On the contrary, CAT458, 458s, a subline of 458, and 459 showed cuboidal cell morphology (Fig. 1C), suggesting that they are epithelial cells. In fact, the expression of the epithelial markers, E-cadherin and Claudin-1, was found in CAT458, 458s, 459, and mouse thyroid, but not in 411, 413, 427, and MLg cells (Fig. 1D). In contrast, Vimentin, a mesenchymal marker, was not found in CAT458, 458s, and 459, but expressed in the other CATs, MLg, and in thyroid (Fig. 1D), consistent with cellular morphologies. These results suggest that CAT458/458s and 459 cells originated from epithelial cells, while the other CATs were from mesenchymal cells, presumably residing in the thyroid.
CAT458/458s and 459 cells originate from thyroid epithelial cells
To confirm that CAT458/458s and 459 epithelial cells originated from thyroids, the expression of thyroid transcription factors critical for thyroid development and function 36,37 was first examined. Nkx2-1, Pax8, and Foxe1 messenger RNAs (mRNAs) were found in CAT458/458s (Nkx2-1 not detected in CAT458 cells) and CAT459 cells, while Nkx2-1 mRNA was not detected in all other CATs and MLg cells (Fig. 2A). It was noted that NKX2-1 protein, a master regulator of thyroid development, 36,38 was expressed in CAT459 cells at comparable levels as found in mouse thyroid (Fig. 2B). On the contrary, PAX8 protein was detected in only mouse thyroid (Fig. 2B).
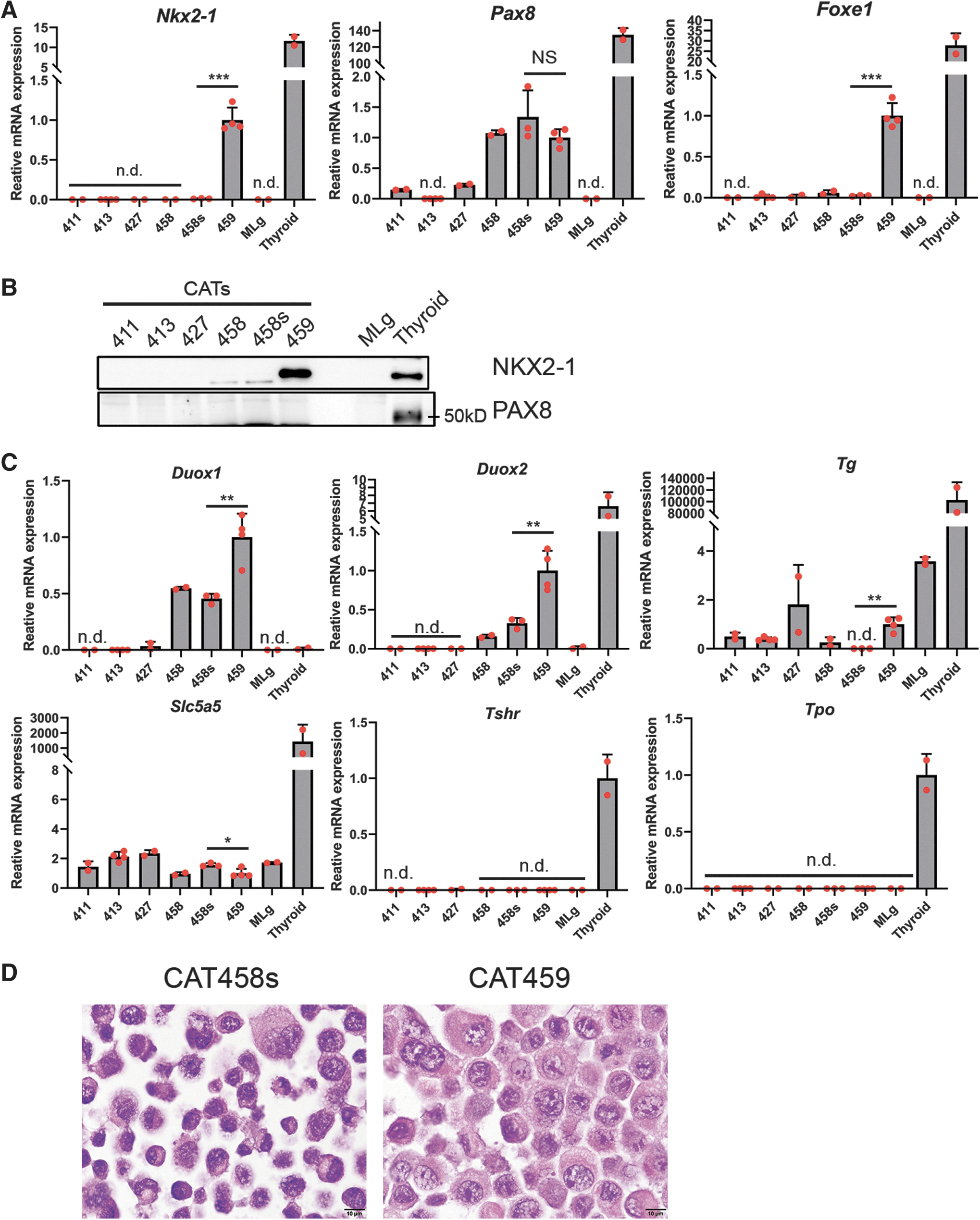
CAT458/458s and 459 cells express thyroid transcription factors and thyroid markers. (
Next, mRNAs encoding thyroid markers 37 were quantified. Both Duox1 and Duox2, NADPH oxidases involved in thyroid hormone synthesis, were expressed in CAT458/458s and 459 cells (Fig. 2C). Notably, Duox1 mRNA was much higher in CAT458/458s and 459 cells than in the mouse thyroid (Fig. 2C) due to an unknown mechanism. However, Tg encoding thyroglobulin, a main precursor of thyroid hormones, and Slc5a5, encoding sodium iodide symporter (NIS), were barely expressed in CAT458/458s (Tg not detected in CAT458s cells) and CAT459 cells (Fig. 2C). Furthermore, no expression of thyrotropin receptor (Tshr) and Tpo mRNAs was detected in either CAT458/458s or 459 cells (Fig. 2C), as reported in human thyroid cancer cell lines. 39 Collectively, these results suggest that CAT458/458s and 459 cells originated from thyroid follicular epithelial cells.
NKX2-1-positive CAT459 cells showed higher expression of thyroid marker mRNAs including Foxe1, Duox1, Duox2, and Tg than the NKX2-1-negative CAT458s cells, except for a slightly lower expression of Slc5a5 mRNA (Fig. 2A, C). Consistent with this finding, Hematoxylin & Eosin staining revealed that CAT458s cells showed a higher degree of cellular atypia than CAT459 cells, and CAT459 cells had more cytoplasm than CAT458s cells (Fig. 2D). These results suggest that NKX2-1-positive CAT459 cells are more differentiated.
NKX2-1 induces partial thyroid differentiation in CAT458s and 459 cells
Next, to examine the effect of NKX2-1 on the expression of thyroid differentiation markers, CAT458s and 459 cells stably expressing Tet-inducible rat NKX2-1, named 458s- and 459-TNKX cells, respectively, were established (Fig. 3A). Overexpression of NKX2-1 under doxycycline (DOX) was confirmed in both cell lines (Fig. 3B, C). Enlargement of cells was found 3 days after DOX treatment in both cell lines (Fig. 3D), suggesting that NKX2-1 may induce thyroid differentiation. Furthermore, the expression of Pax8 and thyroid markers including Duox1, Duox2, Tg, and Tshr mRNAs was higher in both 458s- and 459-TNKX cells with NKX2-1 overexpression (Fig. 3E). On the contrary, Foxe1, Slc5a5, and Tpo mRNAs were not induced by NKX2-1 (Supplementary Fig. S3). These results suggest that NKX2-1 can induce and further enhance partial thyroid differentiation in CAT458s and 459 cells.
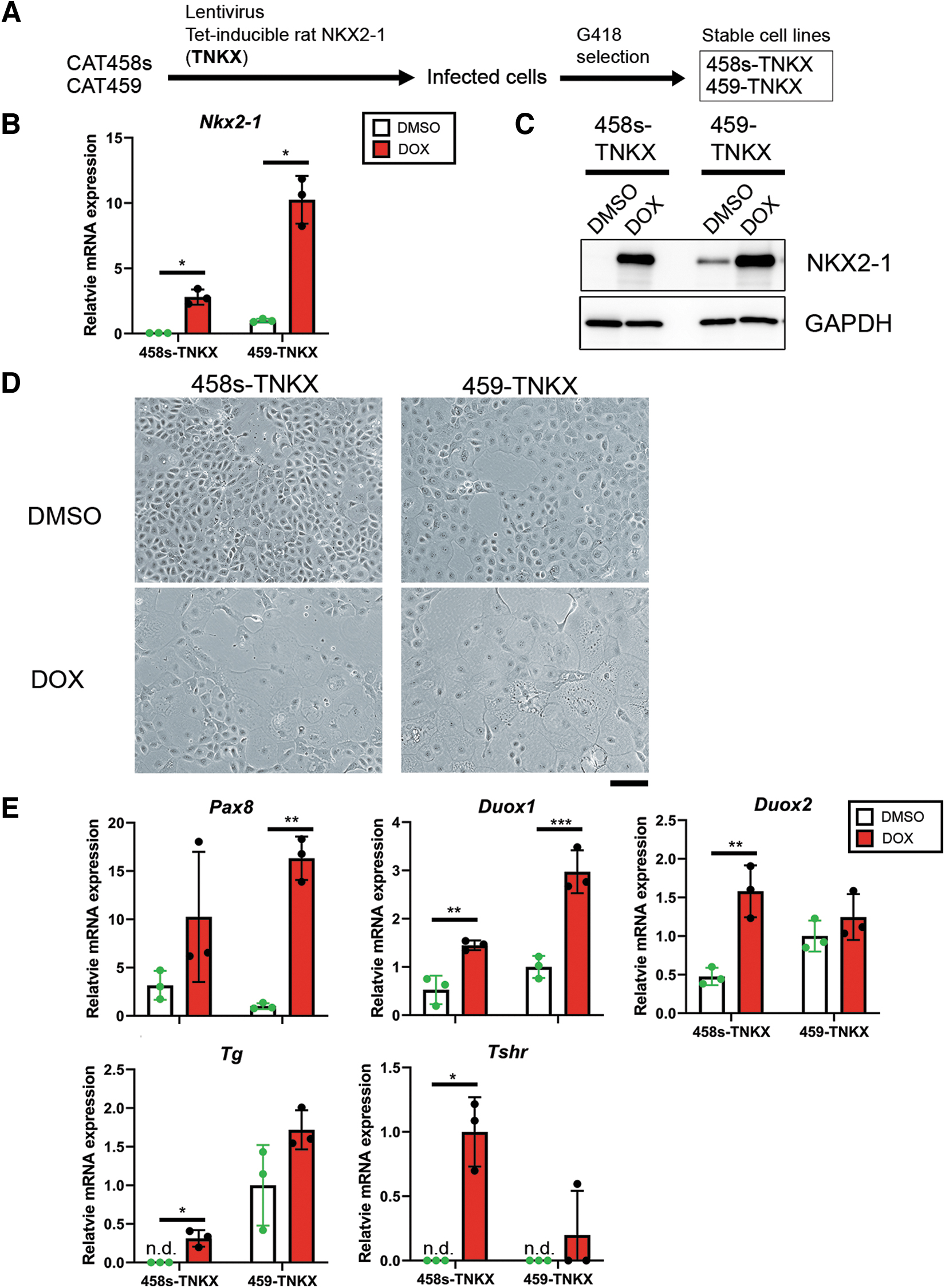
NKX2-1 induces thyroid marker expression in CAT458s and 459 cells. (
Insulin-like growth factor-I signaling is partly responsible for the growth of Tshr-negative CAT458s and 459 cells
It is known that thyrotropin (TSH) signals through the TSHR and is essential for growth and differentiation of thyroid epithelium. 40 However, Tshr expression is deficient in mouse thyroid adenomatous nodule-derived CAT458s and 459 cells (Fig. 2C), as reported for human thyroid cancer cells. 39 Studies using FRTL-5 cells demonstrated that insulin-like growth factor-I (IGF-I) works synergistically with TSH in thyroid follicular epithelium. 41,42 Hence, studies were conducted to determine whether IGF-I signaling is transduced independent of TSHR and is required for the growth of Tshr-negative CAT458s and 459 cells.
Igf1r (encoding IGF-I receptor) mRNA and IGF-I receptor β (IGF-IRβ) protein expression was found in CAT458s and 459 cells (Supplementary Fig. S4A and Fig. 4A), while Igf1 mRNA levels were barely detected (Supplementary Fig. S4B). Induction of phosphorylation of downstream targets of IGF-I signal, AKT and S6 ribosomal protein, 40 was confirmed in both CAT458s and 459 cells after IGF-I treatment (Fig. 4A). Proliferation of CAT459 cells was promoted by IGF-I; however, IGF-I treatment failed to promote the growth of CAT458s cells (Fig. 4B and Supplementary Fig. S5A). Whether endogenous IGF-IR-mediated signaling affects the growth of these cells was next examined using NVP-ADW742 (ADW742), a selective IGF-IR inhibitor. 43 ADW742 inhibited the proliferation of both cell lines (Fig. 4C and Supplementary Fig. S5B). These results suggest that IGF-I signaling is transduced and at least partly responsible for the growth of Tshr-negative CAT458s and 459 thyroid cells.
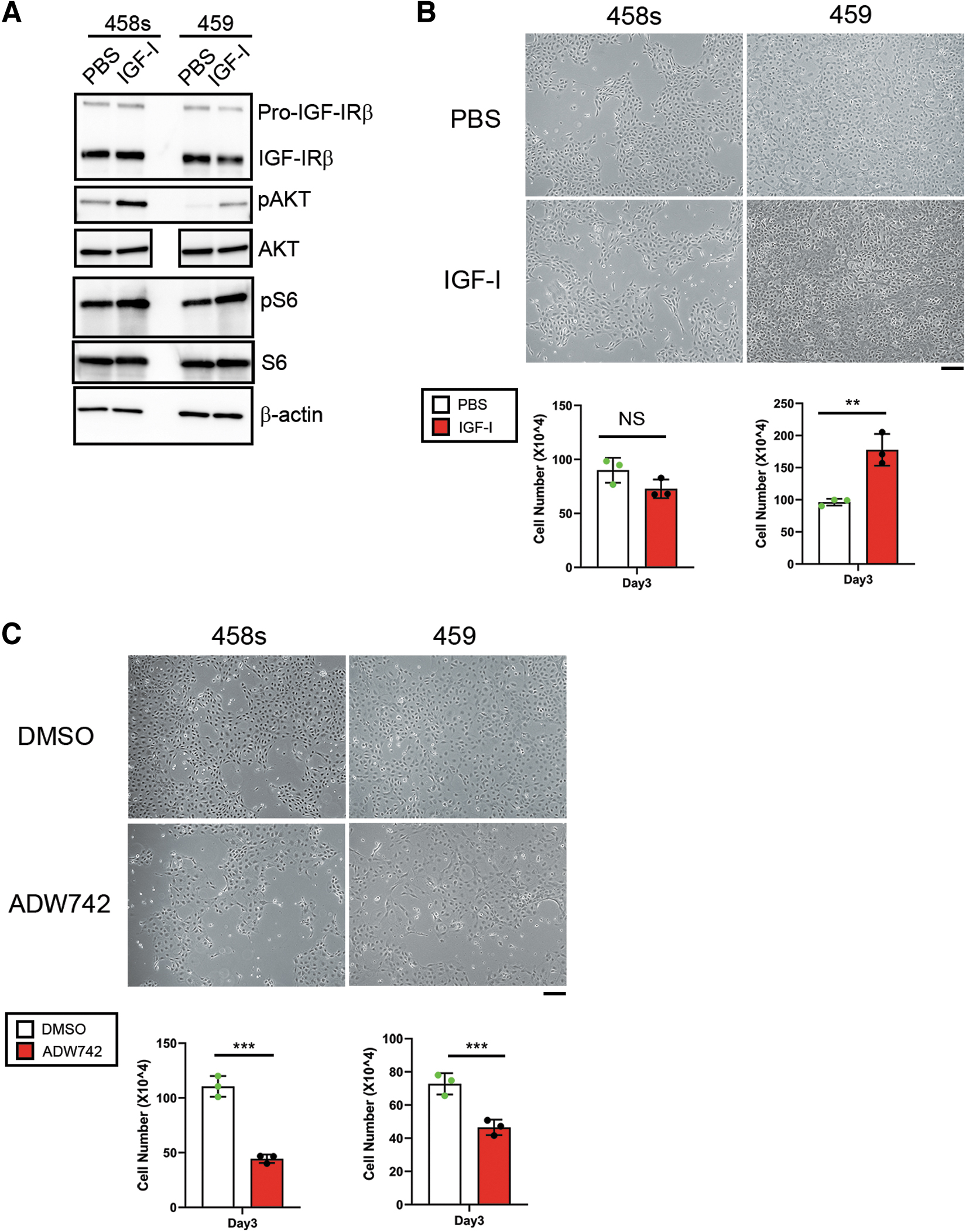
IGF-I signaling is partly responsible for the proliferation of Tshr-negative CAT458s and 459 cells. (
CAT458s and 459 cells are not tumorigenic in vivo
Since whole tumor tissues were used to establish CAT458s and 459 cell lines, there was a possibility that a small number of cancer cells that resided in the tumor might have been selected and formed the resultant cell lines. Whether CAT458s and 459 cells are tumorigenic in vivo was therefore examined by subcutaneous injection of these cells into immunocompromised nude mice. Tumors did not develop in mice when examined 4–8 weeks after injection (Table 1). Indeed, viable injected cells were not observed in subcutaneous tissues 6 days after injection (Supplementary Fig. S6). These results suggest that CAT458s and 459 cells are consistent with benign thyroid cells, different from thyroid cancer cells.
Tumorigenicity of CAT458s and 459 Cells in Nude Mice
1 or 3 × 106 Cells of CAT458s or 459 cells were subcutaneously implanted into nude mice. Incidence of developed growing subcutaneous tumors 4–8 weeks after implantation was evaluated by gross examination (shown as developed growing tumor/mouse).
A tiny fibrous nodule (containing possible epithelial cells, examined by H&E staining, figure not shown) remained in one mouse even 8 weeks after injection of 3 × 106 of CAT459 cells.
CAT, cells from amitrole-treated thyroids; H&E, Hematoxylin & Eosin.
WGS reveals missense gene mutations in tumor suppressor genes in CAT458s and 459 cells
Finally, WGS was carried out to identify somatic gene mutations present in CAT458s and 459 cells. Analysis for structural variations (SV) was first carried out, which showed that both cells have similar relative prevalence among the variations, although CAT459 cells have more SV than 458s cells (Supplementary Fig. S7). Next, missense gene mutations of CAT458s and 459 cells were examined. Since CAT cell lines were derived from Nkx2-1(fl/fl) mice on a mixed genetic background, 14 all the normal variants reported in Ensembl 102 and mgp v5 dbSNP142 were excluded from the mutation lists of CAT458s and 459 cells (Fig. 5A). This resulted in the identification of 65 CAT458s-specifc and 53 CAT459-specifc mutated genes (Fig. 5B and Supplementary Tables S3 and S4).
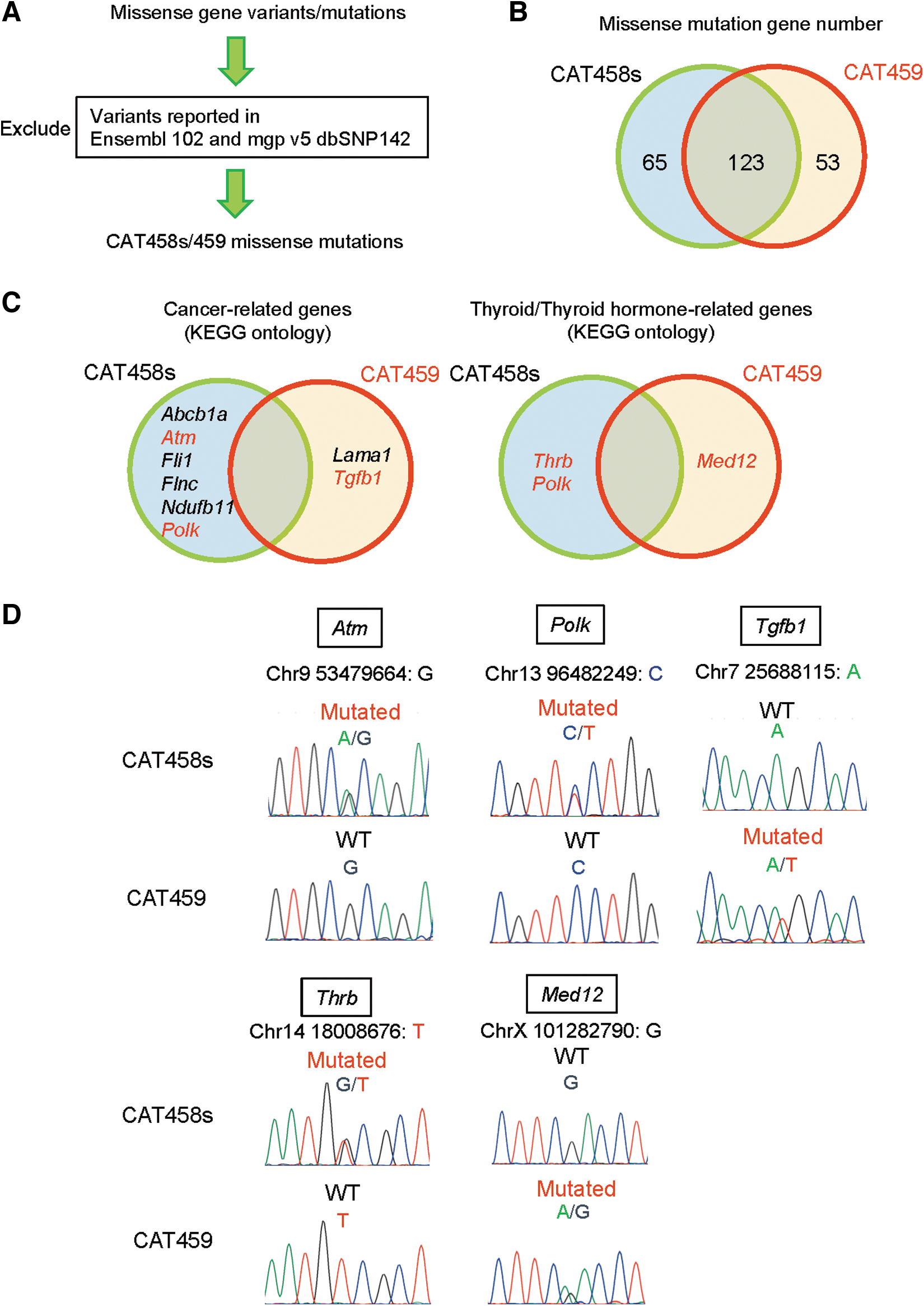
WGS reveals missense mutations in tumor suppressor genes in CAT458s and 459 cells. (
From each 65 and 53 pools of genes for the CAT458 and CAT459 cells, respectively, “Cancer-related genes” and “Thyroid/thyroid hormone-related genes” were extracted (see the Supplementary Methods). Among “Cancer-related genes,” prominent gene mutations known to be involved in thyroid cancer, such as Braf, Trp53 (encoding p53), and Tert (encoding telomerase reverse transcriptase), 1,44,45 were not identified, suggesting that CAT458s and 459 cells are consistent with benign thyroid adenoma cells. Indeed, most of human cancer cell lines possess missense gene mutations in these genes. 46,47 Instead, missense mutations in tumor suppressor genes Atm (encoding ataxia telangiectasia mutated) 48 and Polk (encoding DNA polymerase kappa) 49 in CAT458s, and Tgfb1 (encoding transforming growth factor beta 1) 50 in CAT459 were found (Fig. 5C).
Possible tumor suppressors Thrb (encoding thyroid hormone receptor β) 51,52 and Med12 (encoding mediator complex subunit 12) 53 –55 were additionally found in the CAT458s- and 459-specific mutated gene lists, respectively, in “Thyroid/thyroid hormone-related genes” (Fig. 5C). Therefore, these five genes were selected, and their mutations were validated by PCR amplification followed by sequencing (Fig. 5D and see the mutations details in Supplementary Tables S3 and S4). It was also confirmed that these mutations are absent in CAT411, 413, and 427 cells (Supplementary Fig. S8).
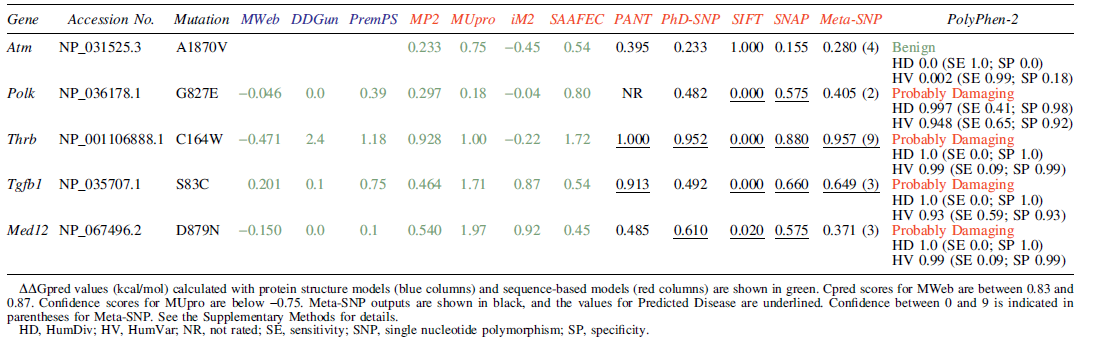
In silico analysis of these mutations in tumor suppressors was carried out to predict protein stability and association with a disease. Predicted stability change by the mutations showed a tendency toward a slight protein instability, suggesting that the mutations might lead to loss of function (Table 2). Meta-Single Nucleotide Polymorphism results with various frequently used methods 24 –28 showed the possibility that the mutations except for Atm are associated with a disease, especially Thrb with a higher likelihood (Table 2). Predictions with PolyPhen-229 also suggested that these mutations may cause disease (Table 2). In addition, predictions of the corresponding mutations in human with MutationTaster202130 also suggested that the mutations in THRB and TGFB1 are deleterious (Supplementary Table S5). These predictions are consistent with the possibility that the mutations found in CAT458s and 459 cells contribute to thyroid adenoma development.
In Silico Analysis of Amino Acid Mutations in Tumor Suppressor Genes in CAT458s and 459 Cells
Discussion
Five cell lines were established from amitrole-induced mouse thyroid adenomatous nodules, among which the CAT458/458s and 459 cell lines possess thyroid epithelial properties. CAT459 cells positive for thyroid master transcription factor, NKX2-1, showed higher expression of thyroid markers than CAT458s cells. In addition, NKX2-1 overexpression induced the expression of some thyroid markers in both cell lines, suggesting that NKX2-1 can induce partial differentiation in these cells, as was reported in mouse embryonic stem (ES) cells. 36 It should be noted that Foxe1 expression is much higher in CAT459 cells than in CAT458s cells. FOXE1 is another essential transcription factor for human thyroid development, 37,56 and FOXE1 expression is induced by NKX2-1 and PAX8 in mouse ES cells. 36 However, in the current study, NKX2-1 overexpression did not induce Foxe1 expression in either cells, suggesting that Foxe1 expression may be regulated independent of NKX2-1. Further studies are necessary to establish the function and regulation of FOXE1 in adult mouse thyroid cell differentiation.
High expression of TSHR, TPO, and NIS is required for thyroid hormone synthesis through TSHR signaling in thyroids. The rat normal thyroid cell lines FRTL-5 and PCCL-3 maintain thyroglobulin production and DNA synthesis in response to TSH under two-dimensional culltures, 3,5 and thus, they are commonly used for normal thyroid function research.
On the contrary, CAT458s and 459 cells barely express or lack Tg, Scl5a5, Tpo, and Tshr mRNA expression. Similar findings were reported in studies using various types of human thyroid cancer cell lines and transformed rat thyroid cell lines. 5,39,47 These findings suggest that CAT458s and 459 cells possess characteristics close to cancer cells that can grow in the absence of TSH. Indeed, bovine TSH (bTSH) treatment did not induce levels of Tg, Slc5a5, and Tpo mRNAs (Supplementary Fig. S9), known target genes associated with TSH signaling. 57 Instead, IGF-I signaling is partly responsible for the growth of Tshr-negative CAT458s and 459 cells. Igf1 mRNA expression is abundant in CAT411, 413, and 427 mesenchymal cell lines, suggesting the possibility that thyroid adenoma-associated fibroblasts could be a source of IGF-I.
Mouse and human thyroid cancer cell lines are known to develop tumor after subcutaneous, intravenous, and orthotopic injection into immunocompromised mice. 47,58,59 When CAT458s and 459 cells were subcutaneously injected into nude mice, small subcutaneous nodules were observed in some mice 6 days after injection; however, only fibrosis and/or inflammatory cells with or without nonviable injected cells were found histologically. Consistent with this, subcutaneous nodules disappeared by 4–8 weeks after injection, except for one tiny fibrous nodule without any growing tumors, suggesting that CAT458s and 459 cells are most likely different from cancer cells. Since CAT458s and 459 cells were derived from thyroid adenomas induced by amitrole treatment, which is known to inhibit iodide uptake as well as TPO, these cells may be useful for the study of iodine deficiency-induced thyroid adenomatous nodules in humans. 60 However, amitrole may have other unknown effects on the thyroid through inhibition of TPO activity or unknown mechanisms, which might limit the use of these cell lines to understand the nature and/or mechanism of thyroid carcinogenesis.
SV are generally thought to cause malignant transformation resulting in cancer, rather than benign adenoma. Thus, missense gene mutations of CAT458s and 459 cells were assessed to obtain insights into the mutations that occur in the early stages of multistep thyroid carcinogenesis. Besides 65 CAT458s-specific and 53 CAT459-specific mutated genes, 123 commonly mutated genes were obtained. Among them, Actl6b (actin-like 6b), Ptprj (protein tyrosine phosphatase receptor type J), Rxrb (retinoid X receptor beta), and Tnxb (tenascin XB) were categorized as “Cancer-related genes” and Atf6b (activating transcription factor 6 beta) and Rxrb as “Thyroid/thyroid hormone-related genes.” However, it is unlikely that the two independent cell lines share mutated genes at the same locus. It is most likely that they are variants derived from the mouse genetic background that were not listed in the public databases used. Indeed, the presence of heterozygous and/or homozygous mutations in Rxrb 61 and tumor suppressor genes Actl6b 62 and Ptprj 63 was confirmed in all five CAT cell lines (data not shown). Therefore, only CAT458s- or 459-spcific missense mutations were studied.
Tshr and Gnas1 (encoding Gsα) point mutations were reported in autonomously functioning thyroid nodules. 1,60,64,65 However, these gene mutations were not found in CAT458s and 459 cells by WGS. It is likely that CAT458s and 459 cells were derived from follicular thyroid adenomas, which could be precancerous. The mutated amino acids in coding proteins due to missense mutations of tumor suppressor genes such as Polk and Thrb in CAT458s and Tgfb1 and Med12 in CAT459 cells are conserved between mice and humans (Supplementary Fig. S10A).
COSMIC database 35 analysis for the presence of missense mutations of these four genes in human cancer patients identified the same G845E mutation in POLK in one glioma patient, 66 G845V in POLK in one lung squamous cell cancer patient, 67 and S83I in TGFB1 in one enodmetrioid carcinoma patient in The Cancer Genome Atlas (TCGA) study (Supplementary Fig. S10B). These findings suggest that these mutations might be involved in early carcinogenesis. On the contrary, the corresponding mutations in THRB and MED12 were not found in the COSMIC database. The Thrb mutation found in CAT458s cells is located in the DNA-binding domain, different from that in the ligand-binding domain observed in human thyroid retinal hormone resistance syndrome (RTH) patients, which caused spontaneous FTC when introduced in mice. 51,52,68
The D878N mutation in MED12 is located in exon 19 (out of 45, middle region). On the contrary, the G44 located in exon 2 of MED12 is one of the hot spot mutation sites observed in some types of malignant tumors such as fatal forms of non-ATC. 53 –55 The G44 is involved in binding to Cyclin C/Cyclin-dependent kinase (CDK) 8, and the mutations at this site were reported to abrogate the activity of CDK8, and the dysregulation of CDK8 possibly contributes to malignancy. 53 –55 These findings suggest that the mutations in Thrb and Med12 we identified may not be potent enough to transform benign thyroid cells. Further studies would be required for clarification of thyroid multistep carcinogenesis.
In conclusion, two mouse thyroid adenomatous nodule-derived cell lines with different thyroid differentiation marker expression were established that possess missense mutations in tumor suppressor genes. However, they lack tumorigenicity in immunocompromised mice, suggesting that they are consistent with benign adenoma cells. CAT458s and 459 cells would provide a tool to further understand the nature and/or mechanism of thyroid carcinogenesis and differentiation.
Footnotes
Acknowledgments
Mice under amitrole diet were maintained at the animal facility run by the National Cancer Institute (NCI) Laboratory Animal Sciences Program. WGS and Sanger sequencing were conducted at Sequencing Facility and the Center for Cancer Research (CCR) Genomics Core, respectively, at the NCI, NIH.
Authors' Contributions
Y.-T.S.: Formal analysis, investigation, writing—original draft, writing—review and editing. N.H.: Investigation, writing—review and editing. J.M.W.: Formal analysis and writing—review and editing. H.L.: Formal analysis and writing—review and editing. R.E.C.: Formal analysis and writing—review and editing. M.P.L.: Formal analysis and writing—review and editing. S.K.: Conceptualization, supervision, writing—review and editing.
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
This work was supported by an Intramural Research Program of the National Cancer Institute, Center for Cancer Research (ZIA BC 005522).
Supplementary Material
Supplementary Methods
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
Supplementary Table S9