
Abstract
Background:
Familial non-medullary thyroid carcinoma (FNMTC) is a genetically predisposed disease with unclear genetic mechanisms. This makes research on susceptibility genes important for the diagnosis and treatment options.
Methods:
This study included a five-member family affected by papillary thyroid carcinoma. The candidate genes were identified through whole-exome sequencing and Sanger sequencing in family members, other FNMTC patients, and sporadic non-medullary thyroid carcinoma patients. The pathogenicity of the mutation was predicted using in silico tools. Cell phenotype experiments in vitro and models of lung distant metastasis in vivo were conducted to confirm the characteristics of the mutation. Transcriptome sequencing and mechanistic validation were employed to compare the disparities between PAK4 wild-type (WT) and PAK4 mutant (MUT) cell lines.
Results:
This mutation alters the protein structure, potentially increasing instability by affecting hydrophobicity, intra-molecular hydrogen bonding, and phosphorylation sites. It specifically promotes phosphorylated PAK4 nuclear translocation and expression in thyroid tissue and cell lines. Compared with the WT cells line, PAK4 I417T demonstrates enhanced proliferation, invasiveness, accelerated cell division, and inhibition of cell apoptosis in vitro. In addition, it exhibits a significant propensity for metastasis in vivo. It activates tumor necrosis factor signaling through increased phosphorylation of PAK4, JNK, NFκB, and c-Jun, unlike the WT that activates it via the PAK4-NFκ-MMP9 axis. In addition, PAK4 MUT protein interacts with matrix metalloproteinase (MMP)3 and regulates MMP3 promoter activity, which is not observed in the WT.
Conclusions:
Our study identified PAK4: c.T1250C: p.I417T as a potential susceptibility gene for FNMTC. The study concludes that the mutant form of PAK4 exhibits oncogenic function, suggesting its potential as a novel diagnostic molecular marker for FNMTC.
Introduction
Familial non-medullary thyroid carcinoma (FNMTC) refers to families in which two or more first-degree relatives have thyroid carcinoma of follicular epithelial cell origin, unrelated to specific thyroid pathogenic factors. FNMTC represents ∼5% of all non-medullary thyroid carcinoma cases. 1,2 While non-medullary thyroid carcinoma was initially considered sporadic, recent studies have established a genetic predisposition for FNMTC, which results in a more aggressive behavior and worse prognosis compared with sporadic non-medullary thyroid carcinoma (SNMTC). 3 –6
Although a small proportion of FNMTC families is classified as syndromic FNMTC, most FNMTC cases are non-syndromic, and the underlying genetic mechanisms remain uncertain. 7
Mutations in genes such as SRGAP1 (12q14), 8 FOXE1 (9q22.33), 9 –15 DUOX2 (15q21.1), 16 CHEK2 (22q12.1), 17 –19 and NOP53 (19q13.33) 20 have been reported to potentially participate in the occurrence of FNMTC. However, in other known susceptibility genes and chromosomal loci, such as 14q32 locus (MNG1), 21 –25 PTCSC1 (8q24), 26 SRRM2 (16p13.3), 27 MAP2K5 (15q23), 28 –30 HABP2 (10q25.3), 31 –34 19q13 locus (TCO, MYO1F, GRIM19, and TIMM44), 21 –25,35 –39 2q21 locus (NMTC1), 23,25,35,40,41 8p23.1-p22 locus (PTEN), 42 1q21 (fPTC/PRN), 23,25,43,44 BROX (1q41), 45 POT1 (7q31.33), 46 –48 WDR77 (1p13.2), 49 and 6q22 locus, 43 most of these variations only exist in specific study families and have not been validated in other families or cannot be replicated in heredity.
Thus, FNMTC may have significant genetic heterogeneity, with each family having its unique pathogenic gene. Our study identified a novel mutation, PAK4 I417T, for the first time. The mutated form of PAK4 exhibited oncogenic function and distinct mechanisms, indicating its potential as a new diagnostic molecular marker for FNMTC.
Materials and Methods
Ethics statement
This study was approved and conducted by the Ethics Committee of Union Hospital Tongji Medical college, Huazhong University of Science and Technology (2017-S062). All participants were informed of their rights to consent and signed an informed consent form. Animal experiments were approved by the Animal Ethics Committee of Tongji Medical College, Huazhong University of Science and Technology (2021-3247). Our research was carried out in accordance with relevant ethical regulations.
Patients and management
The inclusion criteria for family pedigrees were as follows: (1) Three or more first-degree relatives who have been pathologically diagnosed with non-medullary thyroid cancer; (2) older adult controls: first-degree relatives who were over 50 years old and had no history of tumors; (3) no intermarriage with ethnicities other than Han in the family; (4) no consanguinity between spouses; (5) no clear history of external radiation exposure; and (6) no other genetic diseases.
A five-member affected family was included, with all members undergoing health screening, including ultrasound examination of the thyroid and cervical lymph nodes, as well as serological thyroid function testing. Clinicopathological information was collected during preoperative and postoperative follow-up, with regular reassessment every six months. In addition, peripheral blood leukocyte DNA samples were obtained from validation cohort comprising 31 additional FNMTC patients and 280 sporadic thyroid cancer patients without a family history.
The selection of the validation cohort is described in detail in the Supplementary Methods section of Supplementary Data S1. Please refer to Supplementary Figure S1 and Supplementary Table S1 for the flow diagram, demographic characteristics, and clinical features of the validation cohort.
Peripheral blood whole-exome sequencing
Peripheral blood DNA extraction and whole-exome sequencing (WES) were processed according to the procedures outlined in the Supplementary Methods section of Supplementary Data S1.
Polymerase chain reaction amplification and Sanger sequencing
We obtained DNA fragment primers (Supplementary Data S1) of selected candidate genes using the Primer-Blast website. 50 Purified polymerase chain reaction (PCR) products were sequenced using the BigDye Terminator v3.1 sequencing kit (Thermo ABI) and analyzed on an ABI3730xl sequencer (Applied Biosystems, Foster City, CA).
Selection of candidate genes
Through WES data analysis, shared mutation sites were identified in the exomes of five thyroid cancer patients (I1, II2, II9, II11, and III6) within this specific family. Subsequently, the older adult normal controls (II3, II5), introns, synonymous mutations, and fully homozygous genes were sequentially excluded. The Exome Aggregation Consortium (ExAC) 51 and Genome Aggregation Database (gnomAD) 52 were utilized to exclude mutations with allele frequencies >0.001 in Asians. In silico predictions, including PolyPhen-2, 53 SIFT, 54 and Mutation Taster 55 models, were used to screen candidate genes for the identified variants. Emphasis was placed on genes associated with thyroid and cancer function.
Sanger sequencing was utilized within the family to confirm the co-segregation of the gene with the phenotype. It was also employed among different families to determine whether there are shared susceptibility genes, and for verification in sporadic cases. The Cancer Genome Atlas (TCGA) database 56 was also consulted for additional validation in sporadic cases.
Protein structure model prediction
The domain structure model prediction and gene conservation information were obtained from The Ensembl Genome Browser website. 57 The three-dimensional structure models were generated using The Swiss-Model Server. 58 Protein phosphorylation site prediction is performed using NetPhos Server, 59 whereas DeepSite website 60 is utilized as a binding pocket predictor.
Immunohistochemistry
Tissue samples from FNMTC and SNMTC were collected and fixed. Immunohistochemistry was performed by incubating the samples with primary antibodies (p-PAK4) specific to the target proteins, followed by incubation with secondary antibodies. Please refer to the Supplementary Methods section for detailed information on the sample processing procedures.
Cell culture
The normal human thyroid cell line Nthy-ori-3-1 and the human thyroid cancer cell line TPC-1 were purchased from European Collection of Authenticated Cell Cultures and American Type Culture Collection, respectively. The cells were maintained in RPMI (Viva Cell) or Dulbecco's modified Eagle's medium (HY CEZMBIO) medium supplemented with 10% fetal bovine serum (Viva Cell) at 37°C and 5% CO2 in a cell culture incubator.
Construction and transfection of over-expression lentiviral vectors
Using the Oligobio-LV-003 vector, constructs were made to overexpress wild-type (WT) PAK4, mutant (MUT) PAK4, and control lentiviruses. These constructs were controlled by the PCDH-CMV-MSCEF1-copGFP-T2A-Puro promoter. The transfection efficiency was finally verified by reverse transcription quantitative polymerase chain reaction and Western blotting. The samples were processed according to the procedures outlined in the Supplementary Methods section.
Phenotypic experiments in vitro
Cell immunofluorescence, cell viability assay, colony formation assay, wound healing assay, transwell assay, 5-ethynyl-2-deoxyuridine (EDU) incorporation assay, and flow cytometry were conducted following the procedures outlined in the Supplementary Methods section.
Animal preparation in vivo
Four-week-old female BALB/c nude mice (Beijing Vital River Laboratory Animal Technology Co., Ltd.) were housed in the isolation facility of the experimental center. TPC-1-NC, TPC-1-WT, and TPC-1-MUT cells were collected, washed with phosphate-buffered saline, resuspended in serum-free medium, and made into a suspension of 100 μL per mouse with 1 × 105 cells.
The cell suspension was injected into the tail vein to establish a pulmonary metastasis model. After eight weeks, the mice were sacrificed, the lungs were removed, fixed with 4% paraformaldehyde, and stained with hematoxylin and eosin for histological analysis.
Transcriptome sequencing and analysis
The samples of Nthy-ori-3-1-NC, Nthy-ori-3-1-WT, and Nthy-ori-3-1-MUT cells were used for transcriptome sequencing. The detailed data processing and analysis strategies are described in the Supplementary Methods section.
Immunoprecipitation, chromatin immunoprecipitation, and dual-luciferase reporter assay
TPC-1-WT and TPC-1-MUT cells were used for experiments. The samples were processed following the procedures described in the Supplementary Methods section.
Statistical analysis
SPSS software (version 22.0; IBM Corp., Armonk, NY) was used for statistical analysis. Figures were created by GraphPad Prism software (version 9.1.2; GraphPad Software, La Jolla, CA) and R (4.0.3; R Development Core Team). Following the evaluation of normal distribution and similarity of variances, one-way ANOVA with Tukey's post hoc test was conducted. All experiments were independently repeated at least three times. A p-value <0.05 was considered statistically significant for both tests. All tests were two-sided.
Results
PAK4: c.T1250C: p.I417T identified as FNMTC family candidate gene
We studied a family with five members across three generations affected by papillary thyroid carcinoma. Total thyroidectomy with central or lateral neck dissection was carried out on II2, II9, II11, and III6. Patient I1 declined surgery but underwent fine needle aspiration, which revealed papillary thyroid carcinoma. Our experienced director of thyroid surgery diagnosed the patient with clinically significant thyroid cancer.
During the 79-month follow-up period after diagnosis, distant metastases (lung and brain) were observed in the proband II9. Further, as indicated by the test report, a somatic mutation BRAFV600E was also detected using fluorescence quantitative PCR. The clinical and pathological information of these patients is summarized in Tables 1 and 2.
Clinical Characteristics of Family Members
Ultrasound report results at final follow-up.
ATG, anti-thyroglobulin; ATPO, anti-thyroid peroxidase; fT3, free triiodothyronine; fT4, free thyroxine; TSH, thyrotropin.
Pathological Characteristics of Patients
Using the screening process illustrated in Figure 1a, we identified two candidate gene variants based on their allele frequency and in silico analysis (Table 3). Finally, PAK4: c.T1250C: p.I417T was selected for further study. Its allele frequency was found to be 0.0003 in ExAC_EAS and 0.00023 in gnomAD. Moreover, SIFT, Polyphen2, and MutationTaster analyses all indicated that the variant is potentially damaging.
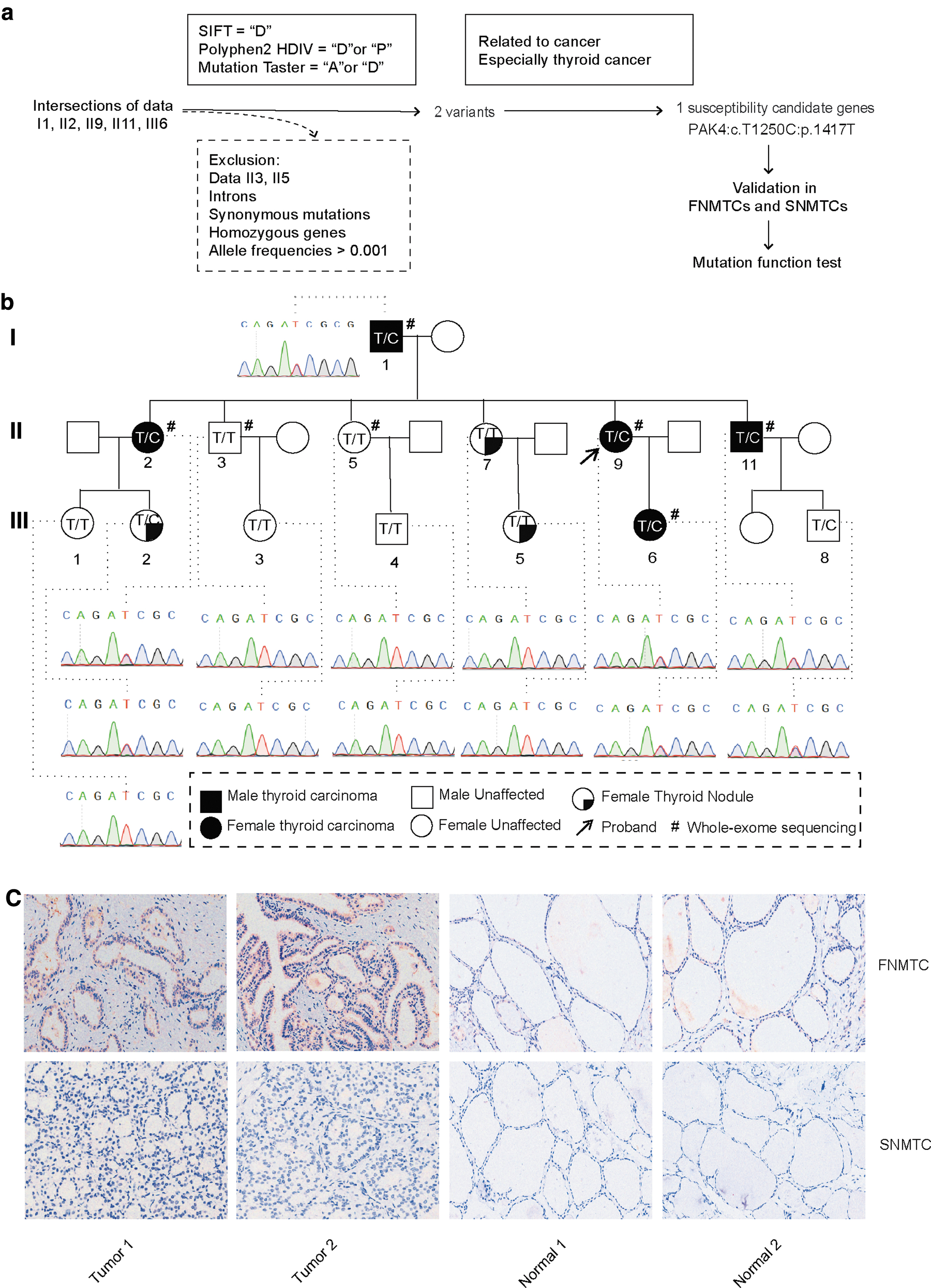
The identification of candidate genes. (
Information of Candidate Genes
ExAC, Exome Aggregation Consortium; gnomAD, Genome Aggregation Database.
The pedigree of the FNMTC family is illustrated in Figure 1b. Sanger sequencing results revealed that II7, III1, III3, III4, and III5 were found to be WT, whereas III2 and III8 carried the PAK4 I417T mutation. Patient III2 has suspicious thyroid nodules detected on a thyroid ultrasound and is currently being closely monitored. Patient III8 is only 10 years old. Hence, further observation is necessary for these two individuals.
The PAK4: c.T1250C: p.I417T mutation was not detected in the additional 280 sporadic cases (0/280) or in the TCGA database (0/481). Among the Sanger sequencing data from 31 other FNMTC patients, one patient was found to carry the same mutation (1/31). Regrettably, in the newly identified pedigree, only the son from a three-member affected family was able to participate in the experiment and provide peripheral blood, as the mother has passed away and the other daughter is unable to participate for personal reasons.
IHC (Fig. 1c) results of thyroid cancer tissues and adjacent non-cancerous tissues revealed that p-PAK4 protein was expressed in both the cytoplasm and nucleus in FNMTC (representing the mutant PAK4 genotype), whereas no significant expression was detected in patients with SNMTC (WT PAK4 genotype).
Protein functional prediction of PAK4 I417T
The PAK4 protein's domain structure consists of a protein-binding domain and a catalytic domain, as shown in Figure 2a. The I417T mutation occurs at the catalytic site in exon 6 and is located in a conserved region of the gene. Three-dimensional structure prediction (Fig. 2b) reveals that the amino acid change caused by this missense mutation is situated on a β-helix. The mutation leads to reduced hydrophobicity, which may contribute to increased protein instability.

Protein structure prediction. (
NetPho prediction (Fig. 2c) indicates that the I417T mutation introduces a phosphorylation site with a SCORE >0.05, indicating potential alterations in protein phosphorylation. In addition, DeepSite predictions (Fig. 2d) demonstrate that the mutation of amino acid 417T creates a nearby binding site (id2). The presence of the mutated Threonine (Thr) at position 417, forming an intramolecular hydrogen bond with the neighboring Asparagine (Asn) at position 413, leads to protein structural instability. This implies that the mutated residue may interact with other ligands, potentially affecting the protein's function and stability.
PAK4 I417T promotes phosphorylated PAK4 nuclear translocation and expression
Cell lines overexpressing WT and MUT PAK4 were successfully generated using Nthy-ori-3-1 and TPC-1 cells, as confirmed by mRNA expression, and protein expression analyses (Fig. 3a–d).
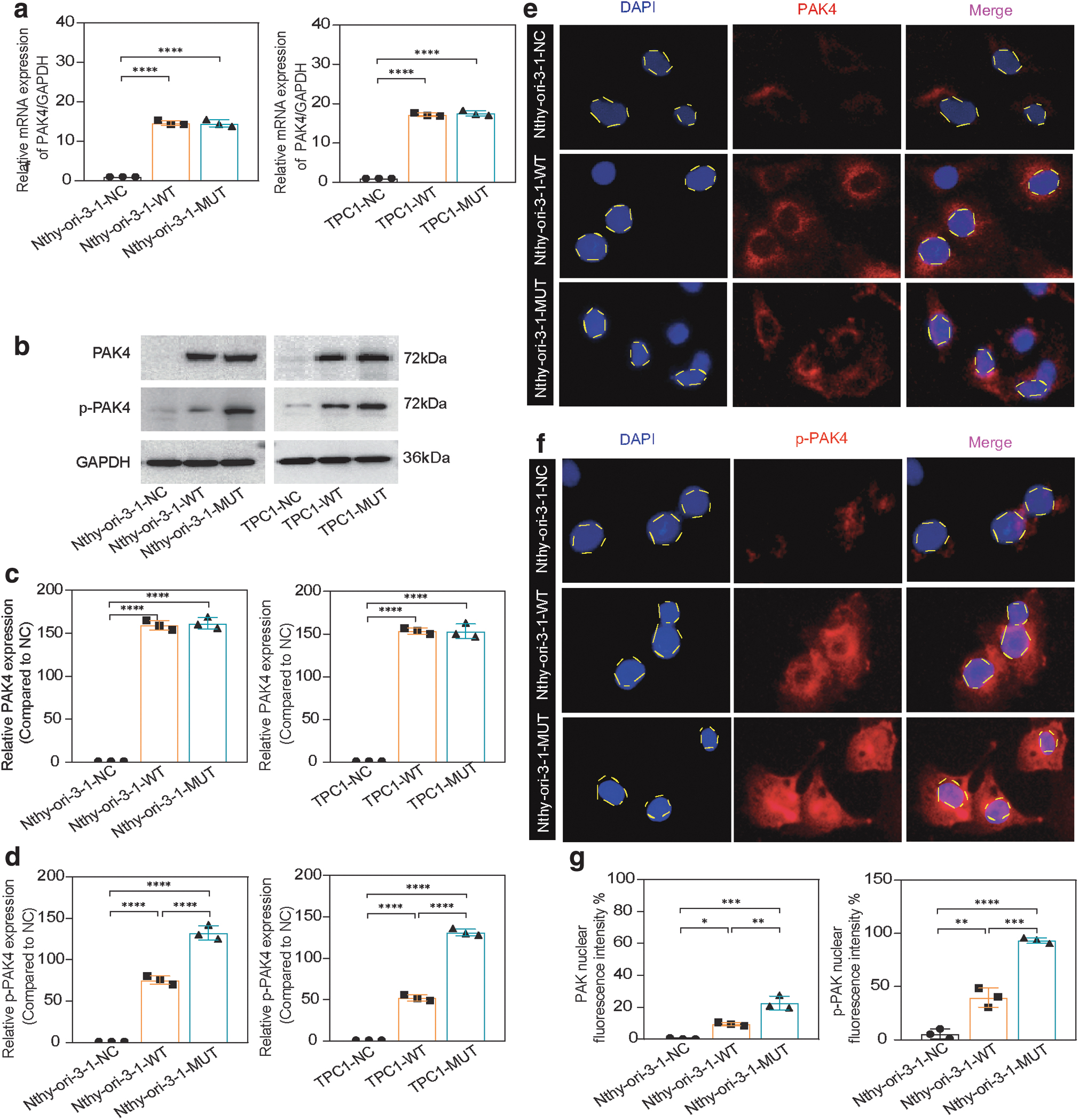
Changes in protein localization and expression after mutation. (
Cell immunofluorescence localization revealed continuous expression of PAK4 in the cytoplasm, whereas phosphorylated PAK4 was primarily localized to the cytoplasm and nucleus (Fig. 3e–f). The MUT group displayed higher nuclear fluorescence intensity and protein expression levels of p-PAK compared with the WT group, and these differences were statistically significant (Fig. 3d, g).
PAK4 I417T demonstrates enhanced proliferation, invasiveness, accelerated cell division, and inhibition of cell apoptosis in vitro
Overexpression of PAK4 MUT significantly enhances cell proliferation compared with the WT, as shown by cloning formation (Fig. 4a,4b), CCK-8 proliferation (Fig. 4c), and EDU assays (Fig. 4d, e). In addition, MUT-overexpressing cells exhibit increased invasiveness and migratory abilities compared with the WT group in transwell (Fig. 4f, g) and scratch assays (Fig. 4h, i).
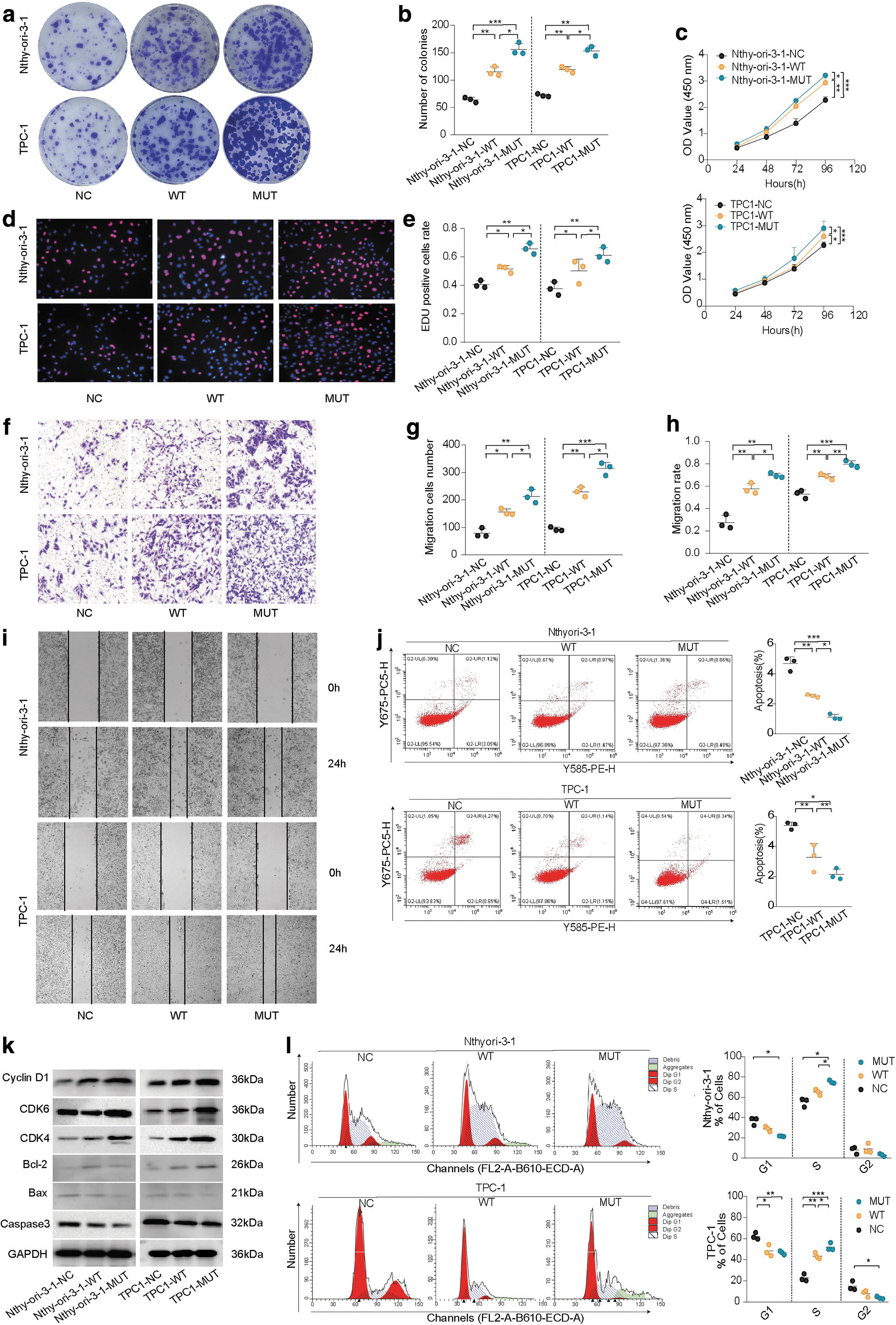
Functional validation of WT and mutant PAK4 in vitro. (
Overexpression of MUT leads to a reduction in the proportion of cells in the G1 phase and an increase in the S phase, resulting in accelerated cell division (Fig. 4j). This effect is likely due to the upregulation of Cyclin D1 and CDK4/6 expression (Fig. 4k). Both the WT and MUT groups exhibit inhibition of cell apoptosis, with MUT showing a stronger and statistically significant effect compared with WT (Fig. 4l). PAK4 overexpression promotes the expression of Bcl-2, while inhibiting the expression of Bax and caspase-3, thereby suppressing cell apoptosis, particularly in the MUT group (Fig. 4k).
PAK4 I417T demonstrates a significant ability for metastasis in vivo
The MUT group demonstrated a higher incidence of lung metastases in comparison to the NC and WT groups (NC:WT:MUT = 1.75:3.75:7, mean). The statistical analysis further confirmed the presence of a significant disparity (Fig. 5a).
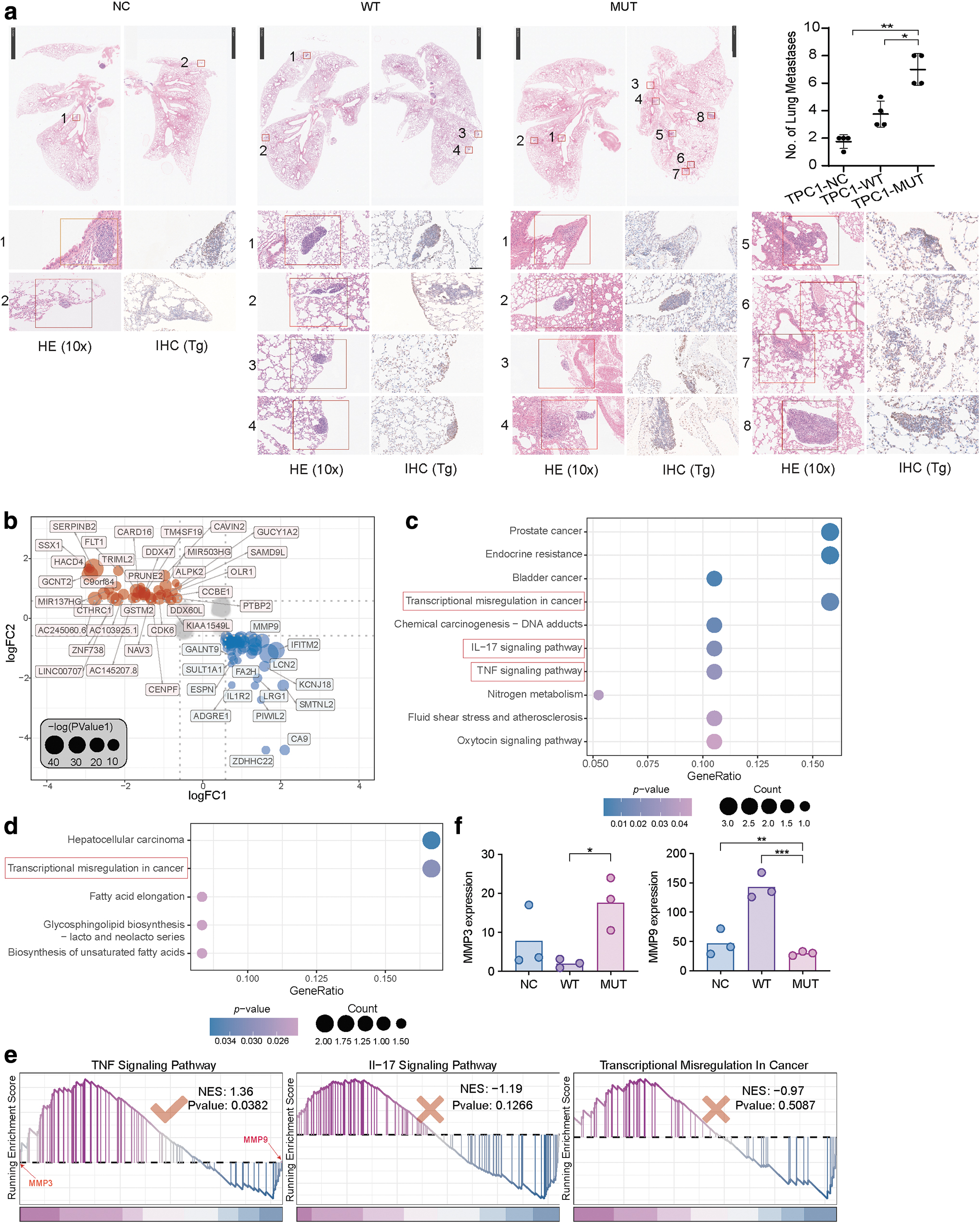
Tumor lung metastasis in vivo and transcriptome sequencing identifies differences between WT and mutant PAK4. (
PAK4 WT and MUT modulate the expression of different matrix metalloproteinase families through tumor necrosis factor signaling pathway
A total of 86 differential expression genes (DEGs) (Fig. 5b) were gained according to the DEGs screening strategy in the method section. The Kyoto Encyclopedia of Genes and Genomes enrichment analysis results showed that the DEGs that were upregulated in WT versus NC and downregulated in MUT versus NC were enriched in some cancer-related signaling pathways, particularly those related to thyroid cancer, such as transcriptional misregulation, interleukin-17, and tumor necrosis factor (TNF) pathways (Fig. 5c), whereas differentially expressed genes that were downregulated in WT versus NC and upregulated in MUT versus NC were enriched in transcriptional misregulation signaling pathways (Fig. 5d).
GSEA analysis between the WT and MUT groups revealed that only the TNF signaling pathway showed significant enrichment (Fig. 5e). In the TNF signaling pathway, the differentially expressed gene MMP9 (Fig. 5b) is involved, along with other genes in the same family such as MMP3. Further, MMP3 was significantly upregulated in the MUT group compared with the NC group (Fig. 5f). In addition, GSEA analysis showed that MMP3 and MMP9 were respectively at the front and end of the log2FoldChange ranked list, suggesting their significant roles in the MUT and WT groups, respectively (Fig. 5e).
The levels of MMP9 were higher in the WT group compared with the MUT group, whereas the expression of MMP3 was lower in the WT group, consistent with the transcriptome sequencing results (Fig. 6g).
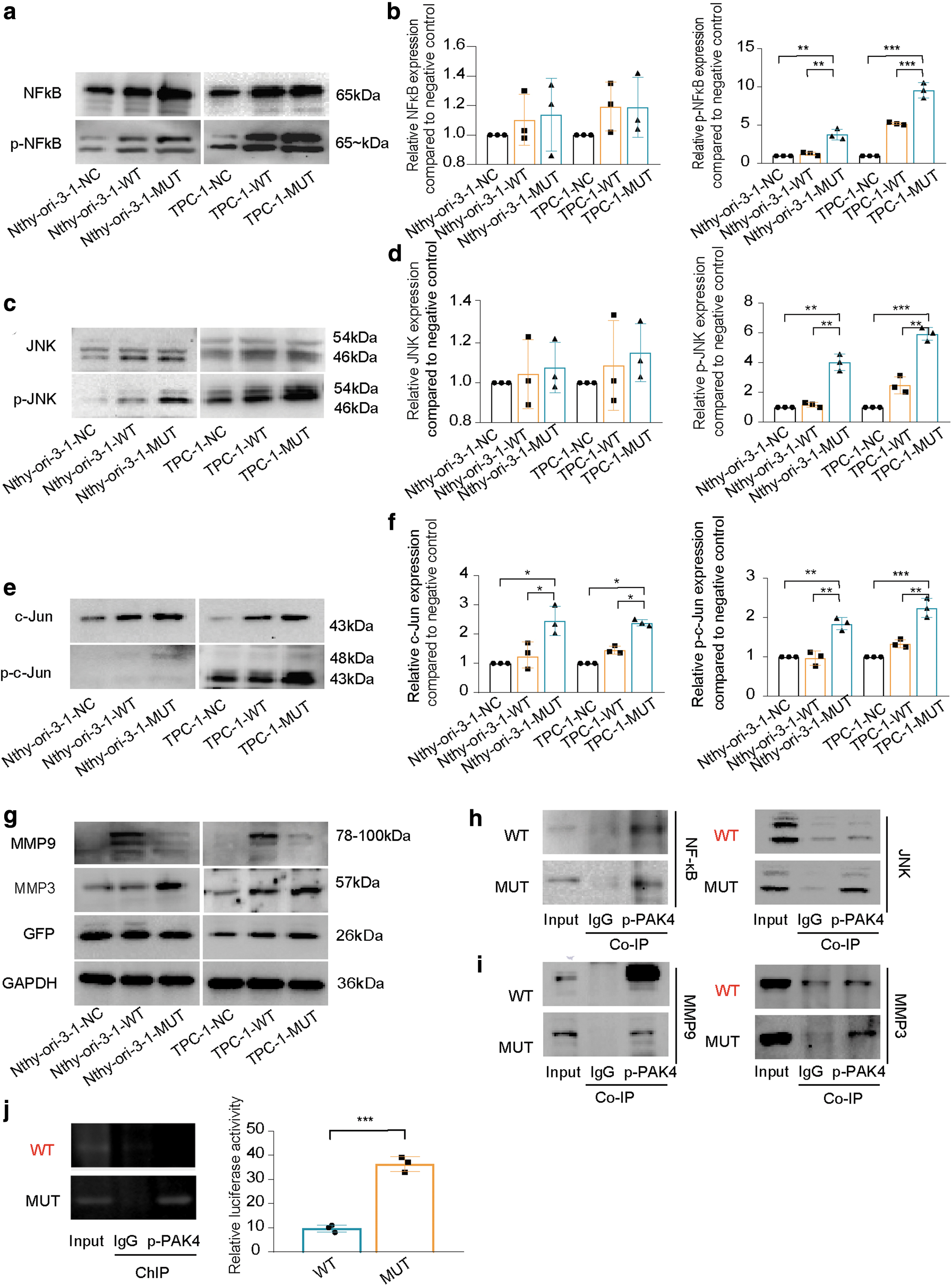
Changes in the expression of related proteins through the TNF signaling pathway. (
PAK4 I417T activates the signaling pathway of the TNF pathway by promoting phosphorylation levels PAK4, JNK, NFκB, and c-Jun
MUT enhanced the protein phosphorylation levels of PAK4, JNK, and NFκB (Fig. 6a–d). Due to JNK activation, downstream c-Jun was also activated, resulting in a protein kinase cascade reaction (Fig. 6e–f).
The mutant PAK4 can enhance the oncogenic effect by interacting with proteins and regulating MMP3 promoter activity
Both p-PAK4 I417 and T417 were found to directly interact with NFκB, facilitating the transfer of phosphate groups. However, only T417 can directly bind to JNK (Fig. 6h).
Further, p-PAK4 T417 exhibited direct interactions with both MMP3 and MMP9 in the nucleus, leading to simultaneous promotion of their expression. However, p-PAK4 I417 only displayed direct interaction with MMP9 (Fig. 6i).
The chromatin immunoprecipitation experiments and dual luciferase assays (Fig. 6j) provided further evidence that p-PAK4, acting as a transcription factor, can bind to and increased activity of the MMP3 promoter in the MUT group.
Discussion
This study explores the genetic susceptibility of FNMTC starting from one family. At the same time, the variant PAK4: c.T1250C: p.I417T was not detected in sporadic thyroid cancer. This observation suggests that there may be differences in the underlying mechanisms of disease onset between individuals with FNMTC and SNMTC, consistent with the genetic characteristics of FNMTC.
Moreover, the presence of the same mutation in one patient from a different pedigree, combined with the enhanced oncogenic function exhibited by PAK4: c.T1250C: p.I417T, indicates its potential as a novel susceptibility gene for FNMTC.
In previous studies, we have observed significant genetic heterogeneity in FNMTC, with many findings specific to individual families. Some identified susceptible genes are related to FNMTC's specific combined diseases. For example, 1q21 (fPTC/PRN) is a specific locus for FNMTC combined papillary renal neoplasms (PRN) 44 ; the 2q21 (NMTC1) locus is associated with follicular variant papillary thyroid carcinoma (fvPTC) 40 ; TTF-1 may be a susceptible gene for FNMTC combined multinodular goiter (MNG). 61
In our study, patients II2, II9, II11, and III6 exhibited elevated levels of anti-thyroglobulin or anti-thyroid peroxidase (ATPO), and their pathological results indicated Hashimoto's thyroiditis. However, PAK4 WT family member II5 also showed elevated levels of ATPO without thyroid cancer. Therefore, the role of mutated PAK4 in the development of FNMTC associated with Hashimoto's disease remains unclear, but it shows potential as a specific susceptible gene. And further research is needed.
Our research findings indicate that the PAK4: c.T1250C: p.I417T mutation is associated with more aggressive malignant characteristics. We analyze the reasons for the enhanced functionality of the oncogene. First, starting from the fundamental change in protein conformation, it is possible that the mutation of the amino acid at position 417 to Thr results in the formation of an intramolecular hydrogen bond with the nearby Asn at position 413.
As a result, it leads to structural instability of the protein. This suggests that the mutated residue may form new interactions with other molecules or ligands, thus affecting the function or stability of the protein. In addition, the substitution of Thr at position 417 leads to increased phosphorylation and binding sites in the PAK4 protein.
Changes in protein structure often affect protein function. Further, combining transcriptomic data, we have discovered that PAK4 I417T can promote phosphorylated PAK4 nuclear translocation and expression. It also promotes the phosphorylation levels of JNK, NFκB, and c-Jun, activating the TNF pathway (Fig. 7). This conclusion validates our prediction.
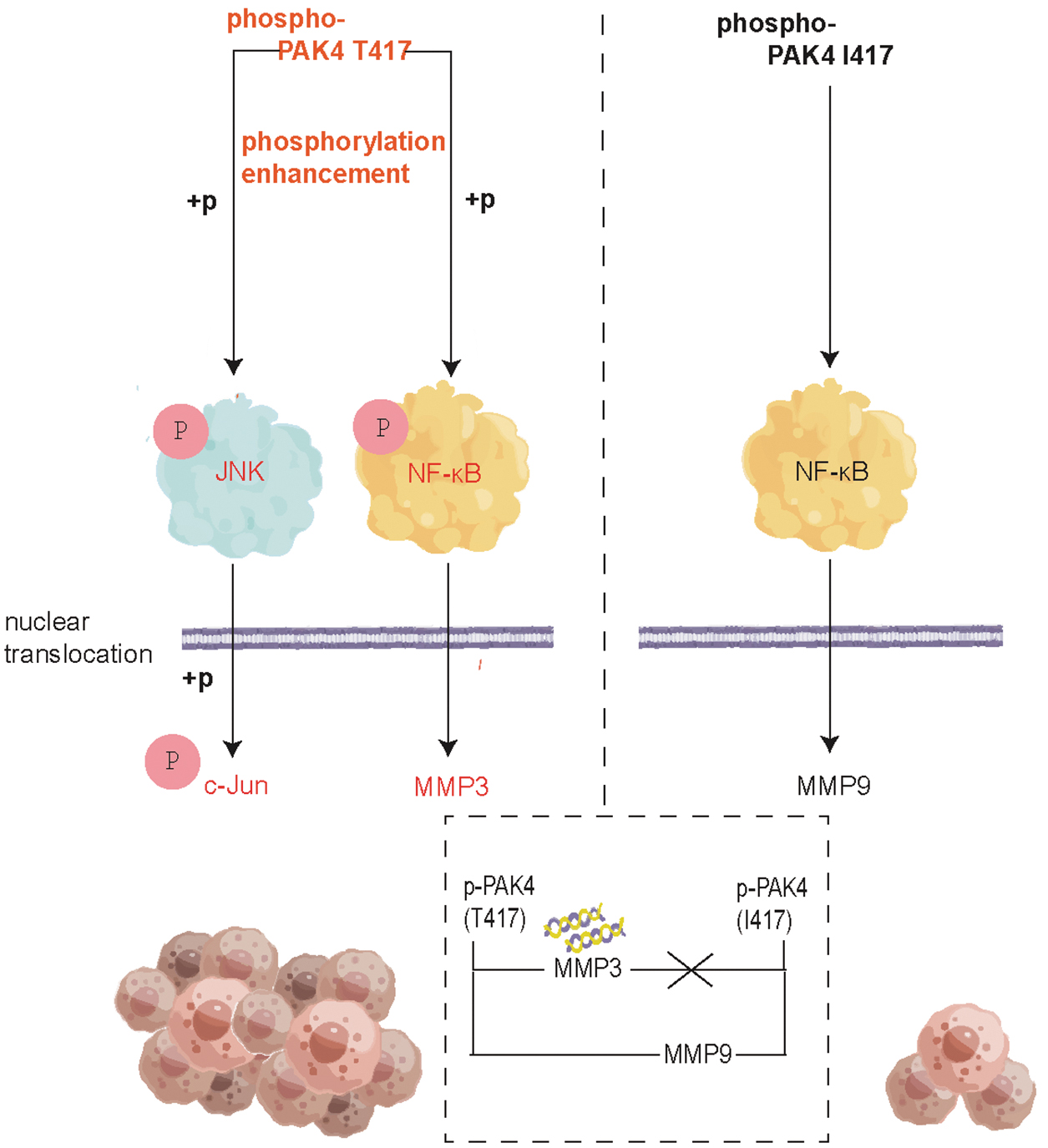
Changes in the mechanism of mutant PAK4 in the TNF signaling pathway. The diagram mechanisms of WT and mutant PAK4 in the TNF signaling pathway (by Figdraw).
The downstream of the TNF signaling pathway involves the matrix metalloproteinases (MMPs) family. Currently, studies have shown that PAK4 can affect the expression of MMP9 in patients with adenomyosis, colorectal cancer, and breast cancer, 62 –64 but its mechanism of action has not been explained and has not been verified in thyroid cancer. Our study found that both the mutant and WT forms can promote MMP9 expression through direct binding, but this effect is more pronounced in the WT form.
There are also studies showing the association between MMP3 gene and the occurrence of thyroid cancer, 65,66 but there is currently no study exploring the association between PAK4 and MMP3. In our verification experiment, PAK4 I417T not only has the ability to directly bind to the promoter region of MMP3 but also exerts an influence on its transcription and expression. This functionality has not been observed in the WT PAK4 (Fig. 7).
The limitations of our study include the lack of comprehensive somatic mutation analysis, as we only tested for BRAF mutations in patient II9 and did not screen for other commonly observed mutations such as TERT promoter or TP53. This limited evaluation may have missed important mutations that contribute to disease progression. Another limitation is the absence of a papillary thyroid carcinoma cell line with the same BRAFV600E mutation as patient II9.
This introduces variability in the results and may limit generalizability. Future studies should broaden somatic mutation analyses and utilize cell lines with matched genotypes to gain a comprehensive understanding of the molecular landscape.
Finally, the genetic causal relationship model of FNMTC is complex, with unique pathogenesis likely present in different affected families. In one family, PAK4: c.T1250C: p.I417T could represent a potential susceptibility gene for FNMTC. However, more pedigrees and replication studies are needed to confirm this finding.
Availability of Data and Materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Footnotes
Authors' Contributions
Y.-J.J.: Data curation, Conceptualization, Funding acquisition, Writing—original draft, and Formal analysis. Y.X.: Validation, Software, Data curation, and Formal analysis. Y.-X.H.: Writing—original draft, Validation, and Data curation. Z.-J.H.: Visualization, Resources, and Data curation. A.-Y.G.: Data curation, Resources, and Writing—review and editing. T.H.: Methodology, Investigation, Data curation, Conceptualization, Writing—review and editing, and Funding acquisition.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by “the Fundamental Research Funds for the Central Universities” and “Key Program of Natural Science Foundation of Hubei Province” (Grant Nos. [No. 2020JYCXJJ008] and [No. 2021BCA142]).
Supplementary Material
Supplementary Data S1