
Abstract
Background:
Tanycytes are specialized glial cells within the mediobasal hypothalamus that have multiple functions, including hormone sensing and regulation of hypophysiotropic hormone secretion. There are ongoing discussions about the role of tanycytes in regulating the supply of hypothalamic thyroid hormones (THs) through the expression of TH transporters (Slc16a2, Slco1c1) and deiodinases (Dio2, Dio3). In this study, we investigated the potential feedback effect of thyrotropin (TSH) on the transcription of these gatekeeper genes on tanycytes.
Methods:
We analyzed the changes in the expression of TH-gatekeeper genes, in TSH-stimulated primary tanycytes, using quantitative polymerase chain reaction (qPCR). We also used RNAScope® in brain slices to further reveal the local distribution of the transcripts. In addition, we blocked intracellular pathways and used small-interfering RNA (siRNA) to elucidate differences in the regulation of the gatekeeper genes.
Results:
TSH elevated messenger RNA (mRNA) levels of Slco1c1, Dio2, and Dio3 in tanycytes, while Slc16a2 was mostly unaffected. Blockade and knockdown of the TSH receptor (TSHR) and antagonization of cAMP response element-binding protein (CREB) clearly abolished the increased expression induced by TSH, indicating PKA-dependent regulation through the TSHR. The TSH-dependent expression of Dio3 and Slco1c1 was also regulated by protein kinase C (PKC), and in case of Dio3, also by extracellular signal-regulated kinase (ERK) activity. Importantly, these gene regulations were specifically found in different subpopulations of tanycytes.
Conclusions:
This study demonstrates that TSH induces transcriptional regulation of TH-gatekeeper genes in tanycytes through the Tshr/Gαq/PKC pathway, in parallel to the Tshr/Gαs/PKA/CREB pathway. These differential actions of TSH on tanycytic subpopulations appear to be important for coordinating the supply of TH to the hypothalamus and aid its functions.
Introduction
Thyroid hormones (THs) are important during the perinatal period for proper neuronal development. 1 In adults, THs play an essential role in heart function, 2 bone formation, 3 fertility, 4 and energy homeostasis through central and peripheral mechanisms. 5,6 To keep the TH levels constant, pathological changes of THs are counter regulated by central feedback mechanisms, adapting thyrotropin-releasing hormone (TRH) and thyrotropin (TSH) synthesis and release. The underlying feedback mechanisms involve not only the transport of THs across the blood–brain barrier (BBB) through specific transporters (MCT8 encoded by Slc16a2, OATP1C1 encoded by Slco1c1) but also the conversion of inactive thyroxine (T4) into active triiodothyronine (T3). 7 The brain-specific conversion of THs is mediated by type 2 deiodinase (DIO2) and type 3 deiodinase (DIO3). DIO2 primarily converts T4 to T3. On the contrary, DIO3 converts T4 into the inactive reverse T3 and T3 into the inactive T2, effectively inactivating THs.
A peripheral decrease in TH levels leads to an increase in the expression of Dio2 in astrocytes. 8 The function of DIO2 in maintaining T3 supply to the brain is supported by the finding that global Dio2 knockout mice have lower T3 levels in the brain, and elevated Trh messenger RNA (mRNA) and TSH plasma levels, similar to hypothyroid mice. 7 However, the astrocytic Dio2 knockout failed to affect the DIO2 activity in the hypothalamus, in contrast to other brain areas, and the hypothalamic-pituitary-thyroid (HPT) axis, which indicates a special regulation of the TH supply to the hypothalamus. 9
In the hypothalamus, Slc16a2, Dio2, and other TH-gatekeeper genes are expressed primarily in tanycytes. 10,11 Tanycytes are glial cells that have long processes extending into hypothalamic nuclei and the median eminence (ME), where they come into close contact with blood vessels and neuronal terminals. 12,13 They regulate the entry of peripheral substances 14 –18 and control the release of hypophysiotropic hormones, including TRH and gonadotropin releasing hormone. 19 –21 Therefore, tanycytes are crucial for the regulation of HPT and gonadal axes. 19 –22
There is limited evidence available to support the hypothesis that peripheral hormones of the HPT axis (TSH and THs) modulate tanycytic functions in controlling the endocrine axis by regulating the expression of TH-gatekeeper genes. Indeed, hypothyroidism increases Dio2 expression in the hypothalamus, but a direct inhibition of Dio2 transcription by TH is unlikely. 23 As an alternative explanation, TSH may induce Dio2 mRNA. 24 Whether TSH directly modulates tanycytic TH gatekeepers has not been investigated. Therefore, in this study, we investigated the transcriptional regulation of Slc16a2, Slco1c1, Dio2, and Dio3 in tanycytes by TSH, and elucidated the involved intracellular pathways activated by TSH receptor (TSHR) using small-interfering RNA (siRNA) knockdown and pharmacological blockers. Our data provide the first evidence for a direct effect of TSH on tanycytic gatekeeper functions and elucidate different reactions of tanycytic subpopulations.
Methods
Animals
All national and institutional guidelines for the care and use of laboratory animals have been followed and have been approved by the local regulatory authority (Ministerium für Landwirtschaft, ländliche Räume, Europa und Verbraucherschutz, and University of Lübeck) with the proposal numbers: Az.: 60-7-18 and 22_2022-03-29_Müller-Fielitz. Sprague-Dawley rats and C57Bl/6N mice were bred in the animal facility, University of Lübeck. All animals were kept at a 12 hour:12 hour light:dark cycle with ad libitum access to food and water. Tanycytes were isolated from p10 Sprague-Dawley rats by dissecting the wall of the third ventricle of the mediobasal hypothalamus and ME as described previously. 25
AAV injection
The original plasmid containing Flamindo2 was a gift from Tetsuya Kitaguchi (No. 73938; Addgene). The ORF of Flamindo2 was subcloned in an AAV-vector. The AAV-CAG-Flamindo2 was generated with a mosaic capsid of serotypes 1 and 2, and purified by affinity chromatography as described before. 19 For stereotaxic injections, mice were anesthetized with ketamine (65 μg/g, i.p.) and xylazine (14 μg/g, i.p.), and fixed in the stereotaxic frame (Kopf Instruments) as described previously. 19 AAV (109 gp) was injected into the lateral ventricle (anteroposterior −0.1 mm, mediolateral −0.7 mm, and dorsoventral −2.3 mm from bregma). Two weeks after AAV injection, the brains were dissected for ex vivo cAMP measurement.
cAMP and Ca2+ measurements
As described previously, 19,26 brain slices (200 μm) were prepared using a vibrating blade microtome (Leica VT1200 S) and loaded, for [Ca2+]i measurements, with Fura2-AM. Changes in tanycytic [cAMP]i and [Ca2+]i were measured in a flow chamber, perfused with artificial cerebrospinal fluid, using an imaging setup (Till Photonics) mounted on the Axio Examiner D1 upright fluorescent microscope (Zeiss). 19
Ex vivo stimulation
Hypothalamic blocks were placed in neurobasal medium (neurobasal medium, 10% charcoal stripped fetal bovine serum [FBS], 2% B27, 0.5% glutamax, 0.5% penicillin/streptomycin), and stimulated with 60 mIU/mL TSH or phosphate-buffered saline (PBS) for 4 hours, fixed with 4% PFA overnight, and 50 μm slices were prepared for in situ hybridization.
Pituitaries were incubated, after dissection, in 50 μL PBS with or without taltirelin (1 μM) for 4 hours. The supernatants were used to treat primary tanycytes.
Primary tanycytes treatment
Primary tanycytes were seeded into 24-well plates and were starved (Dulbecco's modified Eagle's medium [DMEM]/F12 without phenol red, 0.15% insulin, and 0.3% putrescin) for 6 hours. Then, they were simulated with 20 μL supernatant from pituitaries or bovine TSH (60 mIU/mL) in medium (DMEM high glucose, 10% charcoal-stripped FBS, 2 nM L-glutamine, and 1% penicillin/streptomycin) for 12 hours.
Tanycytes were also treated with 50 μM S37a (TSHR-antagonist), 5 μM 666-15 (cAMP response element-binding protein [CREB]-antagonist), 10 μM bisindolylmaleimide-I (protein kinase C [PKC]-antagonist), or 50 μM FR180204 (extracellular signal-regulated kinase [ERK]-antagonist) along with TSH. Vehicle-treated cells were used as controls. For siRNA knockdown, tanycytes (80% confluence) were transfected with 5 μM siRNA against Tshr or scramble using Lipofectamine RNAiMAX transfection reagent, as described by the manufacturer. Sixteen hours after transfection, the cells were starved for 6 hours and treated as described.
Quantitative polymerase chain reaction
mRNA was isolated from primary tanycytes using NucleoSpin RNA Kit, and RNA was transcribed using Moloney-Murine-Leukemia-Virus, Reverse-Transcriptase, and random-hexamer primers (Promega). Quantitative polymerase chain reaction (qPCR) was performed with the FastStart Essential DNA Green Master using a three-step amplification on a Roche LightCycler as described before. 27
In situ hybridization
The in situ hybridization for Slc16a2, Slco1c1, Dio2, Dio3, Thra1 (encoding THRα2), Thrb, and Tshr mRNA was performed using RNAScope® Multiplex Fluorescent Reagent Kit v2 (ACD). 25 After in situ hybridization, immunohistochemistry for MCT8 was performed. Brain slices were imaged using confocal microscope Leica TCS SP5. A blinding strategy was followed while scoring the RNAScope images.
Statistics
Data were analyzed using GraphPad Prism 8. Significance was considered when p < 0.05. Parametric statistics (one-way or two-way analysis of variance) was applied only if the assumptions were met; that is, datasets were examined for Gaussian distribution using the D'Agostino-Pearson test and homogeneity of variances by Brown-Forsythe, Levene's or F-test (depending on the statistical method). If assumptions for parametric procedures were not met, nonparametric methods were used as indicated.
Detailed methods are described in the Supplementary Data.
Results
We validated the basal expression of the TH-gatekeeper genes Slc16a2 (Fig. 1A), Slco1c1 (Fig. 1B), Dio2 (Fig. 1C), and Dio3 (Fig. 1D), the TH-receptors (THRs) Thra1 (Fig. 1E; encoding THRα2) and Thrb (Fig. 1F), and the TSH receptor (Fig. 1G; Tshr) in the ependymal layer of the basal third ventricle of mice using RNAScope. The basal expression of Dio3 mRNA was quite low compared with Dio2. Tshr mRNA was expressed in the same cells as Dio2 (Fig. 1H) and Slco1c1 (Fig. 1I). The positive cells of the ependymal layer were identified as tanycytes based on their location and specific morphology, visualized by immunohistochemically staining for MCT8.

Localization of Slc16a2 (
To test whether TSH released from the pituitary mediates changes in tanycytic gene expression, we used supernatant from pituitaries stimulated ex vivo with taltirelin, a TRH-receptor agonist. Supernatant from vehicle-treated pituitaries (PitPBS) increased Slco1c1 (Fig. 2B) and Dio3 (Fig. 2D) but did not affect Slc16a2 and Dio2 mRNA levels (Fig. 2A, C). Supernatant from taltirelin-stimulated pituitaries (PitTalti) elevated expression of Slco1c1 (Fig. 2B), Dio2 (Fig. 2C), and Dio3 mRNA levels (Fig. 2D). All three genes were not induced by taltirelin alone, indicating that a TRH-dependent factor, from the pituitary, stimulates gene transcription in tanycytes.

Transcriptional regulation of Slc16a2 (
To test whether TSH qualifies as the pituitary-released factor regulating the expression of TH gatekeepers in tanycytes, we stimulated primary tanycytes with TSH. In addition, tanycytes were treated with the selective TSHR-antagonist S37a 28 or were transfected with siRNA targeting Tshr (siTshr) to knock down Tshr expression. Stimulation with TSH resulted in no change in Slc16a2 (Fig. 3A, F), and an increase in Slco1c1 (Fig. 3B, G), Dio2 (Fig. 3C, H), and Dio3 mRNA levels (Fig. 3D, I). Incubation with S37a or siTshr, which reduced the Tshr mRNA by 60% (Fig. 3E), diminished or abolished the effects of TSH on Slco1c1 (Fig. 3B, G), Dio2 (Fig. 3C, H), and Dio3 mRNA levels (Fig. 3D, I). This demonstrated a TSH-dependent regulation of TH-gatekeeper genes through TSHR.
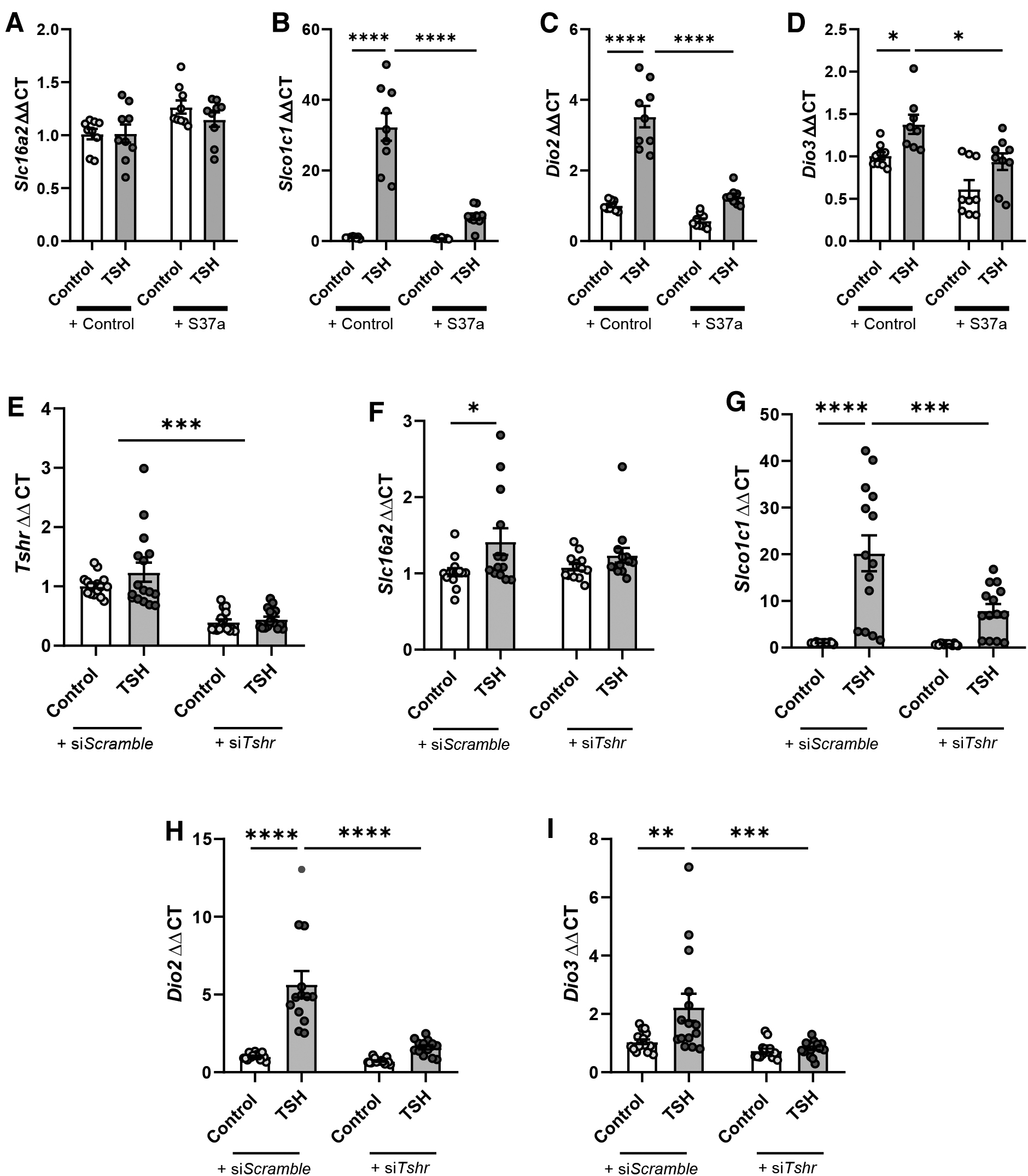
Transcriptional regulation of Slc16a2 (
Tanycytes represent a heterogeneous population of cells with specific properties depending on their anatomical location. 12,29 Since tanycytic subpopulations cannot be distinguished in primary cell culture, we explored the effects of TSH-mediated tanycytic gene regulation on acute brain slices using RNAScope. TSH treatment clearly increased Slco1c1 (Fig. 4B), Dio2 (Fig. 4C), and Dio3 (Fig. 4D) expression compared with controls.

Effect of TSH (TSH; 60 mIU/mL) on mRNA expression levels of Slc16a2 (
Congruent with primary tanycytes, a distinct regulation of gene expression for Slc16a2 mRNA was not apparent (Fig. 4A). Interestingly, the signal for Dio2 mRNA was only increased in tanycytes projecting to the ME (Fig. 4C). The expression of Dio3, however, was only increased in tanycytes on the wall of the third ventricle (Fig. 4D). This indicates that the tanycytic subpopulations differentially regulate TH-gatekeeper genes in response to TSH.
TSHR is a seven transmembrane receptor (GPCR) known to be coupled to Gαs signaling and is involved in cAMP production. This coupling was verified in mice by transducing tanycytes with the citrine-based cAMP sensor, Flamindo2. The fluorescence of Flamindo2 is reduced on binding cAMP. Two weeks after transduction of tanycytes, bath application of TSH on acute brain slices led to reduction of Flamindo2 signal (Fig. 5A, B), indicating an increased [cAMP]i (−5.4% ± 1.1%). Stimulation with forskolin, a stimulus of cAMP production, also reduced fluorescence in tanycytes (−6.4% ± 0.98%, Fig. 5C, D). The TSH-mediated increase in [cAMP]i was confirmed in primary tanycytes, using a cAMP assay, which could be antagonized by the TSHR-antagonist S37a (Fig. 5E).
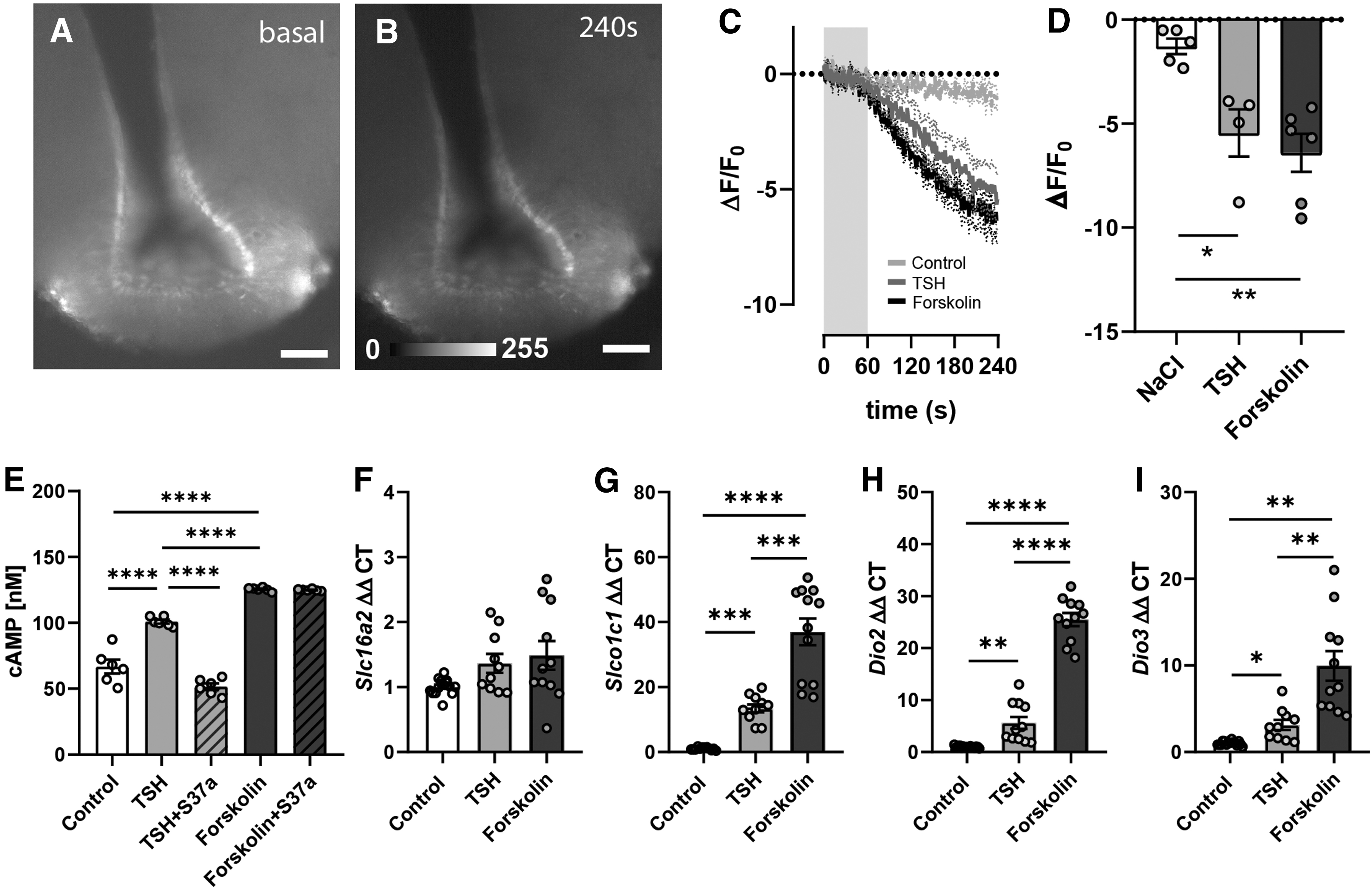
(
Therefore, we propose a cAMP-dependent regulation of Slc16a2, Slco1c1, Dio2, and Dio3. In line with this concept, activation of adenylyl cyclase by forskolin elevated the expression of Slco1c1 (Fig. 5G), Dio2 (Fig. 5H), and Dio3 mRNA (Fig. 5I) like TSH. Remarkably, forskolin had no effect on the expression of Slc16a2 mRNA (Fig. 5F) analogous to TSH.
To explore the signaling pathways linking cAMP to the regulation of the gatekeeper genes, we investigated activation of the CREB through phosphorylation. Phospho-CREB accumulated in nuclei of primary tanycytes after TSH and forskolin stimulation (Fig. 6A, B). The small molecule 666-15 that is known to inhibit the recruitment of coactivators by phosphorylated CREB reduced the transcriptional activity 30 (Supplementary Fig. S3). 666-15 did not affect Slc16a2 expression (Fig. 6C). However, the CREB inhibitor abolished the TSH-induced elevation of Slco1c1 (Fig. 6D), Dio2 (Fig. 6E), and Dio3 (Fig. 6F) mRNA, proving a CREB-mediated regulation of Slco1c1, Dio2, and Dio3.

(
In addition to Gαs, TSHR is Gαq-coupled and activates mitogen-activated protein kinases (MAPKs) through PKC. 24,31 Activation of Gαq by TSH was investigated by loading tanycytes, in acute brain slices, with Fura2. ATP, a known stimulator of [Ca2+]i in tanycytes, was used as a positive control. 19 The calculated ratio ΔF/F0 showed a rapid increase upon administration of TSH in the tanycytic layer (Fig. 7B), similar to ATP (Fig. 7C), demonstrating that TSH elevated [Ca2+]i in tanycytes.
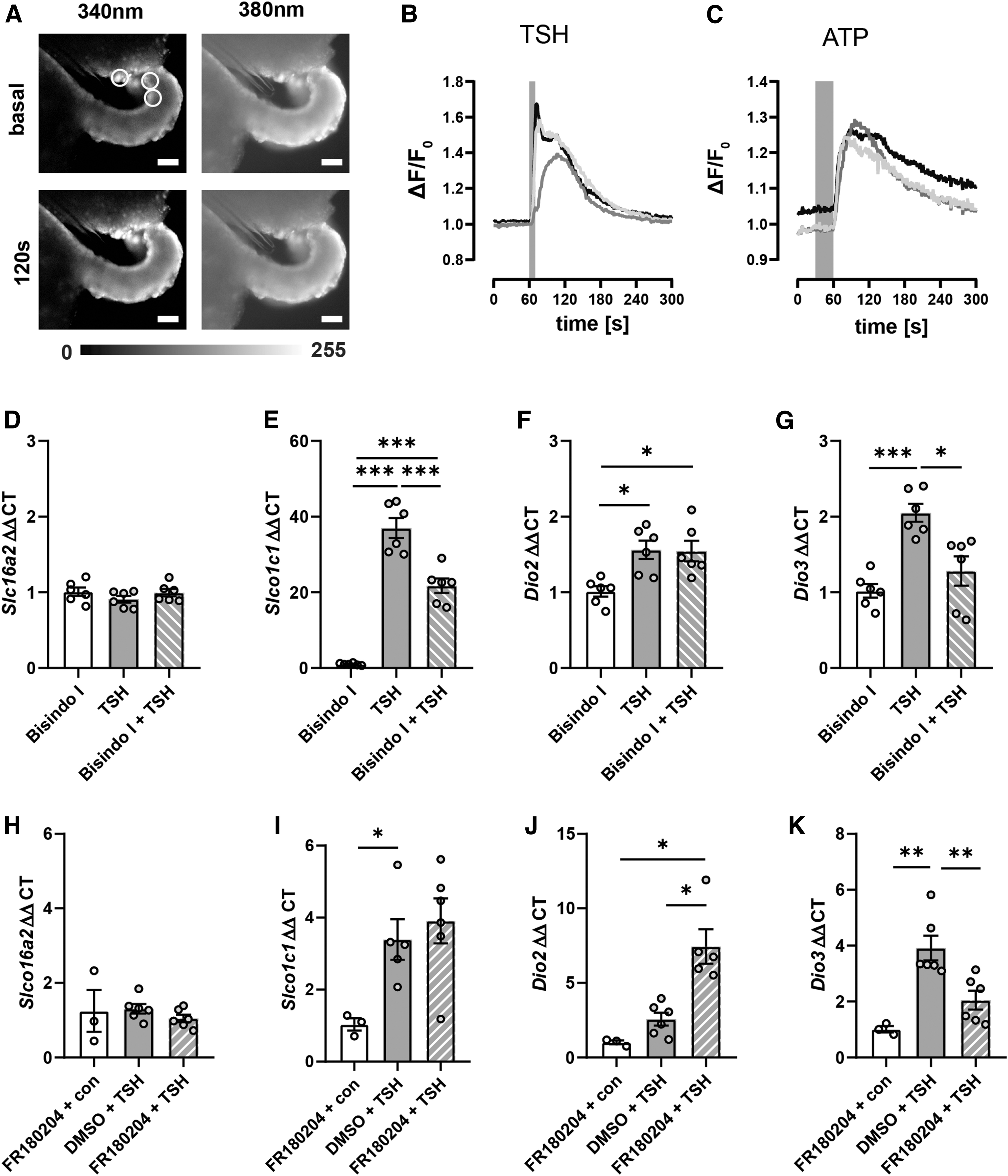
(
To identify the pathways linking [Ca2+]i increase to gene regulation, we used bisindolylmaleimide-I, an inhibitor of the Ca2+-dependent PKC (Fig. 7D–G). Bisindolylmaleimide-I significantly reduced Slco1c1 and Dio3 mRNA (Fig. 7E, G) but had no effect on TSH-stimulated Dio2 expression (Fig. 7F). Neither TSH nor bisindolylmaleimide-I influenced Slc16a2 expression (Fig. 7D). These results suggest a PKC-dependent regulation of Slco1c1 and Dio3 in addition to the previously described CREB-mediated regulation.
A target of PKC is the ERK that has been shown to be activated by TSH on primary tanycytes. 24 To test whether ERK mediates the regulation of TH-related genes in tanycytes, we used the ERK-inhibitor FR180204 32 (Fig. 7H–K). Congruent with other experiments, TSH did not influence Slc16a2 expression (Fig. 7H). Contrary to the findings with the PKC inhibitor (Fig. 7E), FR180204 did not inhibit the elevation of Slco1c1 mRNA by TSH (Fig. 7I). Interestingly, the ERK inhibitor further increased Dio2 expression (Fig. 7J) on TSH stimulation. In contrast, the increase of Dio3 mRNA was abolished by FR180204 (Fig. 7K). These results suggest that ERK mediates Dio3 expression on TSH stimulation but attenuates Dio2 induction.
Discussion
Through in situ hybridization performed on mouse brain sections and PCR analyses conducted on rat primary tanycytes, we verified the expression of key TH transporters (Slc16a2 and Slco1c1), as well as the deiodinases, Dio2 and Dio3 in tanycytes. These findings are consistent with previous studies conducted in various species such as mice, 10,11,33,34 rats, 35 hamsters, 36 and birds. 37 In line with previous findings, 38 –40 we demonstrated tanycytic expression of THRs and TSHR, which allow a response to the peripheral hormones of the HPT axis.
Among the THRs, Thra1 appears to be the dominant receptor, while only low mRNA levels of Thrb were present. 10,11 Characterizing the localization of the transcripts, we identified that Slc16a2 and Thra1 were present in all areas of the tanycytic layer. Dio2 and Tshr were mainly located and colocalized with tanycytes next to the arcuate nucleus and the ME. Dio3 and Slco1c1 transcripts were mainly found in tanycytes dorsal to the ME, but in low abundance.
In both normal and hypothyroid states, Dio3 mRNA expression in tanycytes is rather low, but increases in response to elevated T3 levels. 41,42 This appears to play a crucial role in protecting the hypothalamus against excessive TH stimulation. The distribution of the genes in the ependymal layer is in line with single cell sequencing results of Campbell et al. 10,11 Differences in the gene expression pattern of the gatekeeper genes support a region-specific function of tanycytic subpopulations.
Primary hypothyroidism is known to increase Dio2 expression in tanycytes. 43 So far, no TH response element has been found in the Dio2 promoter that could explain gene regulation by T3. 23,44 Instead, elevated TSH serum levels that occur in primary hypothyroidism may be responsible for increasing Dio2 mRNA in tanycytes. This notion is supported by experiments conducted on seasonal breeders (for review 45,46 ) and a study by Bolborea et al., 24 identifying TSH as a regulator of Dio2 transcription in tanycytes.
In addition, TSH elevated Slco1c1, which could be essential for a faster equilibration of altered TH concentrations, 47 and Dio3 mRNA levels. For all three genes, the induction depended on TSHR, as shown by the TSHR-antagonist S37a and TSHR siRNA knockdown. TSH did not regulate the expression of Slc16a2. Remarkably, the induction of Slco1c1, Dio2, and Dio3 by TSH, in acute brain slices, followed a distinct spatial pattern, very similar to the basal expressions. This subtype-specific regulation could be essential for the distribution and homoeostasis of TH in distinct hypothalamus nuclei to adapt TH-dependent functions such as HPT feedback, 48 energy metabolism, 5 and fertility 45 in hypothyroidism.
In addition, a TSH-dependent regulation of MAPK activity in tanycytes can influence the tanycytic leptin transport with an impact on feeding and energy homoeostasis. 14 These physiological changes can, in parts, be observed under global inhibition 49,50 or overactivation, by expressing constitutive active mutations, of the TSHR. 51 Tanycytic TSHR is perhaps involved in these processes, but this idea requires further validation.
As a GPCR, TSHR activates various G proteins. 52 TSH increases intracellular cAMP levels in tanycytes, leading to elevated expression of Slco1c1, Dio2, and Dio3 mRNA through TSHR. Stimulation with forskolin, an adenylate cyclase activator, mimics TSH-mediated gene expression pattern, highlighting the importance of cAMP. Furthermore, both TSH and forskolin stimulation resulted in the phosphorylation of the transcription factor CREB. Using the CREB inhibitor 666-15, we have shown that CREB plays a crucial role in mediating TSH effects on Slco1c1, Dio2, and Dio3 transcription. These results confirm a TSHR- and CREB-dependent mechanism for regulating Slco1c1, Dio2, and Dio3 mRNA in primary tanycytes. This regulation aligns with similar findings in other cell types involved in cAMP-dependent gene regulation. 23,44,53,54
Bolborea et al. have shown a TSH-dependent increase in phospho-ERK in primary tanycytes, which was independent of the Gαs subunit, 24 indicating another pathway. In the thyroid gland, TSHR is furthermore Gαq-coupled. 31,52,55 We demonstrate, in this study, the TSH-dependent increase of [Ca2+]i in tanycytes measured by Fura2. In Gαq-coupled pathways, [Ca2+]i and diacylglycerol activate PKC, which in turn leads to an activation of the MAPK pathway.
TSH-induced elevation of Dio3 mRNA was abolished by inhibition of PKC or ERK. This result corroborates studies that show an ERK-mediated increase of Dio3 expression in astrocytes. 56 In thyroid carcinoma cells, several inhibitors of the MAPK pathway reduce the expression of Dio3 mRNA modulated by ERK. 57 In our study, TSH stimulated Dio2 mRNA expression in tanycytes, which was further increased in the presence of the ERK inhibitor. This indicates that the activation of ERK counteracts the CREB-dependent regulation of Dio2 in tanycytes. ERK may mediate the inhibitory effect of PKC on the cAMP-induced Dio2 expression, as reported by Imai et al. 58
Conclusion
TSH from the pituitary has a significant effect on the regulation of TH-gatekeeper genes in tanycytes. This points to a TSH-dependent feedback, which influences local TH supply to the hypothalamus. Interestingly, the effect of TSH differs between specific subpopulations of tanycytes and is based on distinct intracellular signaling pathways. In peripheral hypothyroidism, TSH can maintain TH supply by increasing Dio2 and Slco1c1 to adapt to the low TH levels. Parallel induction of Dio3 in the dorsal ventricular wall could enhance negative feedback on the HPT axis, decrease sympathetic output to BAT, 5 and affect fertility 45 by further decreasing local TH concentrations. However, whether the TSH-mediated effects on tanycytes are involved in the regulation of hypothalamic functions needs to be investigated in vivo by tanycyte-specific manipulations of TSHR.
Footnotes
Acknowledgments
We thank Ines Stölting and Wiebke Brandt for their technical assistance. We also thank Tetsuya Kitaguchi for providing the Flamindo2 plasmid (deposed by Addgene; plasmid No. 73938).
Authors' Contributions
A.C. contributed to investigation (lead), resources (lead), visualization (support), formal analysis (equal), writing—original draft (equal); P.M.S. assisted with investigation (support), visualization (support), formal analysis (support); V.N., M.R., and F.S. contributed to investigation (support), resources (support); K.G. performed methodology (support), writing—review and editing (equal); V.P. assisted with funding acquisition (equal), methodology (support), writing—review and editing (equal); M.S. contributed to conceptualization (support), funding acquisition (equal), writing—original draft (support), writing—review and editing (equal); H.M.-F. supported with conceptualization (lead), funding acquisition (lead), visualization (lead), formal analysis (equal), writing—original draft (equal).
Author Disclosure Statement
All authors declare that they have no interests, financial resources, or employment relationships that improperly influence or affect the integrity of the submission.
Funding Information
H.M.-F. received funding from the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation; Project-ID 424957847-CRC/TRR296, P02); M.S. received funding from the Deutsche Forschungsgemeinschaft (Project-ID 424957847-CRC/TRR296, P01) and from the European Research Council (ERC) Synergy Grant-2019-WATCH-810331; V.P. received support from the ERC Synergy Grant-2019-WATCH-810331; all other authors received no additional support, which were used for this work.
Supplementary Material
Supplementary Data