
Abstract
Background:
Papillary thyroid cancer (PTC) with the BRAFV600E mutation is associated with a poorer prognosis. BRAF inhibitors may demonstrate limited efficacy due to emerging drug resistance. The Warburg effect may have cancer therapeutic implications. It is not known if the BRAFV600E mutation is associated with altered glucose metabolism in PTC.
Methods:
This study examined the effect of BRAFV600E and dynamin-related protein 1 (DRP1) on various cellular processes in PTC cells, including cell proliferation, migration, invasion, mitochondrial fission, glucose metabolism, reactive oxygen species (ROS) generation, and apoptosis. We used RT-qPCR to assess the expression of key glycolytic enzymes in thyroid cancer tissues. Additionally, the regulatory interaction between BRAFV600E and DRP1 was investigated through Western blot and immunohistochemical staining. We further evaluated the impact of DRP1 in PTC and the inhibitory effects of dabrafenib and 2-deoxy-
Results:
We found that the BRAFV600E mutation significantly augments aerobic glycolysis while suppressing oxidative phosphorylation in PTC. We identified the BRAFV600E/p-ERK/p-DRP1(Ser616) signaling pathway as a critical mediator in PTC progression. First, the BRAFV600E/p-ERK/p-DRP1(Ser616) signaling pathway enhances cell proliferation by upregulating hexokinase 2 expression and thereby increasing aerobic glycolysis. Second, it inhibits apoptosis by promoting mitochondrial fission and reducing ROS levels. Moreover, we demonstrated that the combination therapy of 2-DG and dabrafenib markedly impedes the progression of BRAFV600E-positive PTC.
Conclusion:
The BRAFV600E/p-ERK/p-DRP1(Ser616) signaling pathway plays a pivotal role in glucose metabolism reprogramming, contributing to the aggressiveness and progression of BRAFV600E-positive PTC. Our findings suggest that a combined therapeutic approach using 2-DG and dabrafenib has the potential to improve the outcome of PTC patients with BRAFV600E.
Introduction
Thyroid cancer (TC) is the most prevalent tumor in the endocrine system. Papillary TC (PTC) is the predominant histological subtype of TC. The BRAFV600E mutation emerges as the most frequent oncogenic alteration, present in approximately 45–80% of PTC. 1,2 This mutation is strongly associated with adverse clinical outcomes, including increased recurrence rates, lymph node metastasis, advanced tumor stages, poorer prognosis, and increased cancer-related mortality. 3,4 Additionally, BRAFV600E leads to increased resistance to postoperative I131 therapy. 5 However, BRAF inhibitors (BRAFi) such as vemurafenib and dabrafenib have shown limited effectiveness due to the emergence of resistance and severe side effects. 6,7 Therefore, it is crucial to identify novel therapeutic targets for the treatment of BRAFV600E-mutated PTC.
The evolving understanding of tumor biology has acknowledged metabolic reprogramming as a novel cancer hallmark. 8,9 The Warburg effect, a phenomenon where tumor cells predominantly rely on glycolysis for energy even under aerobic conditions, is a well-reported example of this metabolic shift. 10 Therapeutics targeting the Warburg effect have shown effectiveness in halting the progression of various cancers. 11 –13 Notably, the BRAFV600E has been related to augmented aerobic glycolysis in cancers such as melanoma, colorectal, and bladder cancer. 14 –16 Previous research suggests that BRAFV600E-mutated differentiated TCs exhibit significantly higher 18F-FDG uptake compared with wild-type BRAF (BRAFWT) tumors. 17 However, the precise relationship between BRAFV600E and glucose metabolism in PTC remains unknown.
Decreased mitochondrial respiration and increased aerobic glycolysis have been reported in various cancers. 18,19 Central to the regulation of cellular metabolism, and thereby playing a pivotal role in cancer development and progression, is the balance between mitochondrial fission and fusion as well as overall mitochondrial function. 20,21 Emerging evidence underscores that mitochondrial fission facilitates cell proliferation and metastasis. 22 Conversely, mitochondrial fusion exhibits tumor-suppressive effects in various cancers. 23,24 Notably, BRAFV600E has been identified to augment mitochondrial fission. A previous study demonstrated that inhibiting the MAPK pathway can suppress mitochondrial fission and biogenesis in melanoma, suggesting a potential therapeutic target for cancers with the BRAFV600E mutation. 25
In light of these findings, it becomes imperative to delineate the relationship between BRAFV600E, glucose metabolism, and mitochondrial dynamics, especially given the challenging therapeutic effects and poor prognosis associated with BRAFV600E-mutated PTC. Our study identifies a strong association between BRAFV600E and increased aerobic glycolysis, as well as decreased oxidative phosphorylation (OXPHOS) in PTC. We have systematically investigated the role and underlying mechanisms of BRAFV600E in the regulation of mitochondrial dynamics and metabolic reprogramming in PTC cells. Our findings reveal that a combination therapy of 2-deoxy-
Materials and Methods
Public database analysis
Gene set enrichment analysis (GSEA) was conducted to assess gene expression differences between PTCs with BRAFWT and those with the BRAFV600E mutation.
Clinical sample analysis
Eighty pairs of tumor and peritumor tissues were collected to extract RNA. These specimens were obtained from PTC patients who underwent primary surgery, including thyroidectomy and routine lymph node dissection (central and/or lateral lymph node dissection) at the Department of Head and Neck Surgery, Fudan University Shanghai Cancer Center (FUSCC), from March 2018 to June 2018, with follow-up until June 2022. No patients received thyroid surgery, radioiodine, or radiotherapy before the TC surgery. RT-qPCR was utilized to compare the expression levels of glycolysis-related enzymes in PTC versus peritumoral tissue. A total of 155 formalin-fixed, paraffin-embedded tissue PTC samples were collected to construct a tissue microarray (TMA). These samples were obtained from 97 patients who underwent primary PTC surgery at the Department of Head and Neck Surgery, FUSCC, between January 2010 and January 2015, with follow-up until June 2022. No patients received thyroid surgery, radioiodine, or radiotherapy before the TC surgery. The staging for T1/T2 PTC was based on the eighth edition of the AJCC staging system. 26 Locally advanced PTC (LA-PTC) was defined as PTC with invasion into muscles, blood vessels, nerves, esophagus, and trachea. BRAF status was ascertained by an amplification refractory mutation system based on an ultrasound-guided fine-needle biopsy. TMAs were stained by immunohistochemistry (IHC). The mean optical density (MOD) was employed for semiquantitative analysis of protein expression levels. The expression of phosphorylated dynamin-related protein 1 (DRP1) (Ser616) was compared among peritumoral tissue, T1/T2-stage PTC, and LA-PTC. Comparisons were also made between BRAFWT PTC and BRAFV600E PTC. Detailed immunohistochemical staining and evaluation methods are shown in Supplementary Data S1.
The study was approved by the Medical Ethics Committee of Fudan University Cancer Hospital (No. 050432-4-1911D), and patients provided informed consent to use their tissues in the study.
In vitro experiments
Three TC cell lines were used: TPC-1 (BRAF mutation negative), BCPAP (BRAFV600E positive), and K1 (BRAFV600E positive). DRP1 was knocked down by lentivirus–shRNA transfection. DRP1 was reintroduced by plasmid transfection. BRAFV600E was overexpressed by lentivirus–plasmid transfection. BRAFV600E was inhibited by dabrafenib. Cell viability, colony formation, migration, and invasion assays were conducted to assess the progression of PTC in vitro. Glycolysis was evaluated through the extracellular acidification rate (ECAR) and lactate production (LP) assays. OXPHOS was assessed via the oxygen consumption rate (OCR) assay. Mitochondrial morphology was analyzed using transmission electron microscopy (TEM). Cellular reactive oxygen species (ROS) production and apoptosis were assessed by test kits. The effects of dabrafenib, 2-DG, and their combined application were evaluated in vitro. Detailed experimental methods are presented in Supplementary Data S1.
In vivo experiments
Female BALB/C nude mice aged 4–5 weeks (purchased from Shanghai SLAC ANIMAL Co., Ltd.) were used to establish a subcutaneous xenograft model. Dead and nontumorous mice were excluded from the experiment. One researcher implanted tumor cells into the mice. The staff in the animal room who were unaware of the group allocation were in charge of feeding, cage allocation, and measurement. The research was approved by the Experimental Animal Care and Use Committee of Fudan University Cancer Hospital (no. FUSCC-IACUC-S2023-0506).
Knockdown of DRP1
Twelve mice were randomly divided into two groups using a random number generator in Excel software (version 2019; Microsoft, Redmond, WA): shNC and shDRP1. Each mouse was implanted with K1 cells on the right side. Tumor volumes were monitored weekly for 4 weeks. There was one mouse in each of the shNC group and the shDRP1 group, and no subcutaneous tumors formed.
Drug experiments
Twelve mice were randomly divided into four groups using a random number generator: phosphate buffered saline (PBS), dabrafenib (30 mg/kg for each mouse), 2-DG (250 mg/kg for each mouse), and a combination of dabrafenib and 2-DG. Each mouse was implanted with BCPAP cells on both sides. Treatment began when tumor volumes reached ∼ 1500 mm3. Body weight and tumor volume were monitored weekly for 3 weeks.
Statistical analysis
Continuous variables were presented as mean ± standard error of the mean, and classified variable results were shown as frequency and percentage. Paired t and independent t tests were used to compare continuous variables between two groups. Pearson’s χ 2 test and Fisher’s exact test were used for categorical variables. A p value below 0.05 was deemed statistically significant. Data analysis and visualization were performed using SPSS 22.0 and GraphPad Prism 6.0 software on the Windows platform.
Results
Clinical sample
In 80 pairs of samples for RNA extraction, 43 cases carried no BRAF mutation, and 37 cases carried the BRAFV600E mutation. The median follow-up time was 49 months (interquartile range 2 months). Patients’ demographic and clinical characteristics are described in Table 1. In total, 155 TMA samples were subjected to IHC, including 45 PTC in the T1/T2 stage, 48 LA-PTC, and 62 normal thyroid tissues. Out of 97 patients, 30 cases carried no BRAF mutation, 33 cases carried the BRAFV600E mutation, and the remaining BRAF status was unknown. The median follow-up duration was 111 months (interquartile range 27 months). Patients’ information is given in Table 2. A patient flow diagram is shown in Figure 1.
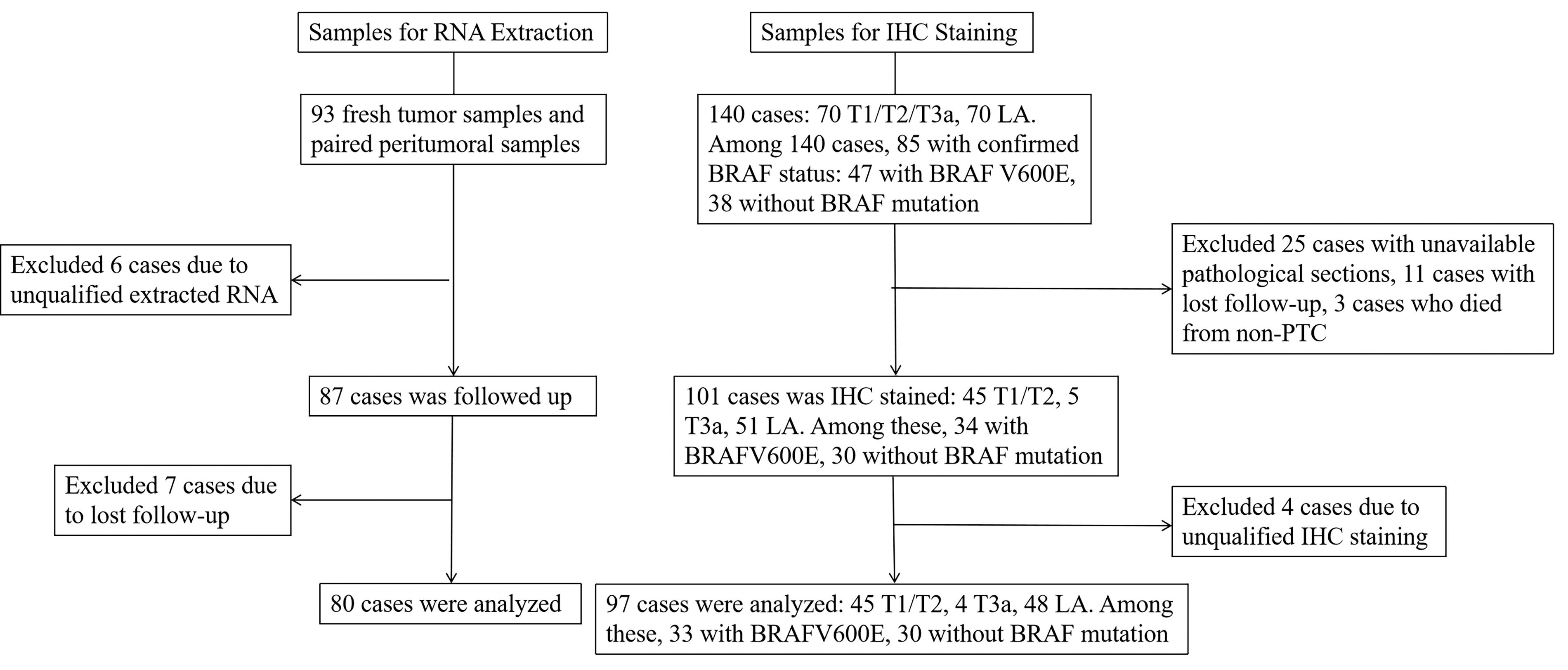
Patient flow diagram. IHC, immunohistochemistry; LA, locally advanced.
Demographic and Clinical Characteristics of Patients (80 Pairs of Tumors and Peritumors)
CLNM, central lymph node metastasis; LLNM, lateral lymph node metastasis; PTC, papillary thyroid cancer.
Demographic and Clinical Characteristics of Patients (Tissue Microarray)
CLNM, central lymph node metastasis; LLNM, lateral lymph node metastasis; PTC, papillary thyroid cancer.
BRAFV600E regulates malignant biological behaviors and glucose metabolism reprogramming in PTC
In vitro experiments demonstrated that BRAFV600E accelerates malignant biological behaviors in PTC (Supplementary Fig. S1). GSEA indicated a notable downregulation in genes associated with respiratory electron transport and the citric acid cycle in BRAFV600E-mutated PTC (Fig. 2A). The expression levels of key glycolytic enzymes, such as LDHA (p = 0.031), SCL2A1 (p = 0.004), and hexokinase 2 (HK2) (p < 0.001), were elevated in PTC compared with peritumor tissue (Fig. 2B). LDHA (p = 0.039) and HK2 (p = 0.014) levels were higher in BRAFV600E-mutated PTC compared with BRAFWT counterparts (Fig. 2C). BRAFV600E was overexpressed in TPC-1 cells, while BCPAP and K1 cells were treated with dabrafenib to inhibit BRAFV600E (Fig. 2D). We observed an increase in ECAR and LP (p < 0.001), along with a decrease in maximal OCR in TPC-1-BRAFV600E cells (Fig. 2E). No significant changes were observed in ECAR, OCR, and LP in TPC-1 cells treated with dabrafenib (Fig. 2F). Dabrafenib treatment reduced ECAR in BCPAP (Fig. 2G) and K1 cells (Fig. 2H). LP significantly decreased in BCPAP when the concentration of dabrafenib reached 1000 nM (p < 0.001). LP significantly decreased in K1 when the concentration of dabrafenib reached 100 nM (p < 0.001). Conversely, OCR increased in dabrafenib-treated BCPAP (Fig. 2G) and K1 cells (Fig. 2H). These findings suggest that BRAFV600E enhances glycolysis and suppresses OXPHOS in PTC.

The impact of BRAFV600E on glucose metabolism reprogramming in PTC.
BRAFV600E regulates mitochondrial fission, ROS production, and apoptosis in PTC
TEM analysis revealed an increase in the number of mitochondria per cell in BRAFV600E-overexpressed TPC-1 and a decrease in BCPAP and K1 treated with 1000 nM dabrafenib. The length of mitochondria was shorter in BRAFV600E-overexpressed TPC-1 and longer in dabrafenib-treated BCPAP and K1 (Fig. 3A). The results indicate that BRAFV600E promotes mitochondrial fission in PTC.
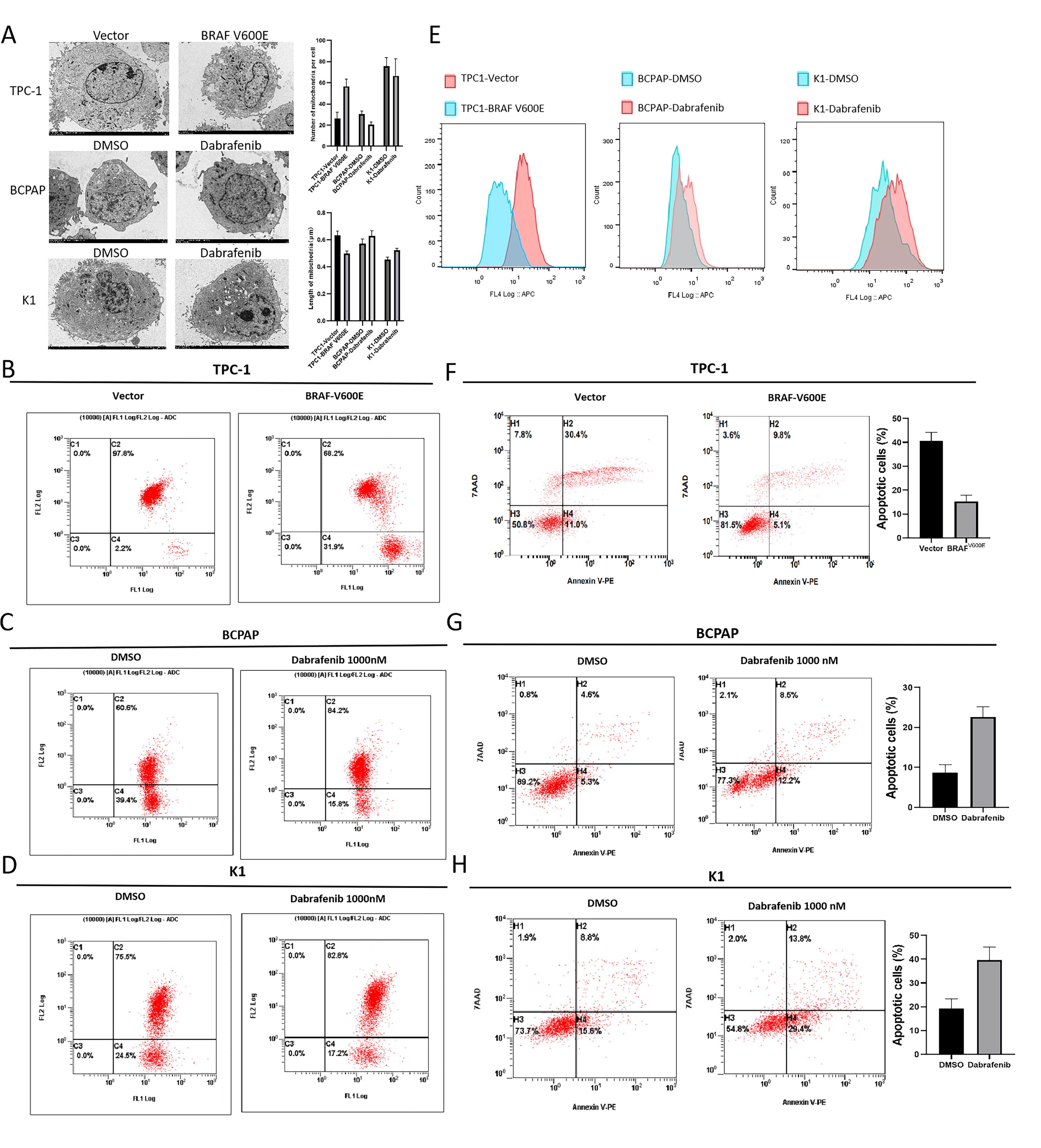
The impact of BRAFV600E on mitochondrial fission/fusion, ROS production, and apoptosis in PTC cells.
JC-1 dye assays showed a reduced proportion of cells with high mitochondrial membrane potential in BRAFV600E-overexpressed TPC-1 cells (Fig. 3B). In contrast, dabrafenib-treated BCPAP (Fig. 3C) and K1 (Fig. 3D) exhibited an increased proportion of cells with high mitochondrial membrane potential, suggesting impaired mitochondrial membrane stability caused by BRAFV600E. Cellular ROS levels decreased in TPC-1 cells overexpressing BRAFV600E and increased in BCPAP and K1 treated with dabrafenib (Fig. 3E). Apoptosis rates decreased in BRAFV600E-overexpressed TPC-1 (Fig. 3F) cells and increased in dabrafenib-treated BCPAP (Fig. 3G) and K1 cells (Fig. 3H).
BRAFV600E/p-ERK/p-DRP1(Ser616) signaling pathway in PTC
Western blot (WB) analysis revealed that both p-ERK and p-DRP1(Ser616) were upregulated in BRAFV600E-overexpressed TPC-1 and HEK-293 cells and downregulated in dabrafenib-treated BCPAP and K1 cells (Fig. 4A). Moreover, Figure 4B shows that the MOD of p-DRP1(Ser616) was significantly elevated in PTC with BRAFV600E (mean: 0.192 [CI 0.190–0.193]) compared with PTC with BRAFWT (mean: 0.185 [CI 0.183–0.187]). These results suggest the presence of a BRAFV600E/p-ERK/p-DRP1(Ser616) signaling pathway in PTC.
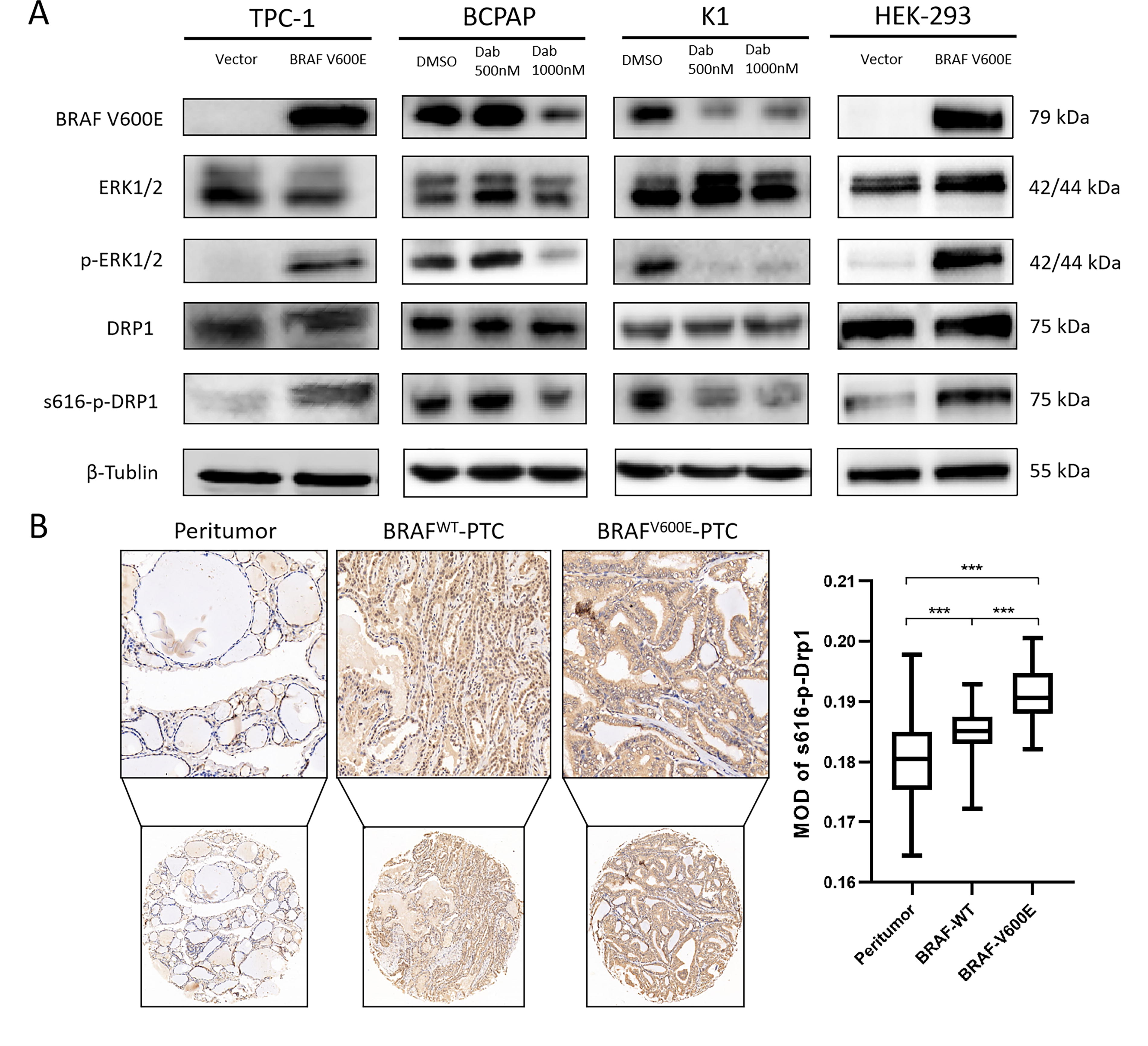
BRAFV600E/p-ERK/p-DRP1(Ser616) signaling pathway in PTC.
DRP1 promotes the progression of BRAFV600E-muted PTC
DRP1 was knocked down in BCPAP, K1, and TPC-1 cells (Fig. 5A). As shown in Figure 5B, the colonies of BCPAP and K1 cells were significantly reduced after DRP1 knockdown (p < 0.001) but were rescued after the reintroduction of DRP1, while the colonies of TPC-1 cells were slightly reduced (p = 0.035) and increased after the reintroduction of DRP1 (p = 0.266). In Figure 5C, the cell viability of BCPAP and K1 decreased and was rescued after the reintroduction of DRP1, while the cell viability of TPC-1 changed little. In Figure 5D, the cell migration of BCPAP was significantly weakened (p < 0.001) and was rescued after the reintroduction of DRP1; the cell migration of K1 was slightly weakened and increased after the reintroduction of DRP1 (p = 0.002). TPC-1 did not show significant changes. In Figure 5E, the cell invasion of BCPAP (p < 0.001) and K1 (p = 0.011) was reduced and rescued after the reintroduction of DRP1, while TPC-1 showed less change. Figure 5F shows that with DRP1 knockdown, the volume of subcutaneous xenograft tumors in mice was significantly smaller than that in the negative control group (p = 0.008). Additionally, the MOD of p-DRP1(Ser616) in LA-PTC (mean: 0.189 [CI 0.188–0.191]) was higher than in T1/T2-PTC (mean: 0.186 [CI 0.184–0.187]), and both were elevated compared with peritumoral tissue (mean: 0.181 [CI 0.180–0.183]) (Fig. 5G). These findings support the role of the BRAFV600E/p-ERK/p-DRP1(Ser616) pathway in promoting PTC progression.
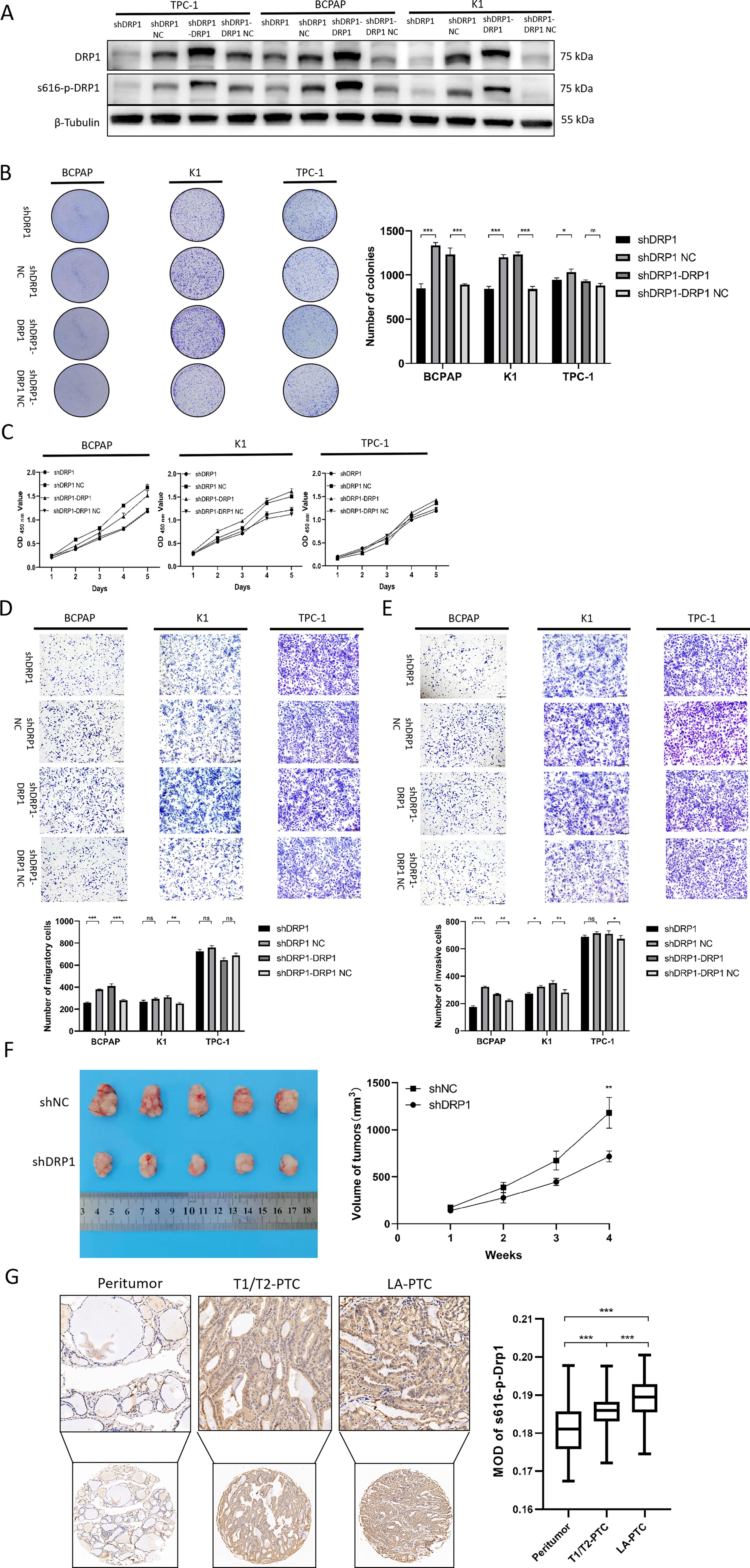
The impact of DRP1 on the malignant behavior of PTC.
BRAFV600E/p-ERK/p-DRP1(Ser616) pathway regulates mitochondrial fission and glucose metabolism reprogramming in PTC
Knockdown of DRP1 in BCPAP and K1 cells resulted in an increased average mitochondrial length and a decreased number of mitochondria per cell (Fig. 6A). Furthermore, with DRP1 knockdown, BCPAP, K1, and BRAFV600E-overexpressed TPC-1 exhibited a higher proportion of cells with high mitochondrial membrane potential (Fig. 6B). A reduction in ECAR and LP was observed in BCPAP-shDRP1 (Fig. 6C), K1-shDRP1 (Fig. 6D), and BRAFV600E-overexpressed TPC-1 with DRP1 knockdown (Fig. 6E). This implies that the BRAFV600E/p-ERK/p-DRP1(Ser616) pathway promotes glycolysis. Conversely, OCR was upregulated in BCPAP-shDRP1 (Fig. 6C), K1-shDRP1 (Fig. 6D), and BRAFV600E-overexpressed TPC-1 with DRP1 knockdown (Fig. 6E), indicating that the pathway inhibits OXPHOS. WB analysis and RT-qPCR showed an upregulation of HK2 with BRAFV600E overexpression in TPC-1 and a reduction following DRP1 knockdown in BCPAP and K1 cells (Fig. 6F).
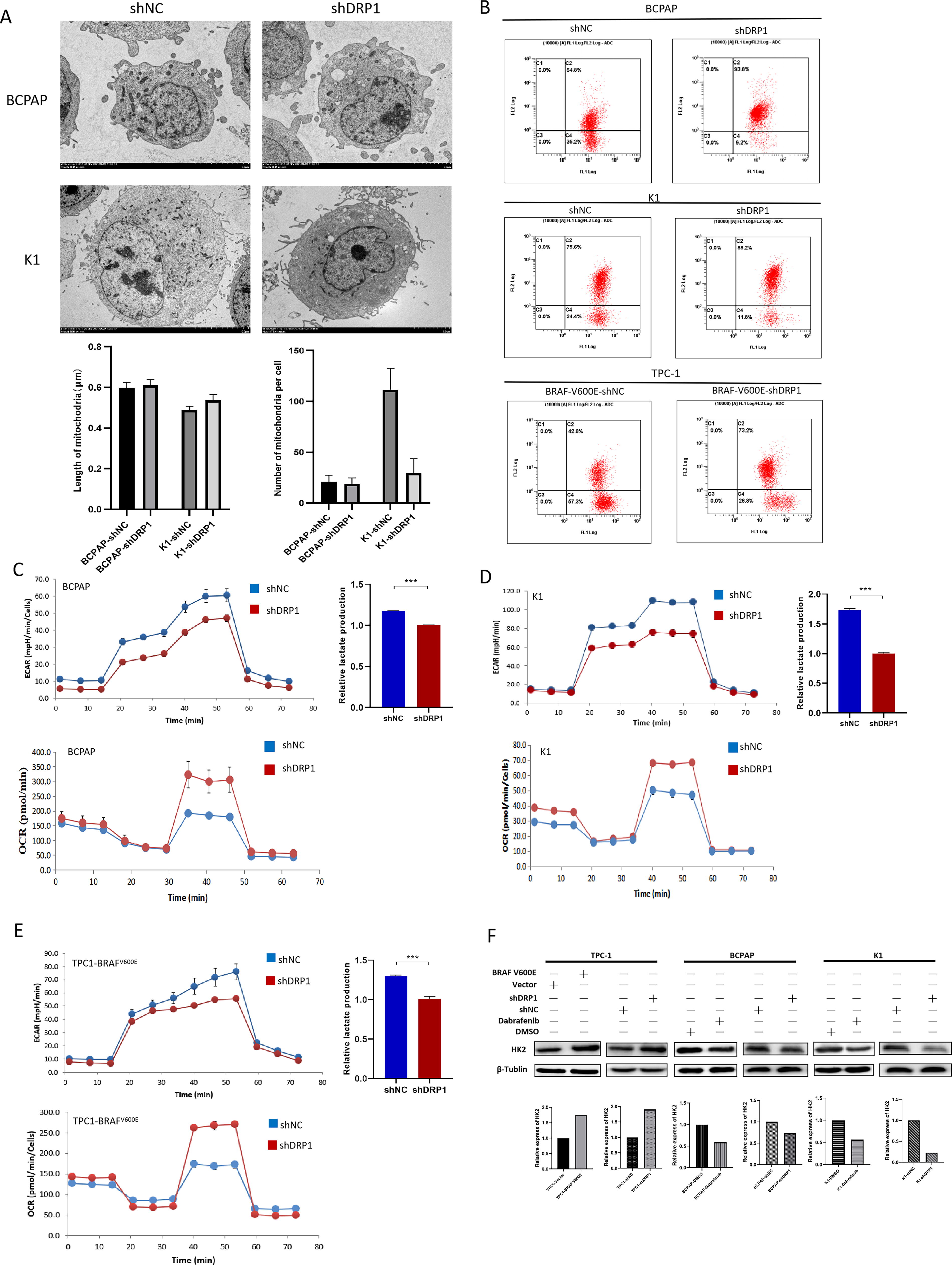
Regulation of mitochondrial fission/fusion and glucose metabolism reprogramming by BRAFV600E/p-ERK/p-DRP1(Ser616) pathway in PTC.
BRAFV600E/p-ERK/p-DRP1(Ser616) signaling pathway regulates ROS and apoptosis in PTC
Knockdown of DRP1 led to an increase of cellular ROS in BRAFV600E-overexpressed TPC-1, BCPAP, and K1 (Fig. 7A). Increased apoptosis occurred following DRP1 knockdown in BRAFV600E-overexpressed TPC-1, BCPAP, and K1 (Fig. 7B, C, and D). These findings indicate that the BRAFV600E/p-ERK/p-DRP1(Ser616) signaling pathway plays a regulatory role in both ROS production and apoptosis in PTC.
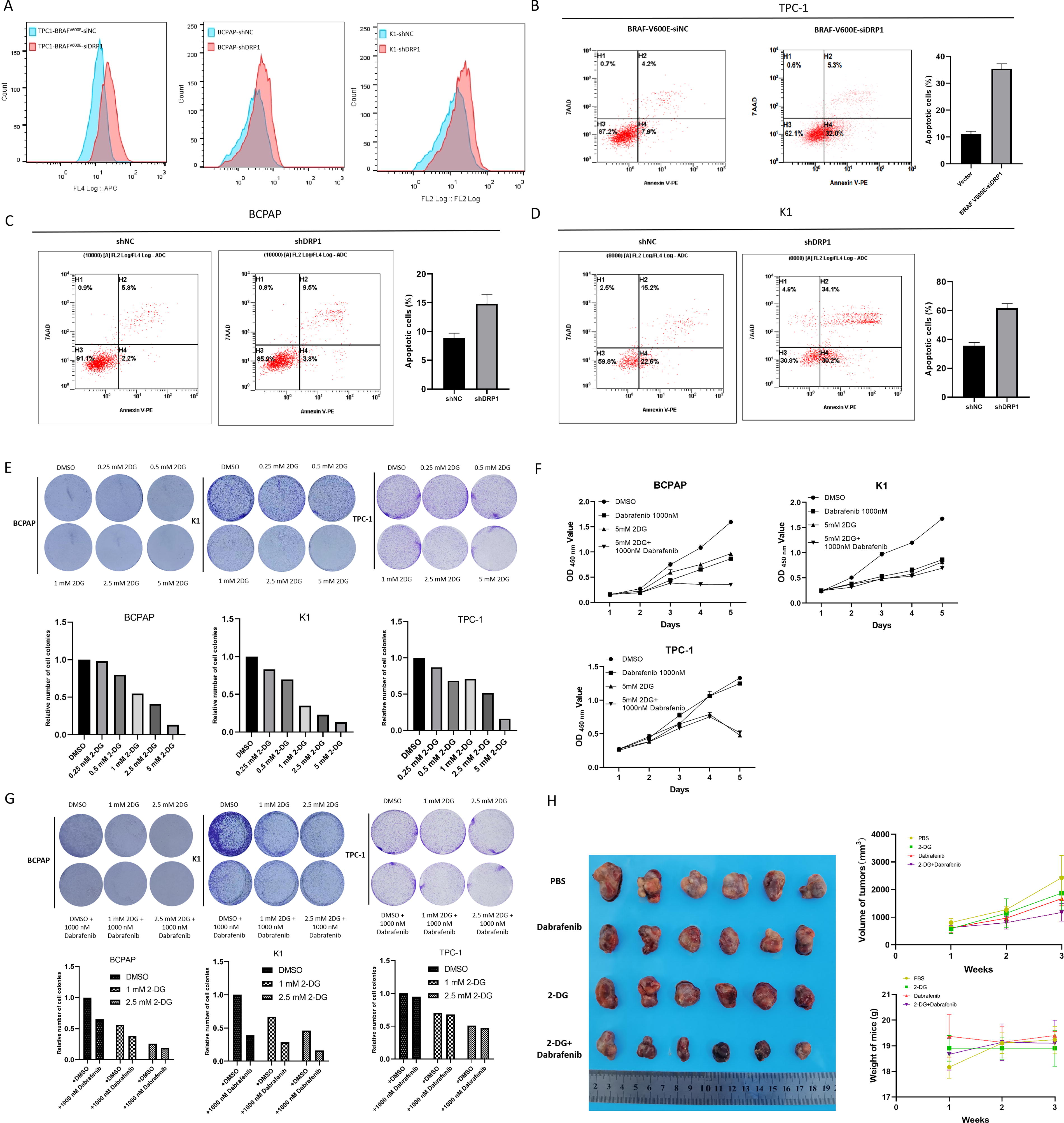
Regulation of ROS production and apoptosis by BRAFV600E/p-ERK/p-DRP1(Ser616) pathway in PTC and drug experiments.
Agent experiments targeting BRAFV600E and glycolysis
In vitro experiments demonstrated the efficacy of 2-DG in inhibiting cell proliferation in BCPAP, K1, and TPC-1 cells (Fig. 7E). The inhibitory effect of 2-DG on TPC-1 is weaker than that of BCPAP and K1 when the concentration is lower than 2.5 mM. Notably, the combination of 2-DG and dabrafenib exhibited a more pronounced inhibitory effect on BCPAP and K1 compared with either drug used alone (Fig. 7F, G). In vivo experiments showed that the combination of 2-DG and dabrafenib more effectively suppressed tumor growth than either agent individually, and no significant differences in mouse body weight were observed among the different treatment groups (Fig. 7H).
Discussion
Despite notable advancements in treating BRAFV600E TC following the introduction of BRAFi, the prognosis for PTC with the BRAFV600E mutation is limited by drug resistance. 27 A previous study found that targeting ACYP1, which promotes glycolysis in hepatocellular carcinoma, can significantly reduce lenvatinib resistance and inhibit tumor progression. 28 Thus, drugs targeting glucose metabolism may be the key to overcoming resistance to tyrosine kinase inhibitors. Our study not only confirms that BRAFV600E supports the Warburg effect in PTC through the BRAFV600E/p-ERK/p-DRP1(Ser616) signaling pathway but also demonstrates its detailed role in regulating glucose metabolism and tumor growth. Furthermore, we propose a promising therapeutic strategy for BRAFV600E-mutated PTC.
GSEA of the TCGA database revealed a significant downregulation of the mitochondrial OXPHOS electron transport chain pathway in patients with BRAFV600E mutations. RT-qPCR analysis of 80 pairs of tissues showed increased expression of glycolysis-related enzymes in PTC, especially in samples with the BRAFV600E compared with BRAFWT. TEM image analysis and flow cytometric analysis indicated that BRAFV600E promotes mitochondrial fission in PTC. Further investigations demonstrated an elevation in glycolysis and simultaneous inhibition of OXPHOS in PTC cells due to BRAFV600E. Additionally, BRAFV600E was found to reduce cellular ROS production and apoptosis in PTC cells.
DRP1 is a fission-mediating GTPase that localizes to mitochondria and initiates membrane scission upon phosphorylation at serine 616. 29,30 After discovering the effect of BRAFV600E on mitochondrial morphology in PTC cells, we attempted to explore the regulation of BRAFV600E on DRP1, which plays a crucial role in mitochondrial fission. WB analysis revealed a BRAFV600E/p-ERK/p-DRP1(ser616) regulatory axis in PTC. This pathway also exists in HEK-293 cells, indicating its widespread presence. IHC results demonstrated that p-DRP1(Ser616) expression is elevated in PTC compared with peritumor tissue and is higher in BRAFV600E-positive PTC than in PTC without BRAF mutation. Furthermore, its expression was higher in LA-PTC than in early-stage T1/T2 PTC, suggesting a possible correlation between p-DRP1(Ser616) expression and poor PTC prognosis. Cellular experiments revealed that the BRAFV600E/p-ERK/p-DRP1(Ser616) pathway enhances mitochondrial fission, glycolysis, and the malignant behavior of PTC cells. Animal models demonstrated that DRP1 knockdown inhibited PTC growth. No significant changes in glucose metabolism and malignant behavior were observed in TPC-1 after knocking down DRP1, suggesting that DRP1 depends on BRAFV600E to exert its function in PTC.
HK2, the initial rate-limiting enzyme in the glycolytic pathway, was found to be influenced by the BRAFV600E/p-DRP1(Ser616) axis. This aligns with the previous study, which reported that DRP1 in Ras-driven pancreatic tumors enhances glycolysis through HK2 regulation. 31 Aerobic glycolysis not only boosts ATP production in energy-deficient environments but also supplies intermediates for rapid cell proliferation and acidifies the tumor microenvironment. It has been observed that cells with higher glycolytic capability exhibit more aggressive malignant behaviors in tumors. 32,33 Therefore, our findings corroborate that the BRAFV600E/p-ERK/p-DRP1(Ser616) axis enhances glycolysis to promote malignant behaviors in PTC cells with BRAF V600E by regulating HK2.
Moreover, mitochondria, the site of OXPHOS, rely on a stable mitochondrial inner membrane for efficient electron chain transfer. Thus, mitochondrial fragmentation and excessive fission can impair OXPHOS, which is a primary source of intracellular ROS. 34 ROS acts as a double-edged sword in tumorigenesis and progression. While it activates protumorigenic signals and enhances cell survival, it also triggers oxidative stress-induced tumor cell apoptosis. 35 Our study suggests that the BRAFV600E/p-ERK/p-DRP1(Ser616) axis promotes mitochondrial fission, thereby weakening OXPHOS, reducing ROS production, and ultimately decreasing cell apoptosis in BRAFV600E-positive PTC.
The first step of glycolysis is that glucose is phosphorylated by HK to form glucose 6-phosphate (G-6-P). 2-DG, a mimic of glucose, interacts with HK to generate 2-deoxy-
While this study provides valuable insights into the treatment of BRAFV600E-positive PTC, it is not without its limitations. One of the critical gaps in our understanding is the detailed molecular mechanism underlying the increased expression of HK2 in PTC. Future studies should focus on unraveling these molecular pathways. The combination of 2-DG and dabrafenib shows promise in treating BRAFV600E PTC. However, more comprehensive trials are needed to detect the efficacy and safety of this drug combination. Such trials will be crucial in translating these findings into clinical practice.
Conclusion
In summary, our findings confirm that the BRAFV600E mutation promotes glucose metabolism reprogramming and mitochondrial fission in PTC through the BRAFV600E/p-ERK/p-DRP1(Ser616) pathway. The dual inhibition of this pathway using 2-DG and dabrafenib presents a promising therapeutic approach for patients with BRAFV600E PTC.
Footnotes
Authors’ Contributions
N.Q., B.M., and Q.J. proposed the concept and conducted the work. S.W. and Y.W. drafted and revised the review. J.W. and Y.W. assisted with data analysis and interpretation. S.D. and X.X. provided technical support. Y.W. collected samples. Y.W. and N.H. assisted in collecting follow-up information. S.W., JW., and Z.N. conducted basic experiments. All authors gave final approval of the version to be published.
Author Disclosure Statement
The authors declare no financial interests related to this study.
Funding Information
This study was supported by grants from the
Supplementary Material
Supplementary Data S1
Supplementary Figure S1