
Abstract
Background:
As an actin cytoskeleton interactor, PDZ (postsynaptic density 65-discs large-zonula occludens 1) and LIM (abnormal cell lineage 11-isket 1-mechanosensory abnormal 3) domain protein 7 (PDLIM7) was supposed to play an essential role modulating cytoskeleton. Focal adhesions (FAs), which are located at the cytomembrane terminus of actin cytoskeleton, function as a force sensor and can transform the mechanical signal to a biochemical signal. Focal adhesion kinase (FAK) localizes to and regulates signal transduction in FAs, which play an essential role in cell polarity, migration, and invasion. However, whether PDLIM7 is involved in FAs-associated signal transduction and its role in tumor invasion and metastasis remains largely unknown.
Methods:
A cohort of 80 patients with papillary thyroid carcinoma (PTC) at The Second Affiliated Hospital of Guilin Medical University, as well as 512 PTC samples from The Cancer Genome Atlas thyroid cancer database was utilized to analyze the expression of PDLIM7 and its association with prognosis. Survival data were assessed using Kaplan-Meier curves, whereas clinicopathological characteristics such as age, sex, tumor size, multilocality, extrathyroidal extension, lymph metastases, Hashimoto's thyroiditis, distant metastasis, and TNM stage were considered. Functional assays were performed in vitro and in a xenograft mouse model to assess the role of PDLIM7 in PTC cell lines. The colocalization of PDLIM7 with FAK and integrin alpha V (ITGAV) was determined using immunofluorescence assay and immunoprecipitation assay. Protein expression levels in cell and tissue biopsies were measured through Western blotting and immunohistochemistry.
Results:
(1) The PDLIM7 protein frequently upregulated in both PTC tissues and cells, and overexpression of PDLIM7 is associated with advanced clinicopathological characteristics. (2) Knockdown of PDLIM7 effectively inhibits cell proliferation, migration, and invasion in PTC cell lines in vitro. (3) Knockdown of PDLIM7 hinders the growth and metastasis of TPC-1 xenografts in vivo. (4) PDLIM7 demonstrates colocalization with both FAK and the FA molecule ITGAV and the knockdown of PDLIM7 resulted in disassembly of FAs and proteosome-dependent degradation of FAK in PTC cell lines.
Conclusions:
PDLIM7 function as an oncoprotein in PTC to promote metastasis, and a novel underlying mechanism is proposed that PDLIM7 keeps FAK protein from proteosome-dependent degradation.
Introduction
Papillary thyroid carcinoma (PTC) is the most common thyroid cancer, with high incidence of lymphatic metastasis and distant metastasis, ∼30% to 59.1%. 1,2 Notably, lymphatic metastasis frequently manifests as an initial symptom on diagnosis, whereas distant metastasis serves as the primary contributor to mortality in individuals. 3,4 However, the published data mining the molecular basis of pathological processes of PTC metastasis, which could be translated into clinical diagnosis and therapy, is scanty.
PDZ (postsynaptic density 65-discs large-zonula occludens 1) and LIM (abnormal cell lineage 11-isket 1-mechanosensory abnormal 3) domain protein 7 (PDLIM7), a member of the PDZ-LIM protein family, contains one PDZ domain and three LIM domains. 5 PDLIM7 functions as a scaffold that, via its PDZ domain, localizes LIM-binding proteins to actin filaments of both skeletal muscle and non-muscle tissues to assemble complexes on actin filaments.
Utilizing these complexes, PDLIM7 transmits tension signaling of actin cytoskeleton into cytoplasm or nuclear and then modulates a series of pathophysiological processes, including development of skeletal muscle, 6,7 platelet-mediated hemostasis, 8 and the integrin-mediated mechanical conduction. 9 PDLIM7-contained actin filament-associated complex transmits Ret proto-oncogene/papillary thyroid carcinoma 2 (Ret/ptc2) mitotic signaling, contributing to cell proliferation. 5,10
Elbediwy et al. reported that PDLIM7 were recruited to focal adhesions (FAs) and then triggered Yes-associated protein signaling cascade in the process of cell proliferation and organ growth. 9 However, the role of PDLIM7 in modulating FAs and focal adhesion kinase (FAK), which are strictly assembled on the ends of actin filaments and play an essential role in the processes of cell migration and invasion, is largely unknown yet.
FAs are contacting points for the cell body with the extracellular matrix (ECM), and function as force sensors that transform the mechanical signal from the extracellular microenvironment to the biochemical signal, which is involved in modulating cell proliferation, migration, and spreading. 11 During cell migration, cell body gained mechanical force through continuous assembly of FAs, then tipping the dynamic balance between assembly and disassembly of FAs, for cell moving. 12,13
FAK bridges actin cytoskeleton with FA complexes as an adaptor independent of its kinase activity, 14 localizes to FAs for transducing FAs-associated signaling, 15,16 and plays an essential role in cell mobility. 17 As a key executor of FAs signaling events, the activation and stabilization of FAK was finely tuned for driving cell migration. 18,19
Of note, both PDLIM7 and FAK interact with the actin cytoskeleton, and the mechanical forces could promote the maturation of FAs and enrichment of PDLIM7 on the stress fibers. 20,21 However, whether PDLIM7 could play a role in the formation of FAs and modulating the activity of FAK still needs to be explored.
In this study, we found that PDLIM7 colocalized with FAK on FAs and protected FAK protein from ubiquitin-proteosome-dependent degradation. PDLIM7 functions as an oncoprotein to promote invasion and metastasis of PTC cell lines. These findings proposed a novel mechanism used by PDLIM7 to regulate FAs-associated signaling events in PTC progression.
Materials and Methods
Tissue samples and cell culture
A total of 80 paraffin-embedded biopsies and 15 frozen samples, along with their corresponding non-cancerous tissues (NT), were acquired from the Department of Pathology at The Second Affiliated Hospital of Guilin Medical University in Guilin, China, between the years 2015 and 2018. These samples were pathologically confirmed as primary thyroid papillary carcinoma and were obtained from patients who had not undergone any radiotherapy or chemotherapy before surgery.
The demographic analysis of the included population is summarized in Supplementary Table S2. The use of human materials in this study adhered to the principles outlined in the Declaration of Helsinki and was approved by the Biomedical Ethics Committee of the Second Affiliated Hospital of Guilin Medical College (EFY-GZR2022007), and informed consent was obtained from patients for the use of their tissues.
The Human PTC cell lines (KTC-1, TPC-1, K1, B-CPAP), human poorly differentiated thyroid carcinoma cell line (WRO), and human immortalized thyroid follicular epithelial cells (Thy-ori-3-1) were obtained from the Cell Bank/Stem Cell Bank (Shanghai, China). These cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) under a humidified atmosphere containing 5% CO2.
Monthly routine testing for Mycoplasma contamination was conducted using the Mycoalert Mycoplasma detection kit (Invitrogen; Thermo Fisher Scientific, Inc.). In addition, the authentication of all cell lines was performed using the GenePrint10 kit (Promega, France) to generate a short tandem repeat profile.
Hematoxylin-eosin staining and immunohistochemistry
In the hematoxylin-eosin (HE) staining procedure, tissue sections were subjected to dewaxing and dehydration. Subsequently, the sections were stained with hematoxylin for a duration of three minutes, followed by eosin staining for one minute. The sections were then fractionated using ethanol dehydration and sealed using neutral gum. Immunohistochemical staining was carried out according to previously established protocols, 22 and details regarding the antibodies used can be found in Supplementary Table S1.
In this study, positivity was operationally defined as the presence of any level of cytoplasmic staining. Conversely, a negative result was defined as the complete absence of staining. High expression was characterized by a robust staining observed in at least one-third of the cells exhibiting cytoplasmic staining. On the other hand, robust staining that was observed in less than one-third of the cells or more than one-third cells but appeared as faint staining was categorized as low expression.
Quantitative real-time polymerase chain reaction
The isolation of RNA and qRT-PCR assay has been fully described in previous publications. 22 GAPDH was utilized as the endogenous control. The primer sequences for PDLIM7 were as follows: The forward primer sequence was 5′ AGCATCGATGGCGAGAATG 3′, and the reverse primer sequence was 5′ TTCTGCGGTTTGCTCTGAA 3′. As for GAPDH, the forward primer sequence was 5′ ACCCAGAAGACTGTGGATGG 3′, and the reverse primer sequence was 5′TCTAGACGGCAGGTCAGGTC 3′.
Construction of stable knockdown/overexpression cell lines
KTC-1 and TPC-1 cells were subjected to infection with PDLIM7 shRNA Lentiviral Particles (Santa Cruz Biotechnology) to establish PDLIM7 stable knockdown cells. Similarly, K1 and B-CPAP cells were infected with PDLIM7 overexpression lentiviral vector (GENECHEM, China) to generate PDLIM7 stable overexpression cells.
Following the manufacturer's instructions, a total of 4 × 104 cells per well were seeded in a 24-well plate and incubated with lentivirus for transfection overnight. Infected cells were subsequently treated with 5 μg/mL puromycin (Solarbio Life Science, Beijing, China) for a duration of seven days, and the transfection efficiency was assessed using Western blotting and qRT-PCR.
Cell counting kit-8 assay and colony formation assay
The proliferation capacity of PTC cell lines was assessed using the cell counting kit-8 (CCK8) assay and colony formation assay. Specifically, 2 × 103 cells were seeded in each well of 96-well plates for the CCK8 assay, as previously described. 22,23 For the colony formation assay, the designated cells were plated in 6-well plates at a density of 2 × 102 cells per well and incubated for 14 days. Following incubation, the colonies were fixed with 4% paraformaldehyde for 30 minutes and stained with a 2.5% crystal violet solution for 15 minutes.
Scratch-wound healing assay and transwell assays
The migratory capacity of the specified cells was assessed using scratch-wound healing assays and transwell assays, as previously described in detail. 22,23 The distance migrated was measured at both 24 and 48 hours. In the transwell assay, 2 × 104 cells per well in 200 μL of serum-free 1640 medium were seeded onto the filter to assess migration, or onto the filter coated with Matrigel (Corning) to assess invasion. After 24 hours, the cells were stained with methylene blue and quantified using a microscope (IX71; Olympus, Japan).
In vivo tumorigenesis assay and metastasis assay of nude mice
The experiments conducted in this study were approved by the Animal Research Committee of Guilin Medical College (GLMC202105130). All animal experiments adhered to the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. The nude mice used in this study were housed in microisolator cages under sterile conditions.
Thymus-free BALB/C nude mice, aged six weeks, were obtained from SLAC ANIMALS (Shanghai, China) and were subcutaneously injected with TPC-1 cells (5 × 106) suspended in 200 μL of phosphate-buffered saline (PBS). After two weeks, the mice were euthanized, and the transplanted tumors were documented through photography and preserved in formalin.
To detect the metastatic ability of PTC cell lines, 5 × 105 cells in 100 μL of PBS were injected into the caudal vein of nude mice to establish an animal model of lung metastasis. Four weeks later, the lung metastases were counted and photographed.
Western blotting assays
Western blotting assays were conducted as previously described. 22 –24 The information of antibodies is summarized in Supplementary Table S1.
Immunofluorescence assay
Immunofluorescence assays were performed as previously described. 24 The information of antibodies is summarized in Supplementary Table S1.
Immunoprecipitation assay
The immunoprecipitation assay was conducted utilizing the BeaverBeads™ Protein A/G Immunoprecipitation Kit (Beaver Biotechnology, Suzhou, China) in accordance with the manufacturer's instructions. In brief, the antibody was subjected to a 15-minute incubation with protein A/G magnetic beads at room temperature. Subsequently, the cell lysates were incubated with the antibody-coated beads for one hour at room temperature. Following three washes with washing buffer, the beads were boiled in sodium dodecyl sulfate (SDS)-buffer (Solarbio Life Science) for five minutes at 95°C.
The immunoprecipitation assay of ubiquitin was conducted using the Signal-Seeker™ Ubiquitination Detection Kit (Cytoskeleton, Inc.) following the manufacturer's protocol. Briefly, plate cells were lysed in BlastR™ lysis buffer, and the protein contents were collected using a supplied filter. Subsequently, 5 mg of lysate was utilized for each immunoprecipitation assay with the Ubiquitination Affinity Bead, incubated for two hours at 4°C. The Bead Elusion Buffer was employed to incubate the Beads at room temperature for five minutes, and the immunoprecipitation sample was collected and boiled for five minutes.
Statistical analysis
The experiments were conducted in triplicate at least, and the statistical analysis was performed using SPSS 23.0 (SPSS, Inc.). Graphing was carried out using GraphPad Prism 9.0 (GraphPad, Inc.). A two-tailed Student's t-test was employed to compare protein levels, mobility, and other measures between the LV-(sh)SCR and LV-(sh)PDLIM7 groups.
χ 2 test was utilized to assess the variation in PDLIM7 expression across different tissues and clinicopathological parameters. The RNAseq data were obtained from the The Cancer Genome Atlas (TCGA) database (https://portal.gdc.cancer.gov) to extract both adjacent and cancer samples. A paired-samples T-test was employed to evaluate the disparity in expression levels between the adjacent and cancer samples. Analysis of overall survival (OS) was assessed using Kaplan-Meier analysis. Statistical significance was defined as p < 0.05 (*p < 0.05; **p < 0.01; # p < 0.001).
Results
PDLIM7 protein is frequently upregulated in PTC tissues and cells and overexpressed PDLIM7 is associated with advanced clinicopathological characters
Immunohistochemistry (IHC) assays showed that PDLIM7 were upregulated in PTC tissues (cases with positively expressed PDLIM7, 34/80, 42.5%), as compared with their paired adjacent NT, with no PDLIM7 being positively expressed in cases (0/80, 0%) (Fig. 1A, B and Table 1; p < 0.001). In a sample of 59 pairs of specimens obtained from TCGA, PDLIM7 mRNA exhibited upregulation in the majority of PTC tissues (54/59, 91.5%) when compared with their NT (Fig. 1C; p < 0.001).

PDLIM7 protein is frequently upregulated in PTC tissues, and overexpressed PDLIM7 is associated with advanced clinicopathological characters. (
Expression of PDLIM7 Proteins in 80 Cases of Papillary Thyroid Carcinoma and compared with Adjacent Non-Cancerous Tissues
NT, non-cancerous tissues; PDLIM7, PDZ (postsynaptic density 65-disks large-zonula occludens 1) and LIM (abnormal cell lineage 11-isket 1-mechanosensory abnormal 3) domain protein 7; PTC, papillary thyroid carcinoma.
In 15 fresh PTC specimens, PDLIM7 protein was upregulated in all cases, as compared with adjacent NT (Fig. 1D). Meanwhile, PDLIM7 mRNA and protein were upregulated in PTC cell lines when compared with immortalized thyroid epithelial cells, as shown by qRT-PCR and Western blotting respectively (Fig. 2A, B). Moreover, PDLIM7 protein expression was positively correlated with larger tumor size (T classification; p < 0.001), lymph node invasion (N classification, p = 0.002), extrathyroidal extension (YES vs. NO, p < 0.001), distant metastasis (M classification, p = 0.002), and high clinical stage (I–II vs. III–IV, p < 0.001) (Fig. 1E, F and Table 2).
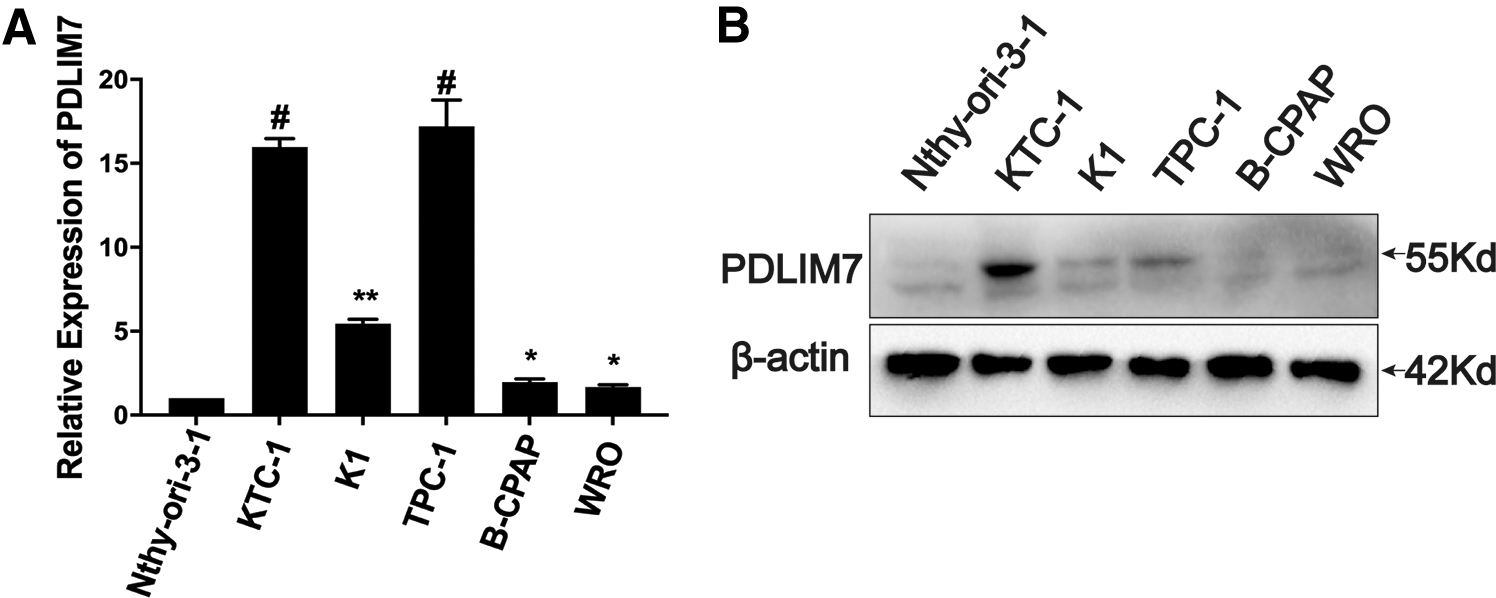
The expression level of PDLIM7 in the indicated PTC or PDTC cell lines and immortalized thyroid epithelial cell lines is assessed using qRT-PCR (
Expression of PDLIM7 in 80 Cases of Papillary Thyroid Carcinoma Patients with Different Clinicopathlogical Features
Simultaneously, based on the patients' information obtained from the TCGA database, a positive correlation was observed between PDLIM7 mRNA expression and various clinical factors, including pathologic T stage (T1 and T2 vs. T3 and T4, p = 0.006), N stage (N1 vs. N0, p = 0.032), extrathyroidal extension (YES vs. NO, p < 0.001), and high clinical stage (I–II vs. III–IV, p = 0.019) (Supplementary Table S3).
Further, an OS analysis was conducted using samples from TCGA. The samples were arranged based on the median RNA expression level, and subsequently, the highly expressed group of PDLIM7 (n = 101) was identified as the 80th to 100th quantile group, whereas the lowly expressed group (n = 101) was determined as the 0th to 20th quantile group. Kaplan-Meier analysis showed higher PDLIM7 is an unfavorable indicator of OS of PTC patients (Fig. 1G; p = 0.048). These results suggested a potential role of PDLIM7 in the development and progression of PTC.
PDLIM7 promotes cell proliferation, migration, and invasion of PTC cell lines
To investigate the role of PDLIM7 in the malignant behavior of PTC cell lines, PDLIM7 were successfully knocked down by infecting PTC cell lines with lentivirus haboring shRNA-expressing constructs. Western blotting and quantitative real-time polymerase chain reaction (qRT-PCR) showed that both mRNA and protein of PDLIM7 were drastically downregulated in KTC-1 and TPC-1 cells (Fig. 3A, B).

Knockdown of PDLIM7 inhibits cell proliferation, migration, and invasion of PTC cells. (
Silencing endogenous PDLIM7 inhibited cell proliferation, as indicated by results of CCK8 assays (Fig. 3C; shSCR vs. shPDLIM7, p = 0.17, p < 0.001, p < 0.001 and p < 0.001 in KTC-1, Fig. 3D; p = 0.015, p < 0.001, p < 0.001, and p < 0.001 in TPC-1 for 24, 48, 72, and 96 hours, respectively) and colony formation assay (Fig. 3E, F; shSCR vs. shPDLIM7, p < 0.001 and p = 0.007 for KTC-1 and TPC-1 respectively).
In addition, migrative ability of PTC cell lines was inhibited by knockdowned PDLIM7, as indicating by scratch-wound healing assays (Supplementary Fig. S1) and transwell migration assays (Fig. 3G, H; shSCR vs. shPDLIM7, p < 0.001 for KTC-1 and TPC-1 respectively). The Boyden assays showed that knockdown of PDLIM7 significantly hampered the invasive potency of PTC cell lines (Fig. 3I, J; shSCR vs. shPDLIM7, p < 0.001 for KTC-1 and TPC-1, respectively).
In addition, exogeneous PDLIM7 promoted cell proliferation (Supplementary Fig. S2C–F), migration (Supplementary Fig. S2G, H), and invasion (Supplementary Fig. S2I, J) of PTC cell lines K1 and B-CPAP. The earlier data showed that PDLIM7 plays an essential role in the malignant progression of PTC cell lines.
Knockdown of PDLIM7 inhibits growth and metastasis of TPC-1 xenografts in vivo
To further examine the growth-promoting effects of PDLIM7 on PTC cell lines in vivo, we performed subcutaneous tumor xenograft experiments in nude mice. shSCR- and shPDLIM7-expressing TPC-1 and KTC-1 cells were subcutaneously injected into nude mice. KTC-1 cells cannot form xenografts in nude mice even injected 107 per mouse.
The growth of TPC-1 xenografts, showing drastically decreased tumor size (Fig. 4A, B), retarded tumor growth (Fig. 4C), and decreased tumor weight (Fig. 4D; shSCR vs. shPDLIM7, p < 0.001), was inhibited by PDLIM7 silencing. In addition, silencing endogenous PDLIM7 inhibited lung metastasis of TPC-1 cells (Fig. 4E, F; shSCR vs. shPDLIM7, p < 0.001), which were injected into the tail vein of nude mice. HE staining confirmed the tumorlets of lung metastases (Fig. 4G). These findings suggested a potential role of PDLIM7 in cell growth and metastasis of PTC cell lines in vivo.
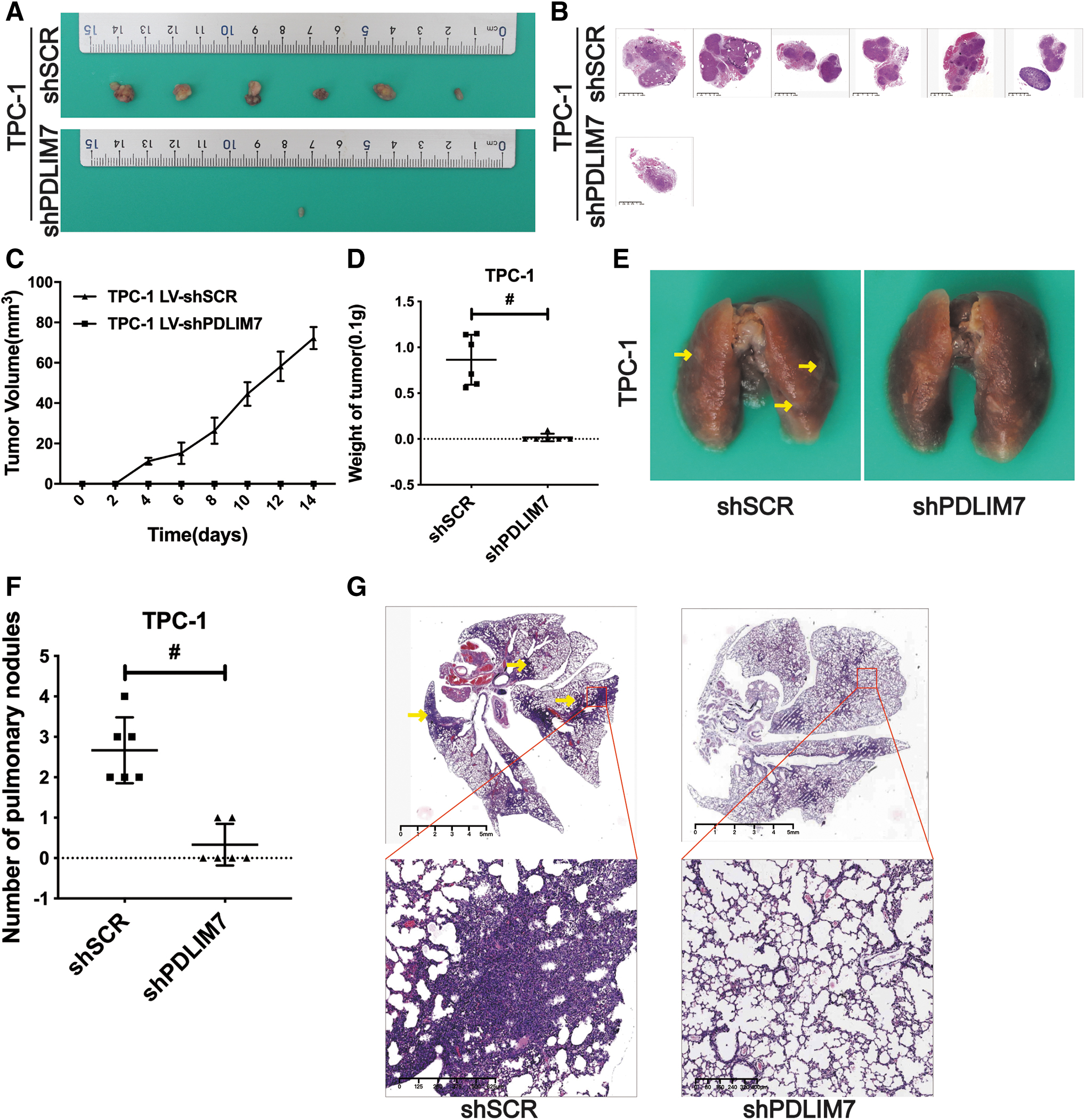
Knockdown of PDLIM7 inhibits growth and metastasis of TPC-1 xenografts in vivo. (
PDLIM7 colocalizes with FAK and the FA molecule integrin alpha V, and knockdown of PDLIM7 decreases the expression of FAK and integrin alpha V
FAs were reported to be linkers between ECM and the intracellular bundles of actin filaments named as stress fibers. Whether PDLIM7 localizing to actin filaments could modulate FAs and FAKs was explored in this study. Immunofluorescence assays showed FAK and integrin alpha V (ITGAV, a component of FAs) colocalized with PDLIM7 at the end of linear arrangement PDLIM7 particles (Fig. 5A).

PDLIM7 colocalizes with FAK and ITGAV and knockdown of PDLIM7 decreases the expression of FAK and ITGAV (
Meanwhile, the fluorescence intensity of FAK (shSCR vs. shPDLIM7, p < 0.001 in KTC-1 and TPC-1, respectively) and ITGAV-labeled FAs was all reduced after the knockdown of the PDLIM7 (shSCR vs. shPDLIM7, p < 0.001 in KTC-1 and TPC-1, respectively) (Fig. 5B–D). The Western blotting results showed that depletion of endogenous PDLIM7 by shRNA decreased the expression of FAK, p-FAK, and ITGAV in KTC-1 (shSCR vs. shPDLIM7, p = 0.038, p = 0.028, p = 0.017 and p = 0.021 for PDLIM7, ITGAV, FAK and p-FAK, respectively) and TPC-1 cells (shSCR vs. shPDLIM7, p = 0.014, p < 0.001, p < 0.001, and p < 0.001 for PDLIM7, ITGAV, FAK, and p-FAK, respectively) (Fig. 5E–G).
In mice xenografts, IHC assays showed that FAK and ITGAV were downregulated by knockdown of PDLIM7 (Fig. 5H). In the human PTC tissue, the expression of PDLIM7 was also positively correlated with the high expression of FAK (32/47, 68.1%, p < 0.001) and ITGAV (34/55, 61.8%, p < 0.001) (Fig. 5I, J and Supplementary Table S4). These data indicate that knockdown of PDLIM7 is involved in modulating FAs and FAK signaling.
PDLIM7 interacts with FAK and ITGAV, stabilizes FAK, and inhibits the ubiquitin-based degradation of FAK
PDLIM7 were reported to be involved in the assembly of an actin filament-associated complex essential for transmission stress-associated signaling. To investigate PDLIM7's role in assembly of FAs and FAK signaling, immunoprecipitation was used to evaluate the interactions between PDLIM7 proteins and FAs-associated protein FAK, ITGAV. FAK, ITGAV were pulled down by PDLIM7 antibody (Fig. 6A, a, b) and of course β-actin (Supplementary Fig. S3a, b).

PDLIM7 interacts with FAK and ITGAV, stabilizes FAK, and inhibits the ubiquitin-based degradation of FAK. (
Vice versa, FAK antibody can also pull down PDLIM7 proteins (Fig. 6A, c). In addition, ITGAV was also pulled down by FAK antibody (Fig. 6A, d). Consistent with the previous reported data, β-actin was also pulled down by PDLIM7 and FAK antibody. 14,25 The earlier data demonstrated that PDLIM7 colocalizes and interacts directly with FAK and ITGAV.
To investigate whether PDLIM7 could stabilize FAK proteins, MG-132 (5 mg/mL, two hours) was used to inhibit the proteosome-dependent degradation. MG-132 reversed the downregulation of FAK and then phosphate-FAK, which was induced by knockdown of PDLIM7 (Fig. 6B). Moreover, using ubiquitin antibody for immunoprecipitation increased ubiquitination of FAK, as observed in shPDLIM7 group (Fig. 6C). These results suggest that PDLIM7 stabilized FAK through inhibiting the ubiquitinated degradation of FAK, and then affected the quantity of phosphate-FAK.
Discussion
PTC has a good prognosis and long-term survival, but for patients with distant metastasis and recurrence, the prognosis worsens drastically. Distant metastasis is the main cause of death in patients with PTC. 26,27 The inconspicuous slow growth and subtle changed immunogenicity makes PTC frequently with regional lymph node metastasis at first diagnosis. 2 Therefore, it will be of great significance to explore the mechanism of PTC metastasis and develop potential intervention targets for management of thyroid cancer metastasis.
Here, we reveal that PDLIM7 is upregulated in PTC tissues (Fig. 1A–D) and cells (Fig. 2) and that PDLIM7 expression is significantly associated with aggressive clinicopathological characteristics, such as larger tumor size, lymph node metastasis, distant metastasis, and advanced clinical stage in PTC patients (Fig. 1E, F and Table 2 and Supplementary Table S3).
Meanwhile, a cohort from TCGA database indicated overexpressed-PDLIM7 as a poor prognostic factor for OS of thyroid cancer patients (Fig. 1G). Collectively, the results regarding the high level of PDLIM7 in the advanced-stage cases imply that PDLIM7 functions as a tumor promotor in the progression of PTC.
Several reports have confirmed that PDLIM7 is upregulated in many cancers. In gene expression analysis of skin tumors, PDLIM7 expression was higher in metastatic tumors. 28 In acute myeloid leukemia cases, PDLIM7 expression in peripheral blood and bone marrow, and high PDLIM7 and DNA methyltransferase 3 alpha mutation coexist with mean lower survival. 29 Similarly, high PDLIM7 expression in breast cancer is also associated with low survival. 30 This research suggests that PDLIM7 functions as a tumor promotor.
Our study also demonstrated that PDLIM7 has a tumor-promoting function in PTC cell lines. In papillary carcinoma cells KTC-1 and TPC-1, PDLIM7 knockdown attenuated cell proliferation, and it inhibited the migration and invasion of PTC cells (Fig. 3). In addition, exogeneous PDLIM7 promoted cell proliferation, migration, and invasion in K1 and B-CPAP cells (Supplementary Fig. S2).
In the nude mice xenografts, knockdown of PDLIM7 inhibited tumor growth of TPC-1 cells (Fig. 4). Meanwhile, knockdown of PDLIM7 decreased lung metastasis of TPC-1 cells in the mouse tail vein injection model. These data suggest that the PDLIM7 promoted the malignant progression of PTC cell lines and PDLIM7 acted as a tumor-promotor in PTC cell lines.
PDLIM7 is reported to activate the PI3K/AKT signaling pathway 31 and transmit the Ret/ptc2 signaling 5 to promote tumor proliferation in thyroid carcinoma. In addition, the literature has reported that PDLIM7 may interact with Afadin and Claudin-2, 32 enhance serum response factor/insulin like growth factor 2 mRNA binding protein 1 expression, and promote cell invasion. 33 The earlier data suggest that PDLIM7 can promote the proliferation and migration and invasion of thyroid carcinoma, which are consistent with our data in PTC cell lines.
Based on the fact that PDLIM7 is a cytoskeletal regulatory protein 8,9,25 and affects the ability of migration and invasion of PTC cell lines both in vitro and in vivo, the associations between PDLIM7 and FAs/FAK, which play an essential role in cell migration and invasion, were explored here. The positive correlation between PDLIM7 and FAK proteins or FAs marker ITGAV was documented in mice xenografts (Fig. 5H) and human tissue specimens (Fig. 5I).
Since PDLIM7 is mainly localized in the cytoskeleton, 8,9,25 FAs are also closely associated with the cytoskeleton, 34,35 suggesting a possible direct interaction between PDLIM7 and FAs. In fact, immunofluorescence assays showed that FAK or ITGAV colocalized with PDLIM7 at the membrane end of linear arranged PDLIM7 particles (Fig. 5A) in PTC cell lines.
Meanwhile, immunoprecipitation assays show a reciprocal interaction between PDLIM7 and FAK and that ITGAV could interact with both PDLIM7 and FAK (Fig. 6A). In addition, knockdown of PDLIM7, indeed, reduced the expression levels of FAK and ITGAV in PTC cell lines, as indicated by both immunofluorescence assays (Fig. 5B) and Western blotting assays (Fig. 5E).
Meanwhile, in accordance with the drastic downregulation of FAK, phosphorylated FAK was also decreased after PDLIM7 silencing in PTC cell lines (Fig. 5E). The data implied that the interactions among PDLIM7, FAK, and FAs could modulate FAK signaling and the assembly of FAs.
PDLIM7 was previously reported to interact with and prevent ubiquitinated degradation of P62/Sqstm136 and multiple endocrine neoplasia type 2A. 30 In this study, using proteosome inhibitor MG-132, decreased degradation of FAK (Fig. 6B) and accumulated ubiquitinated FAK (Fig. 6C) was observed in PDLIM7-silenced PTC cell lines. Since proteasomal degradation is the final pathway for inactivation of most proteins, 37 –39 the earlier data suggest an essential role of PDLIM7 in modulating FAK signaling and assembly of FAs through stabilizing FAK protein in a ubiquitin-proteosome dependent way (Fig. 7).
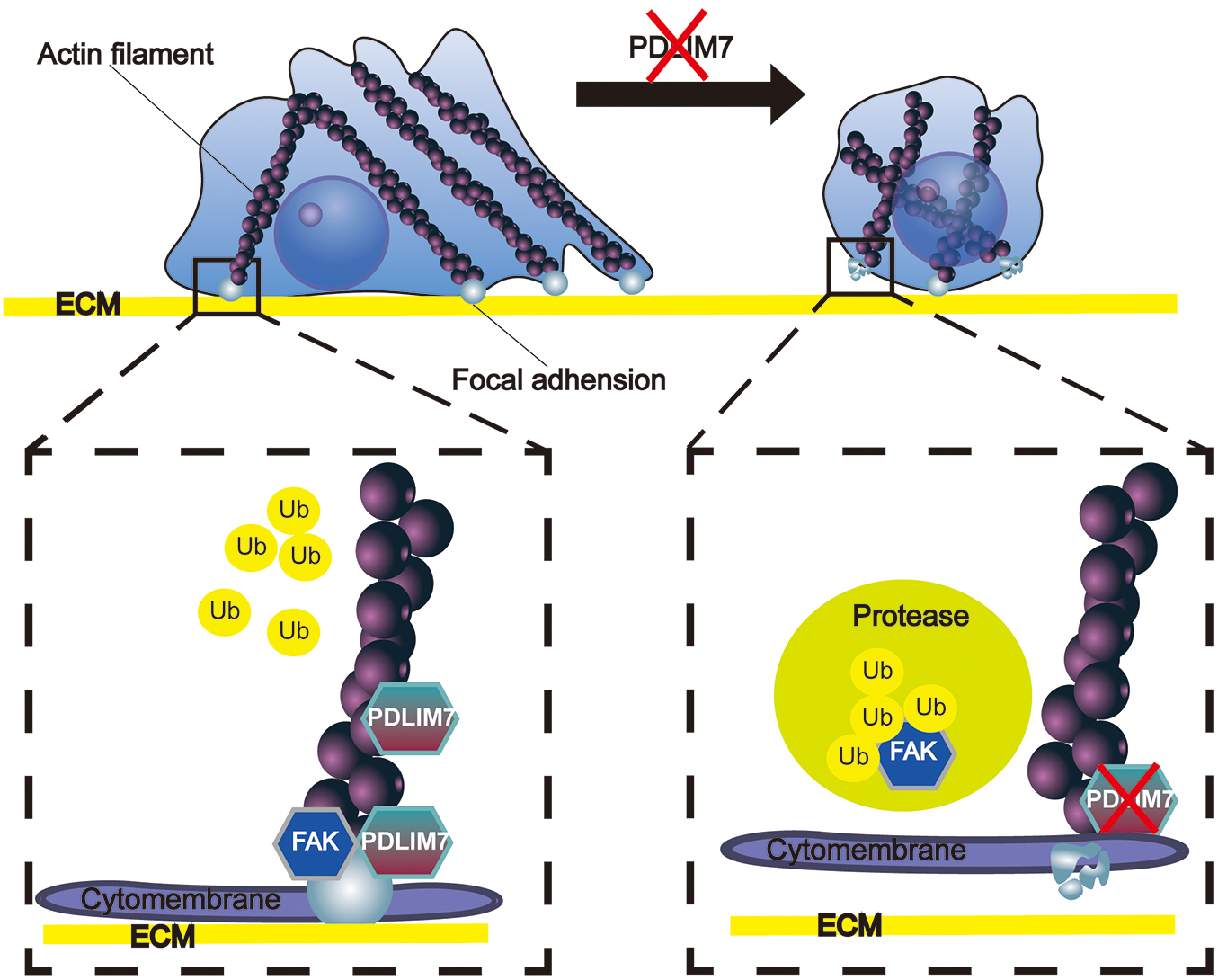
PDLIM7 stabilizes the FAK by inhabiting ubiquitin-based degradation of FAK. Under normal circumstances, PDLIM7 is localized on the Actin Filaments, specifically at the near-cell membrane region of the Actin Filaments terminal. PDLIM7 forms a strong binding interaction with FAK, resulting in its stabilization and promoting tight cell adhesion to the ECM. Conversely, when PDLIM7 is suppressed, FAK becomes destabilized and susceptible to cleavage by the ubiquitin-proteasome system. Consequently, FAs disassemble, leading to a loss of cellular adhesion to the ECM. Further, the diminished presence of PDLIM7 significantly impedes cell proliferation and metastatic capacity. ECM, extracellular matrix; FAs, focal adhesions.
In conclusion, PDLIM7, upregulated in PTC tissues and cells, interacts with, and stabilizes FAK proteins, modulates FAK signaling and FAs in the progression of PTC. Although the detailed role in FAs and the relationship with other FAs' and FAK-modulating proteins of PDLIM7 still need to be identified, our results provided a manipulatable target for the management of PTC, especially highly invasive and metastatic cases.
Footnotes
Authors' Contributions
F.Y. contributed to formal analysis and original draft. M.Y., Y.L., C.Z., and Y.C. were in charge of data curation and investigation. J.W. provided methodology. X.Z. performed conceptualization. S.X. was involved in project administration, funding acquisition, review, and editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China [82260476, 82060500].
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3