
Abstract
Background:
Monocarboxylate transporter 8 (MCT8) is the most specific thyroid hormone transporter identified to date, deficiency of which has been associated with severe intellectual and motor disability and abnormal serum thyroid function tests. However, it is presently unknown if MCT8, similar to other thyroid hormone transporters, also accepts additional substrates, and if disruption of their transport may contribute to the observed phenotype.
Methods:
In this study, we aimed to identify such substrates by applying liquid chromatography–mass spectrometry-based metabolome analysis in lysates of control and MCT8-overexpressing Xenopus oocytes. A subset of identified candidate substrates were validated by direct transport studies in transiently transfected COS-1 cells and human fibroblasts, which endogenously express MCT8. Moreover, transport characteristics were determined, including transport saturation and cis-inhibition potency of thyroid hormone transport.
Results:
Metabolome analysis identified 21 m/z ratios, corresponding to 87 candidate metabolites, with a 2.0-times differential abundance in MCT8-injected oocytes compared with controls. These metabolites included 3,5-diiodotyrosine (DIT) and several amino acids, including glutamate and glutamine. In accordance, MCT8-expressing COS-1 cells had 2.2-times lower intracellular accumulation of [125I]-DIT compared with control cells. This effect was largely blocked in the presence of 3,3’,5-triiodothyronine (T3) (IC50: 2.5 ± 1.5 µM) or thyroxine (T4) (IC50: 5.8 ± 1.3 µM). Conversely, increasing concentrations of DIT enhanced the accumulation of T3 and T4. The MCT8-specific inhibitor silychristin increased the intracellular accumulation of DIT in human fibroblasts. COS-1 cells expressing MCT8 also exhibited a 50% reduction in intracellular accumulation of [125I]-3-monoiodotyrosine (MIT). In contrast, COS-1 cells expressing MCT8 did not alter the intracellular accumulation of [3H]-glutamate or [3H]-glutamine. However, studies in human fibroblasts showed a 1.5-1.9 times higher glutamate uptake in control fibroblasts compared with fibroblasts derived from patients with MCT8 deficiency, which was not affected in the presence of silychristin.
Conclusions:
Taken together, our results suggest that the iodotyrosines DIT and MIT can be exported by MCT8. MIT and DIT interfere with MCT8-mediated transport of thyroid hormone in vitro and vice versa. Future studies should elucidate if MCT8, being highly expressed in thyroidal follicular cells, also transports iodotyrosines in vivo.
Introduction
The intracellular availability of thyroid hormones is governed by transmembrane transporter proteins, of which monocarboxylate transporter (MCT)8 is the most specific one identified to date. MCT8 facilitates the cellular uptake and efflux of thyroxine (T4), 3,3ʹ,5-triiodothyronine (T3), and to a lesser extent 3,3 ʹ,5ʹ-triiodothyronine (rT3) and 3,3ʹ-diiodothyronine (3,3ʹ-T2). 1 –3 Patients with mutations in MCT8 exhibit severe intellectual and motor disability attributed to low cerebral thyroid hormone concentrations and deranged metabolic features attributed to peripheral thyrotoxicosis. 4 –7 Although it is generally accepted that the major clinical hallmarks of MCT8 deficiency originate from defective cellular transport of thyroid hormone, it is not clear if defective transport of other, yet unknown, substrates may contribute to these features.
Available studies on the substrate specificity of MCT8 have only evaluated the direct transport of a limited number of potential substrates in MCT8-overexpressing Xenopus oocytes or mammalian cell lines, or applied cis-inhibition approaches that provide no definitive proof for direct transport. 1 –3,8 These studies showed that MCT8 does not transport aromatic amino acids, whereas MCT8-mediated T3 transport was reduced to a variable extent by different other iodothyronines. As an alternative for these targeted approaches, metabolome studies allow screening of a larger number of substrates and successfully identified substrates for orphan transporters, such as creatine for MCT12. 9 Detailing the substrate specificity of MCT8 would provide essential information to further understand MCT8 deficiency and guide the development of therapeutic approaches.
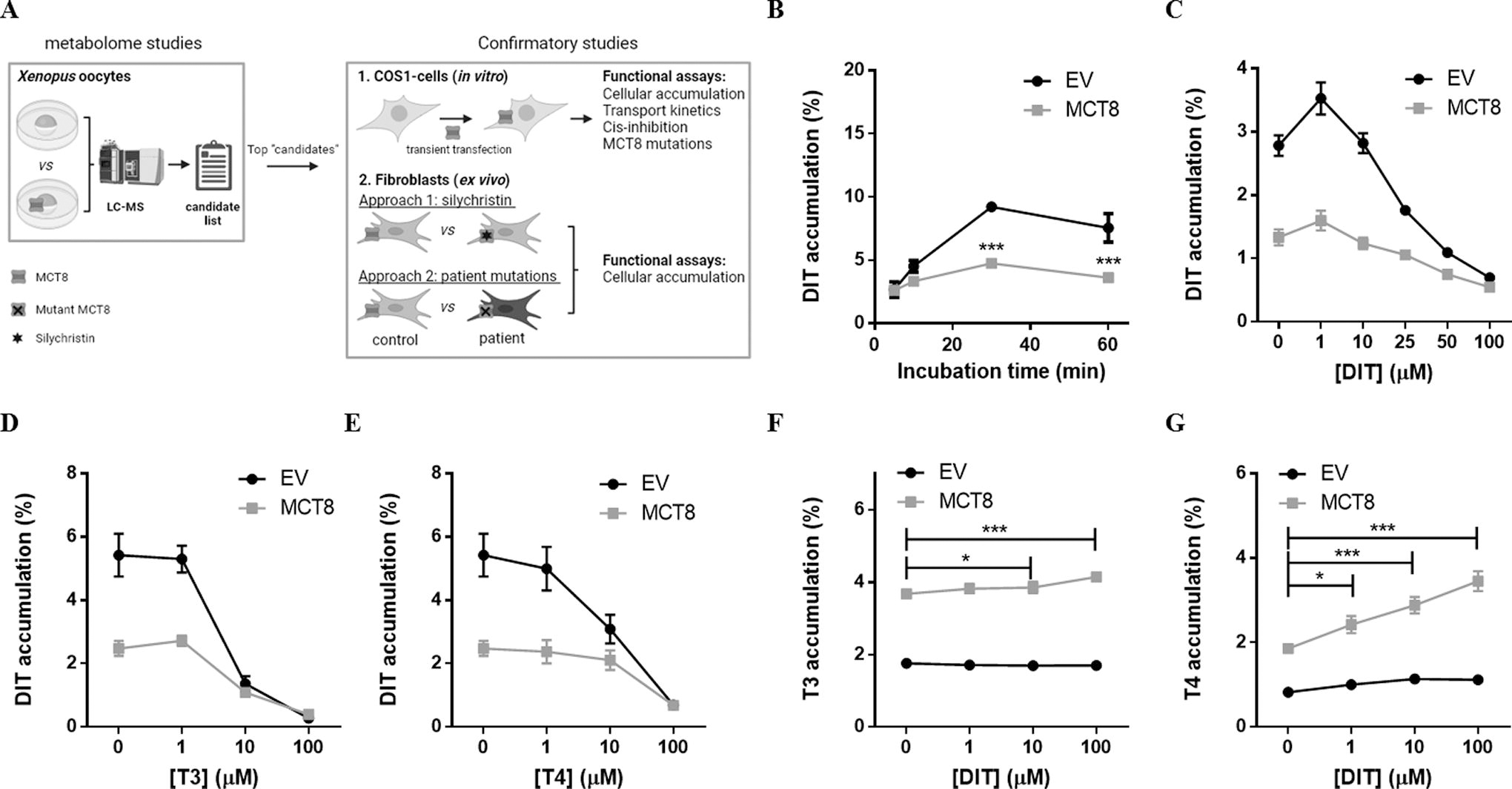
Therefore, we performed metabolome analyses in control and MCT8-overexpressing Xenopus laevis oocytes and validated potential novel substrates in transfected COS-1 cells and (patient-derived) human fibroblasts (Fig. 1A). With this approach, we identified diiodotyrosine (DIT) and [125I]-3-monoiodotyrosine (MIT) as new substrates for MCT8-mediated efflux.
Materials and Methods
Expression constructs, cell culture, and transfection
The cloning of MCT1, MCT8, flag-tagged LAT1, and LAT2 into pcDNA3 has been previously described. 1,8,10,11 Indicated MCT8 variants have been generated as described before. 12 –15 Mouse CD98-pcDNA3 was kindly provided by Dr. Krause (Leibniz-Forschungsinstitut für Molekulare Pharmakologie, Berlin, Germany). COS-1 African green monkey kidney cells (CVCL_0223) were obtained from ECACC (Sigma–Aldrich, Zwijndrecht, The Netherlands) and cultured and transfected under standard conditions. Fibroblasts of patients with MCT8 deficiency and healthy controls were derived from skin biopsies performed in the context of clinical care.
Metabolomics in Xenopus laevis oocytes
Oocytes were surgically removed from Xenopus laevis and handled as previously described. 16 After injection of 20 ng MCT8 or mock cRNA, oocytes were incubated for 2 days in ND96 medium (Hartmann Analytic, Brunswick, Germany) and subsequently for 1 hour or 48 hours in rich medium (n = 20) (see Abplanalp et al. 9 for composition), which was supplemented with metabolites mimicking human plasma 17 and Kao and Michayluk vitamin mix (Sigma Aldrich). 18 Both incubation media did not contain iodothyronines. Lysates were prepared and analyzed by liquid chromatography–mass spectrometry (LC-MS), as described before 9 and detailed in the Supplementary Data S1. The resulting MS data were processed by Marker Lynx XS 4.1 (Waters, Manchester, UK) and analyzed as previously described. 9,19
Functional studies in COS-1 cells and human fibroblasts
Transport studies were performed two days after transfection (COS-1 cells), three days after injection (Xenopus oocytes), or at >95% confluence (human fibroblasts) according to well-established protocols (see Supplementary Data S1). 1,20 For inhibition studies, increasing concentrations of unlabeled DIT, MIT, T3, or T4 (1–100 µM), 1 mM of the indicated amino acids, or 1 mM of the selective L-type amino acid transporter (LAT) inhibitor 2-aminobicyclo-(2,2,1)-heptane-2-carboxylic acid (BCH) were added during incubation. In fibroblast studies, intracellular accumulation of the involved substrates was adjusted for total protein levels measured by Bradford assay (Bio-Rad, Veenendaal, NL).
Immunoblotting and RNA expression studies were essentially performed as previously described (e.g., Zevenbergen et al. 10 and Groeneweg et al. 21 ). Full technical details are available in the Supplementary Data and Supplementary Table S1.
Definition of terms describing transport
In the Results section, we use the term intracellular accumulation to indicate the amount of substrate accumulated intracellularly after an incubation period. Intracellular accumulation is the net result of cellular uptake and cellular efflux. In the Discussion section of the article, we use the latter terms as potential explanations for our observations.
Statistical analyses
Statistical analyses were carried out using GraphPad Prism software (Version 5, GraphPad Software Inc., San Diego, USA). Apparent IC50 values were calculated based on the log(inhibitor) versus normalized response function in GraphPad Prism. All results are presented as mean ± SEM of at least 3 independent experiments performed in duplicate. Where appropriate, one-way or two-way ANOVA was applied followed by the Bonferroni posttest.
Ethical considerations
Skin fibroblasts were provided by care-giving physicians and concerned stored samples that had been obtained for diagnostic purposes. Written informed consent was provided by parents or legal representatives of the involved patients and controls. This study was carried out in agreement with the Medical Research Involving Human Subjects Act and was evaluated and granted by the Medical Ethics Committee of the Erasmus Medical Center, Rotterdam, The Netherlands (MEC-2015-362).
Results
Identification of DIT as substrate for MCT8 using metabolomics
Injection of Xenopus oocytes with MCT8 cRNA increased MCT8 protein expression levels and resulted in a 12.2 ± 2.1-times induction of T3 accumulation (Supplementary Fig. S1). After incubation for 1 hour and 48 hours in a rich medium, metabolome analyses on extracts of oocytes injected with MCT8 or mock cRNA yielded 743 unique mass/charge (m/z) ratios. After exclusion of analytical and chemical noise, restriction to known metabolites in human, and application of a 2.0-times differential abundance threshold, we identified 4 m/z ratios (corresponding to fragments of 14 candidate metabolites) to be less, and 10 m/z ratios (i.e., 42 candidate metabolites) to be more abundant in MCT8-expressing oocytes than in control oocytes after 1 hour of incubation (Table 1). In addition, 5 m/z ratios (i.e., 11 candidate metabolites) were less, and 9 m/z ratios (i.e., 50 candidate metabolites) were more abundant in MCT8-expressing oocytes than in control oocytes after 48 hours of incubation, of which 7 m/z ratios (i.e., 30 candidate metabolites) were overlapping between both time points (Table 1, for a full list of metabolites see Supplementary Table S2). Among the strongest hits, we identified a fragment with an m/z ratio of 431.86, corresponding to the characteristics of 3,5-diiodotyrosine (DIT), a precursor molecule of iodothyronines, which showed a 7.1-times lower accumulation in MCT8-expressing oocytes than in controls.
Overview of Identified m/z Ratios and Their Corresponding Putative Metabolites
Overview of candidate metabolites for MCT8-mediated transport based on the m/z ratios identified by metabolomic analyses in Xenopus oocytes queried against a database (see Supplementary Data S1 for details and selection criteria), assuming that all metabolites only form [M-H]-1 ions. Exemplary metabolites that are considered drugs, or specific to plants, herbs, or spices are presented in italics. The full list of candidate metabolites is provided in Supplementary Table S2. The relative abundance and p-value of the time point that did not meet our selection criteria (a 2.0-times differential abundance in MCT8-injected oocytes compared with controls, or vice versa, and a p-value < 0.05) are shown in italics.
Transport of DIT by MCT8
Transient overexpression of MCT8 in COS-1 cells reduced intracellular DIT accumulation compared with empty vector-transfected control cells, with a maximum of 2.2 ± 0.2-times difference after 30 minutes of incubation (Fig. 1B). This effect was not observed upon overexpression of MCT1, a transporter from the same family that does not transport iodothyronines (Supplementary Fig. S2). Cotransfection with the intracellular thyroid hormone-binding protein CRYM did not potentiate intracellular DIT accumulation, precluding specific evaluation of potential DIT uptake by MCT8 (data not shown). The difference in intracellular DIT accumulation between empty vector and MCT8-transfected cells (further referred to as MCT8-mediated efflux) declined with increasing concentrations of DIT, with an apparent IC50 of 18.8 [CI 13.0–27.3] μM and near-complete inhibition in the presence of 100 μM of unlabeled DIT; although at these concentrations also the cellular accumulation in control cells was greatly reduced (Fig. 1C). MCT8-mediated DIT efflux was inhibited at lower concentrations of T3 (apparent IC50: 2.5 [CI 1.0–6.]) μM) or T4 (apparent IC50: 5.8 [CI 3.0–11.1] μM) (Fig. 1D and E, respectively). Increasing concentrations of DIT progressively enhanced MCT8-mediated intracellular T3 and, in particular, T4 accumulation in the absence of CRYM with strongest effects observed in the presence of 100 μM of DIT (Fig. 1F and G, respectively). Together, these observations suggest that MCT8 primarily facilitates the cellular efflux of DIT, which may compete with MCT8-mediated efflux of T3 and T4, and vice versa.
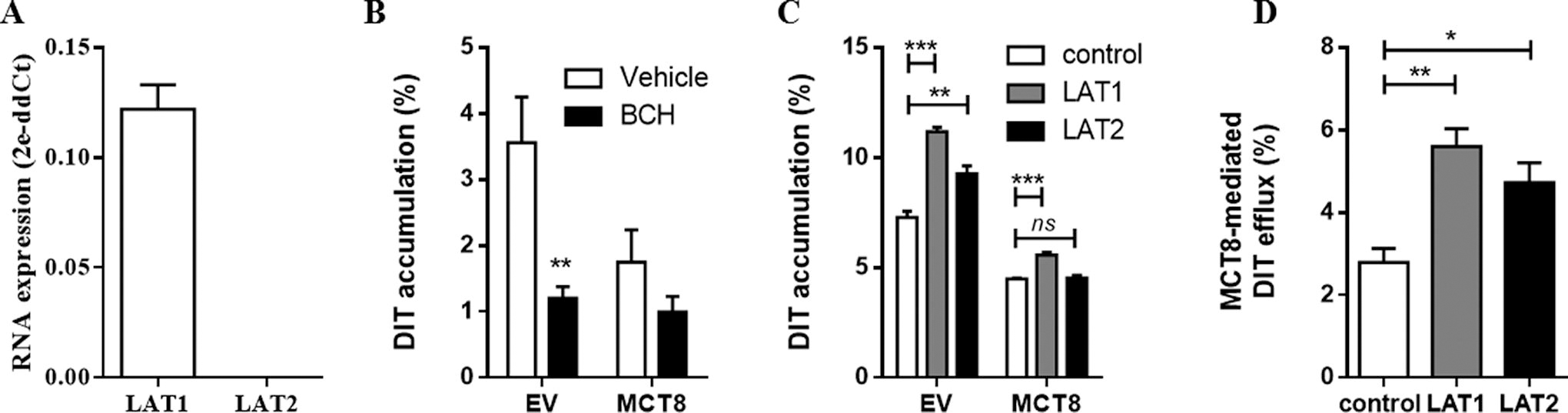
As MCT8 transports its substrates by facilitated diffusion, the concentration gradient of its substrates importantly determines transport efficacy and direction. To identify endogenous DIT transporters in COS-1 cells and thereby enable modulation of the DIT concentration gradient across the cell membrane, we performed quantitative PCR, which showed high mRNA levels of the
The transport of DIT is impaired in MCT8 mutants
To study if MCT8-mediated DIT transport is altered by mutations present in patients with MCT8 deficiency, we selected several well-characterized MCT8 mutations 12 –15,22,23 with variable impact on T3 transport capacity (Fig. 3A). Accordingly, these mutations reduced MCT8-mediated efflux of DIT (Fig. 3B). The impact of mutations on MCT8-mediated intracellular T3 accumulation and DIT efflux strongly correlated (R 2 = 0.88, p < 0.01), further supporting direct efflux of DIT by MCT8 (Fig. 3C).
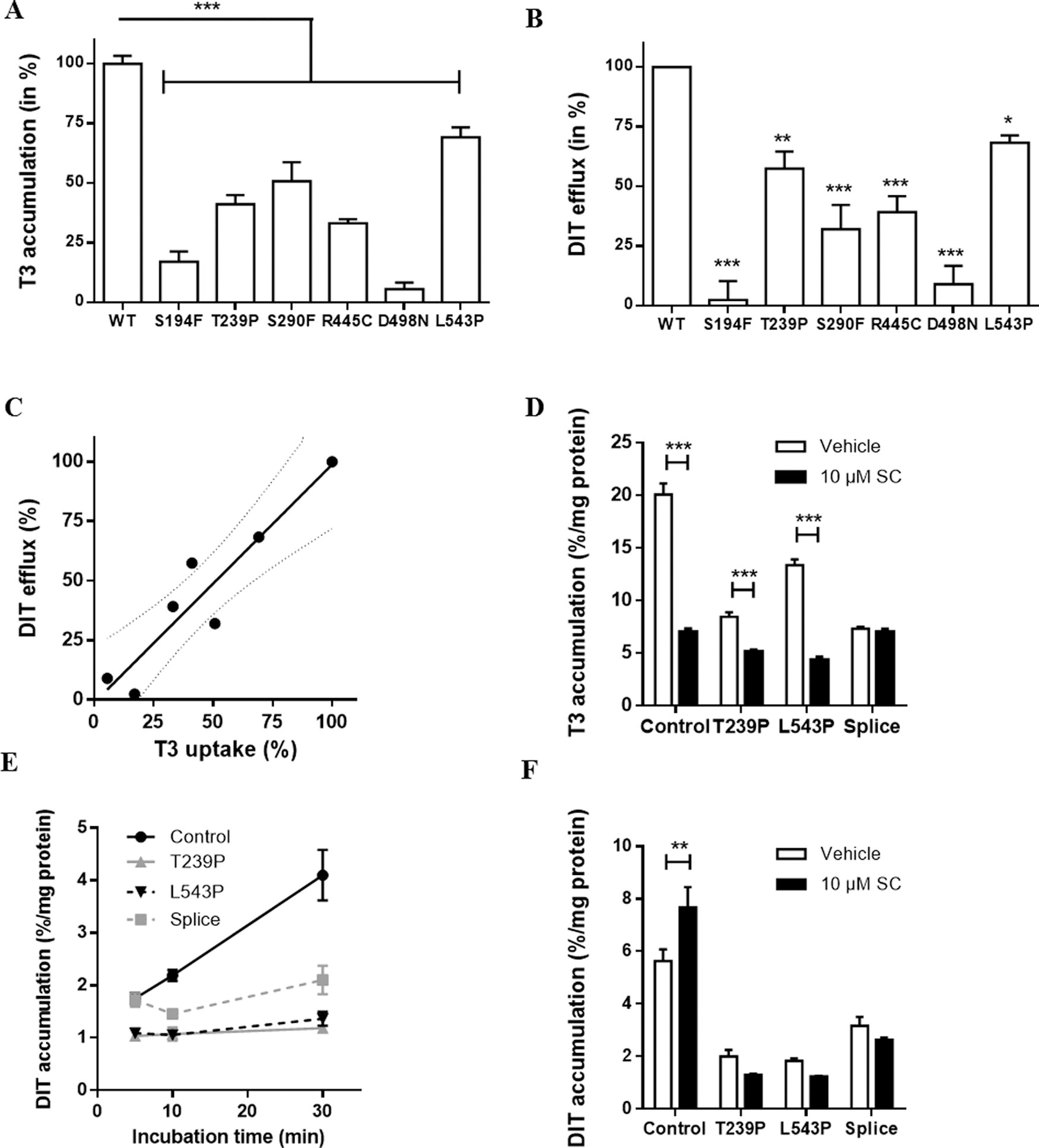
We then studied the effect of MCT8 inactivation on intracellular DIT accumulation in human fibroblasts, a well-established ex vivo model to study MCT8 function. Therefore, we assessed the effect of the MCT8-specific inhibitor silychristin (approach 1) and compared fibroblast cultures from healthy controls with those of three patients with MCT8 deficiency harboring a mutation with a relatively less severe (T239P and L543P) or severe (splice-site mutation resulting in a p.S267-S360del, further referred to as splice-site mutant) impact on MCT8 function (approach 2). 15,24 In line with previous studies (e.g., Masnada et al. 15 and Groeneweg et al. 20 ), cellular T3 accumulation was lower in the patient-derived fibroblasts compared with controls. The MCT8-specific inhibitor silychristin 25 reduced T3 accumulation in control fibroblasts by 64.9 ± 1.2%, whereas no reduction was observed in the splice-site mutant fibroblasts, confirming the complete loss of MCT8 function in these cells (Fig. 3D). Silychristin had intermediate effects in the T239P and L543P fibroblasts, and reduced T3 accumulation by 39.2 ± 4.8% and 67.5 ± 0.9%, respectively. Compared with the COS-1 cell line, the absolute intracellular accumulation of DIT was 10-times lower in human control fibroblasts (Supplementary Fig. S3). The accumulation of DIT in control fibroblasts exceeded the levels observed in any of the patient fibroblast lines (Fig. 3E). In the presence of silychristin, intracellular DIT accumulation significantly increased in control fibroblasts, whereas no significant changes were observed in patient-derived fibroblasts, suggesting that MCT8 also primarily facilitates the efflux of DIT in this system (Fig. 3F).
Transport of monoiodotyrosine by MCT8
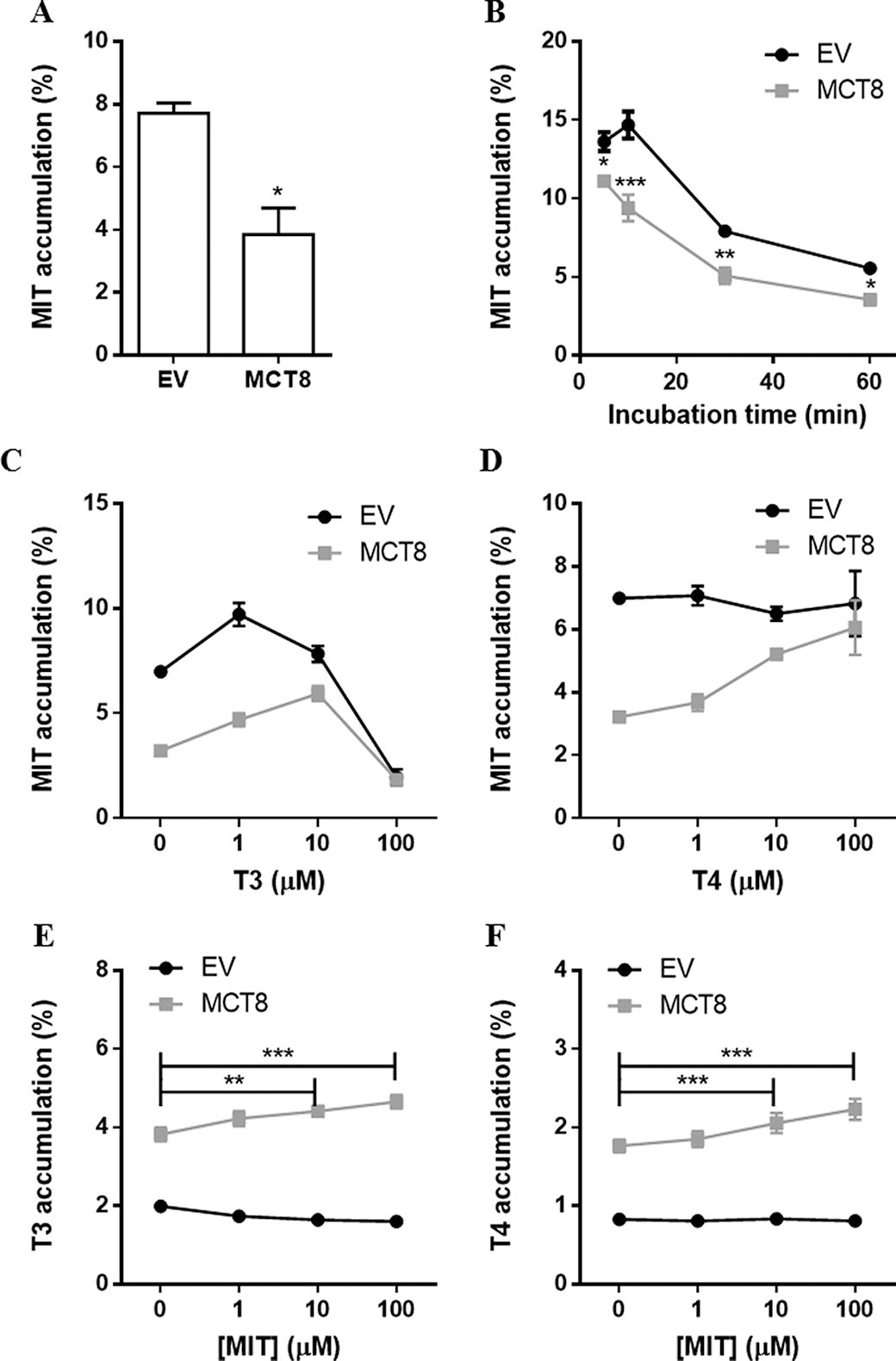
Given its close structural and chemical resemblances to DIT, we also studied if MCT8 facilitates the efflux of MIT. Indeed, the intracellular accumulation of MIT was 40.1 ± 5.5% lower in cells transfected with MCT8 than in empty vector-transfected control cells after 30 minutes of incubation (Fig. 4A), with the largest reduction in intracellular MIT accumulation after 10 minutes of incubation (Fig. 4B). MCT8-mediated efflux of MIT was largely diminished in the presence of 10 μM T3 and T4, with respective apparent IC50 values for T3 of 6.7 [CI, 3.0–15.0] μM and T4 of 6.4 [CI 3.0–14.0] μM (Fig. 4C and 4D). MCT8-mediated intracellular accumulation of T3 and T4 in the absence of CRYM was slightly, but statistically significantly, increased in the presence of 10 and 100 μM of MIT (Fig. 4E and 4F). BCH reduced the intracellular accumulation of MIT in empty vector-transfected control cells with 82.0 ± 3.4%, suggesting that LAT1 facilitates MIT uptake in COS-1 cells.
MCT8 does not directly transport amino acids
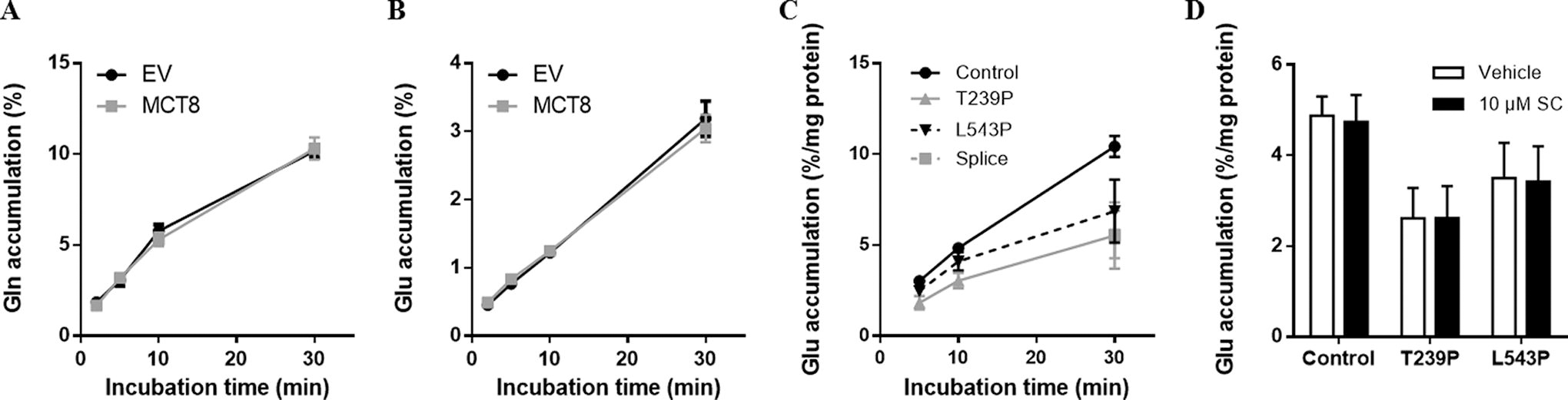
Some other m/z ratios identified in the metabolome analysis corresponded to fragments of amino acids or derivatives thereof, most notably threonine, glutamate, and glutamine as well as products of aspartic acid, proline, and arginine metabolism (Table 1), but, in line with previous studies, 1 neither lactate nor pyruvate, which are well-known substrates of MCT1, 2, and 4, was among them. As several other thyroid hormone transporters, including MCT10, LAT1, and LAT2, also transport amino acids, we studied the effects of different amino acids on MCT8-mediated T3 and T4 transport in transiently transfected COS-1 cells. At a concentration of 1 mM, which is well above their physiological concentrations in human serum, none of the tested amino acids significantly reduced MCT8-mediated intracellular accumulation of T3 (Fig. 5A), whereas a slight, but statistically significant, increase in intracellular T4 accumulation was observed in the presence of 1 mM aspartic acid (Fig. 5B). In line with the metabolome studies in Xenopus oocytes, MCT8 did not facilitate the transport of [3H]-tyrosine, phenylalanine, or tryptophan (substrates for MCT10), nor [5H]-alanine or leucine (substrates for LAT) (Supplementary Fig. S4A-E). We also studied the transport of glutamate and glutamine, selected based on the availability of their [3H]-radiolabeled isotopes and previous studies that showed alterations in these metabolites in brains of Mct8 knockout mice. 26 However, the intracellular accumulation of both amino acids did not differ between MCT8-transfected and control COS-1 cells (Fig. 6A and 6B). In line with the observations in Xenopus oocytes, glutamate accumulation in control fibroblasts significantly exceeded the levels observed in fibroblasts derived from MCT8 patients (Fig. 6C). Despite these differences, glutamate accumulation was not inhibited by silychristin (Fig. 6D), supporting that MCT8 may not directly transport glutamate, but rather modulates other cellular factors involved in the transport and/or metabolism of glutamate.

Intracellular accumulation of T3
Intracellular accumulation of glutamine
Discussion
We demonstrated that, in addition to iodothyronines, MCT8 also facilitates the cellular efflux of the iodotyrosines MIT and DIT. This process modulates MCT8-mediated transport of T3 and T4 in vitro and vice versa. Although metabolome analyses indicated that MCT8 also affects the intracellular concentrations of several amino acids or metabolites thereof, subsequent in vitro and ex vivo studies showed that MCT8 does not directly facilitate amino acid transport. Together, these findings illustrate that MCT8 not only regulates the intracellular concentrations of its direct substrates but also of metabolites that are not directly transported by MCT8 itself.
Although DIT was not included in the rich medium, it was among the strongest hits in the metabolome analyses. We hypothesize that DIT already accumulated inside the oocytes during routine culture and was subsequently released during the experiment. In line with the metabolome analyses and previous findings, 27 our validation studies in transfected COS-1 cells and human fibroblasts show that MCT8 reduces the intracellular accumulation of DIT. This indicates that MCT8 facilitates the cellular efflux of DIT; alternatively, MCT8 may reduce cellular uptake of DIT by endogenous DIT transporters via affecting their expression levels, transport efficacy, or transport direction. However, our studies with silychristin in human wild-type fibroblasts render the latter explanation less likely for the following reasons. First, given the short exposure time to silychristin and the absence of T3 in the incubation medium, alterations in expression levels of endogenous DIT transporters are unlikely. Second, these studies were performed in D-PBS, which is free of any potential substrates for MCT8. Therefore, we anticipate little effects on the availability of (co-)substrates for endogenous DIT-transporters and thus limited effects on their transport efficacy or direction. Although, as expected, insensitive to silychristin, DIT accumulation in patient-derived fibroblasts was lower than in those obtained from healthy controls. A potential explanation holds that the chronic hypothyroid condition in patient-derived fibroblasts alters the expression levels of potential, yet unknown, DIT transporters during routine cell culture.
Several approaches in which the concentration gradient across the cell membrane would favor cellular uptake of DIT, including studies in which potent endogenous transporters were blocked (COS-1 cells) or absent (human fibroblasts), did not support that MCT8 facilitates cellular uptake of DIT. Therefore, the substrate recognition criteria of MCT8 may differ between the intracellular and extracellular side of the transporter. Whereas the absence of the outer-phenolic-hydroxyl-group does not exclude DIT as a substrate for cellular efflux, its presence appears obligatory for cellular uptake. MCT8 also facilitates the efflux of MIT, which lacks the 5-iodine moiety compared with DIT, but not tyrosine, which lacks both iodine moieties, suggesting that the presence of at least one iodine-moiety on the (inner) phenolic ring is critical for substrate recognition during the efflux cycle.
The physiological relevance of MIT and DIT transport by MCT8 is unclear. MIT and DIT are intermediates formed in the thyroid gland during the biosynthesis of thyroid hormones. 28 As MIT and DIT are released from the thyroid into the bloodstream, 29 –31 given the high expression in the thyroid gland, MCT8 might be a good candidate to facilitate their efflux. 32,33
Increasing concentrations of DIT and MIT preferentially inhibited the efflux of T4 over T3. It is unclear if this can have consequences on circulating T3 and T4 concentrations under certain conditions. During TSH-receptor stimulation, circulating MIT and DIT concentrations increase. 34 Should the increased intrathyroidal MIT and DIT content under these conditions result in a relative inhibition of T4 over T3 export, this might represent an additional explanation for the elevated T3:T4 ratio in serum of patients with Graves’ disease (e.g., Izumi et al. 35 ).
MCT8 did not directly transport several other candidate substrates, which may have several reasons. First, other candidate substrates that yield the same fragmentation as those selected for the validation studies might be the actual substrates for MCT8. However, the lack of radiolabeled compounds or appropriate LC-MS detection systems precluded validation studies of all candidate substrates. Second, MCT8 may transport metabolites upstream or downstream in the pathway of the identified candidate metabolites, with the identified candidate substrates accumulating at the final or rate-limiting step. Third, depending on the MCT8-status, oocytes may have been exposed to different thyroid hormone concentrations during routine culture preceding the metabolome studies. For example, this may have affected glutamate/glutamine transport and metabolism, known to be responsive to thyroid state, with lower cellular glutamate uptake and utilization documented during hypothyroidism. 36,37 Indeed, we observed lower glutamate accumulation in fibroblasts derived from patients with MCT8 deficiency compared with healthy controls, providing an independent ex vivo model that confirmed our findings in Xenopus oocytes. As our in vitro and ex vivo studies focused on individual cell types, it is challenging to extrapolate these observations to the in vivo situation in which intermediate metabolism is regulated through a complex interplay of different organs. Future studies on the metabolome of patients with MCT8 deficiency may help elucidating the role of MCT8 in the regulation of intermediate metabolism in vivo.
Potential MCT8 substrates could have been missed with our approach. For instance, certain metabolites might have been absent in the incubation medium. Moreover, we used a 2-times differential abundance threshold for selection of candidate substrates, which may exclude potential substrates that are transported with low efficacy, or bidirectionally. In addition, synthetic and pharmacological substances were deliberately not added to the incubation medium and can therefore not be excluded as substrates for MCT8. Nevertheless, several drugs such as rocuronium and cefodizime, likely originating from procedures preceding the experiment, were detected and differentially present in oocytes expressing MCT8 versus controls. Finally, some substrates may require specific conditions to be transported by MCT8, which were not met in the present experiment.
Since none of the metabolites identified in the metabolome analyses exhibited a 10-times difference between control and MCT8-overexpressing oocytes as was observed for T3, our studies support the present concept that the primary function of MCT8 is the cellular transport of T3 and T4. Therefore, it is reasonable that therapeutic interventions focus on restoring thyroid hormone signaling. Indeed, compensation of defective thyroid hormone transport by T3 analogues that bypass MCT8, such as triiodothyroacetic acid (Triac), fully rescued the neurodevelopmental signs in various animal models for human MCT8 deficiency. 38 –40
Taken together, our studies identified DIT and MIT as novel substrates for MCT8 and thereby expand the substrate specificity of MCT8. Furthermore, deficiency of MCT8 may affect important pathways involved in intermediate metabolism, the implications of which need to be further studied. Nevertheless, in line with the present paradigm, our studies support that the iodothyronines T3 and T4 are the major substrates of MCT8.
Footnotes
Acknowledgments
The authors would like to dedicate this work to the memory of Theo Visser who passed away in 2018. He made a major contribution to the design and interpretation of the studies. They are also grateful for the technical assistance of Ramona EA van Heerebeek.
Authors’ Contributions
S.G., C.Z., E.C.L.d.S., F.S.v.G., E.L., S.M.R.C., and M.E.M performed the experiments. S.G., C.Z., B.K.-G., R.P.P., and W.E.V. designed the experiments and made the first draft of the article. All the authors critically evaluated the article.
Author Disclosure Statement
The authors have nothing to disclose.
Funding Information
This work was supported by an
Supplementary Material
Supplementary Data S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
Supplementary Table S2