
Abstract
Background:
The “neutral” thyrotropin receptor autoantibodies (N-TSHR-Ab) directed at the TSHR ectodomain’s hinge region have been shown to induce thyroid cell damage in vitro. During these earlier studies, we developed a mouse monoclonal antibody (MC1) specific for a peptide (amino acid 322–340) in the region (MC1-Mab) which was able to induce thyroid cell stress and apoptosis when administered in vivo.
Methods:
In order to examine the effect of in vivo generated N-TSHR-Abs, rather than an acutely administered monoclonal antibody, we immunized Balb/c mice with the hinge region peptide over 18 weeks. Serum TSHR antibodies, specific TSHR hinge region antibodies, serum thyroglobulin (TG) and anti-TG as well as thyroxine and thyrotropin (TSH) levels were examined to evaluate the response to the immunization. Histological examination of the thyroid glands and flow cytometry of spleen T cells, B cells and macrophages were also performed to explore the underlying mechanisms.
Results:
We found that TSHR-peptide immunized mice developed N-TSHR-Abs against the peptide which resulted in thyroid damage shown by thyroid follicular destruction with follicular cell apoptosis, M1 macrophage infiltration, thyroglobulin release, and induction of thyroglobulin antibodies. This resulted in hypothyroidism with increased TSH levels.
Conclusion:
This study demonstrated that endogenous neutral antibodies to the TSHR could induce thyroid cell damage from apoptosis and M1 macrophage infiltration and resulted in hypothyroidism.
Introduction
The thyrotropin receptor (TSHR) is the main autoantigen for Graves’ disease (GD) and may also be implicated in Hashimoto’s thyroiditis (HT). 1 Autoantibodies that target the TSHR in patients with GD can be classified as stimulating TSHR Abs, blocking TSHR Abs and neutral (also termed hinge region) TSHR Abs (N-TSHR-Abs) based on their influence on cyclic adenosine monophosphate (cAMP) signaling. 2 Stimulating TSHR Abs induce the activation of the TSHR located on the basolateral surface of thyroid epithelial cells, triggering multiple G protein activation. 3 Blocking TSHR Abs impedes the action of TSH. 3 Antibodies targeting the linear epitopes in the hinge region of the TSHR are commonly referred to as N-TSHR-Abs. They do not stimulate the generation of cAMP. 3 However, previous studies from our laboratory have indicated that these “neutral” antibodies can initiate a series of cellular events that ultimately result in programmed cell death by activating the intrinsic pathway of apoptosis 4 suggesting that they do not possess neutral functionality. The N-TSHR-Abs can be detected in patients with GD, 5,6 suggesting their potential involvement in the disease.
The hinge region of the TSHR comprises 114 amino acids and stretches from amino acid (aa) 280 to 410 and contains a unique 50-aa cleaved region from aa 316 to 366. Based on earlier studies on signaling characterization of neutral antibodies, 7 we developed a mouse monoclonal antibody (MC1) to aa 322–340 (called peptide 21) in the cleaved region of the receptor. 4 Our previous acute in-vivo study demonstrated that 6 injections of MC1 mAb over 12 days led to endoplasmic reticulum stress, mitochondrial stress, and apoptosis in the thyroid glands. 4 These data provided proof of the close involvement of hinge region antibodies in mediating thyroid damage and serving as an antigenic sink for the immune response in the disease. To further validate this concept of active induction of thyroid damage, we immunized mice with this specific peptide 21 from the hinge region of the TSHR and examined its effects over an extended period (18 weeks) to assess its potential to elicit N-TSHR-Abs and study the effects of this chronic immune response on thyroid function.
Material and Methods
Animals and immunization
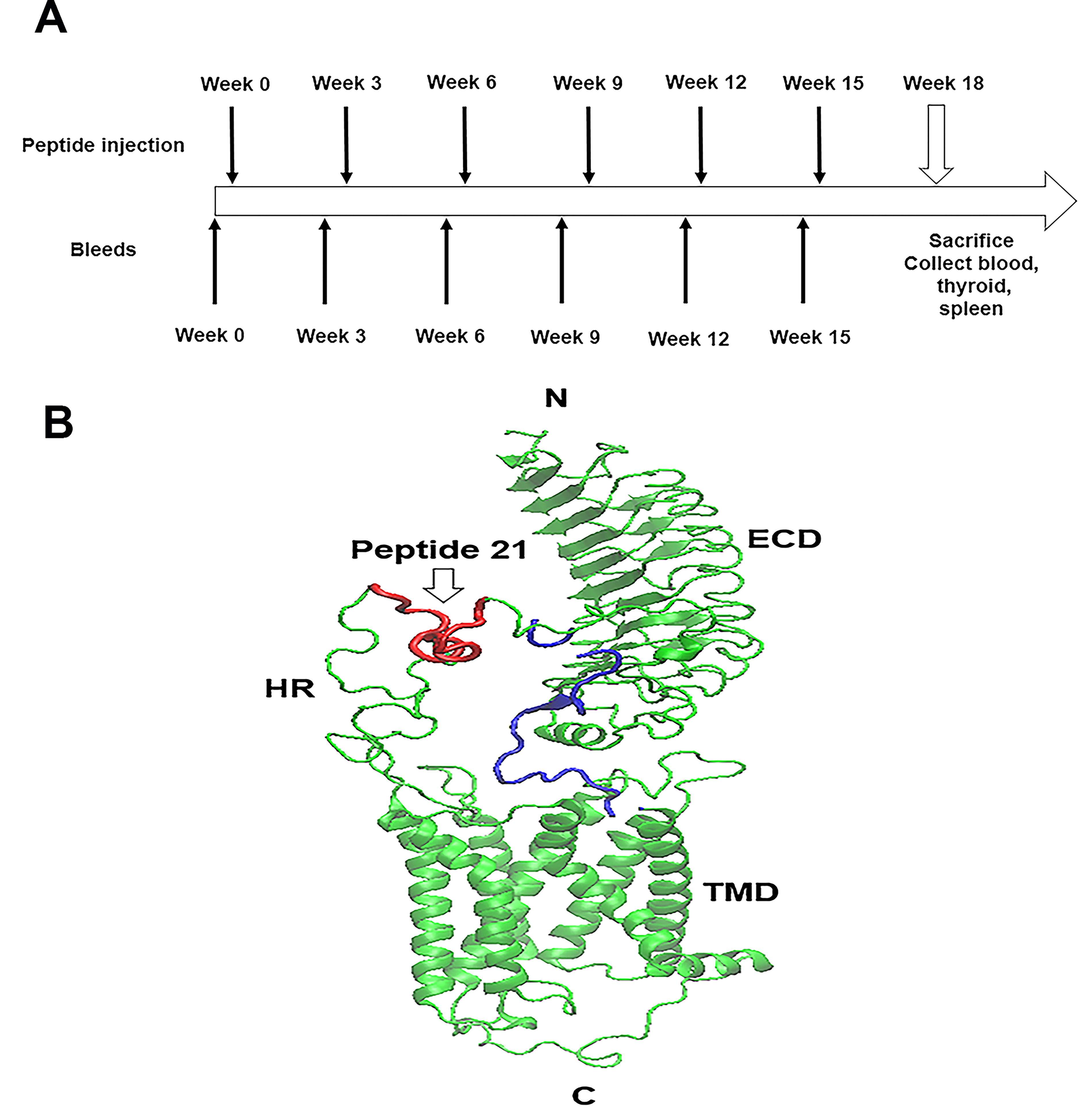
25 SPF Female BALB/c mice aged 6–8 weeks were purchased from Jackson Labs. Ethical approval for all animal studies was obtained from the Institutional Animal Care and Use Committee (IACUC LA11-00267). Keyhole Limpet Hemacyanin (KLH)-conjugated hinge region TSHR peptide (aa 322–340) and scrambled hinge region TSHR peptide (aa 322–340) were synthesized by Biomatik USA LLC. The peptides were dissolved and then diluted to a concentration of 1 mg/mL in calcium and magnesium-free 1X PBS before immunization. The animals were divided into 2 groups: (i) immunized group, 20 mice were given KLH-conjugated hinge region TSHR peptide (peptide 21) via intraperitoneal injection and (ii) control group, 5 mice were injected with KLH-conjugated scrambled hinge region TSHR peptide. The initial injection involved a dosage of 100 μg of peptide along with 100 μL of Incomplete Freund’s adjuvant (IFA) (YA357364, Thermo Scientific, USA), followed by a subsequent dose of 50 μg of peptide and 50 μL of IFA. All animals received 6 injections at 3-week intervals. Blood was collected by submandibular bleeding at weeks 0, 3, 6, 9, 12, 15, and 18 (Fig. 1). Serum samples were separated and stored at −80°C.
Thyroid function and thyroid antibody testing
The concentration of thyroxine (T4) and thyrotropin (TSH) levels were measured by the Millipore MAP thyroid magnetic bead method (Cat # RTHYMAG-30K) following the manufacturer’s protocol. TGAb (L230602857, Biomatik, USA) and thyroglobulin (Tg) (L230525316, Biomatik, USA) concentrations were determined using Enzyme-linked immunosorbent assay (ELISA) as per the manufacturer’s protocol.
TSHR antibodies detected by flow cytometry
Unfixed CHO-TSHR cells were used to detect TSHR antibodies and were cultured overnight at 37°C in F12 medium with 10% Fetal Bovine Serum (FBS) and 100 units penicillin and streptomycin. The cells were detached by 1 mM EGTA/EDTA and washed twice with 1X PBS. Then filtered through 75 μM filter and resuspended in 100 μL of FACS staining buffer (1X PBS with 0.2% sodium azide and 2% FBS) at a density of 0.5 × 106 cells/tube. The cells were incubated with test and control serum (diluted 1:50) for one hour at room temperature. TSHR receptor binding antibodies were detected using phycoerythrin (PE) labeled secondary anti-mouse antibody Fab’. Negative and positive controls were included, using a tube with no primary antibody and anti TSHR mAb MC1 (10 μg/mL), respectively. The results were expressed as the Median Fluorescence Intensity (MFI) of positive cells detected in the test samples compared to the control serum samples. All time period sera were tested.
Detection of TSHR hinge region antibodies
We conducted a series of qualitative ELISAs using nine overlapping 20-mer peptides, which covered the entire human TSHR hinge region (amino acids 280–410) as illustrated in Fig. 1B. 8 Wells were coated with 100 μL of peptides dissolved in bicarbonate buffer pH of 9.6, at a concentration of 10 μg/mL. Subsequently, the plates were blocked with 2% BSA for 1 hour at 25°C. Mice serum samples were diluted 1:100 and incubated for 1 hour at 25°C. Detection was performed using anti-mouse IgG-Fab’ horseradish peroxidase (HRP) as the detection antibody. Optical density measurements were taken at 450 nm using a CalrioStar microplate reader (BMG Inc). The ELISA method exhibited high specificity when tested with control peptides. The cut-off value was determined as ±2 standard deviations from the average of the control serum samples for each of the peptide plates.
Histology
At the end of 18 weeks, half of the thyroid glands and spleens of the control and immunized mice were harvested and preserved and fixed in 10% buffered formalin in 0.9% NaCl. Following fixation, the thyroid gland was embedded in paraffin and subjected to hematoxylin and eosin (H&E) staining to examine gross histological changes. To assess apoptotic cell death, we quantified apoptosis in each tissue slide using a TUNEL Assay Kit (4812–30-K, Trevigen, USA) according to the manufacturer’s instructions. Images of the stained H&E and TUNEL slides were captured using a Hamamatzu NanoZoomer Digital slide scanner at 20x magnification. Images obtained were used for quantification through ImageJ.
Immunofluorescence staining of thyroid gland
At the end of 18 weeks, one half of thyroid glands were embedded in optimal cutting temperature compound and sliced into frozen section (5 µm). The sections were blocked with 10% normal goat serum, and 0.3% Triton X-100 in PBS for 2 hours at room temperature, followed by incubation of primary antibodies: F4/80 rabbit antibody (CST, 30325T), mouse anti-iNOS antibody (Biolegend, 690902), and mouse anti-arginase 1 antibody (Biolegend, 678801) (1:200). Anti-mouse CD86 antibody (Biolegend, 105001) and anti-mouse CD163 antibody (Biolegend, S15049I) (1:100) at 4°C overnight. After washing, the slides were incubated with Alexa Fluor 488 anti-Rabbit IgG (CST, 4412) and Alexa Fluor 594 anti-mouse IgG (CST, 8890) (1:500) for 2 hours at room temperature and washed three times in PBS with Tween 20, mounted with mounting medium containing nuclear dye (4¢,6-diamidino-2-phenylindole), sealed with nail polish and visualized immediately under an ECLIPSE Ti2 inverted microscope (Nikon, Tokyo, Japan) equipped with NIS-Elements software (version 5.30.02, Nikon, Amsterdam, Netherlands). Images obtained were used for quantification through ImageJ.
Statistics
Statistical data were expressed as mean ± standard error of the mean. Two-tailed t-tests were performed. Differences were considered statistically significant at a p value <0.05. GraphPad Prism, version 8.0 (GraphPad Software, San Diego, CA) and SPSS software, version 26.0 (IBM SPSS Statistics, Armonk, NY) was used for all statistical analyses.
Results
The hinge region TSHR peptide generates neutral TSHR antibodies
Twenty Balb/c mice received injections of hinge region peptide, and 5 mice received scrambled peptide every 3 weeks over an 18-week period, with sequential blood collections as shown in Fig. 1A. We detected antibodies against human TSHR, stably expressed on CHO cells, using flow cytometry. The immunized group had detectable TSHR antibodies as early as 3 weeks after immunization. Fig. 2A shows a representative histogram of FACS data for mice. The median MFI and percentage of PE-labeled TSHR expressing cells with sera from the immunized group significantly exceeding those in the control group. With time, more mice developed these TSHR antibodies and showed an increase in the serum levels (Fig. 2B) although the binding of TSHR receptor antibodies to native TSH receptor on CHO cells exhibited significant variability within the immunized group. We also performed indirect peptide ELISA’s with 9 20-mer overlapping peptides covering the entire TSHR hinge region. Positivity was determined using the cut-off criteria of ±2 SD above mean of the control group. As time progressed, more and more mice showed detectable antibody responses to the immunized peptide and this response was sustained until 18 weeks (Fig. 2C). The titers of the antibody ranged up to 1:1600 (see Supplementary Fig. S1). Note also that none of the immunized mice showed significant responses to the other 8 hinge region peptides thus excluding epitope spreading (data not shown). To examine if the immune response included stimulating or blocking TSHR antibodies, we used the luciferase reporter “TSHR-Glo” bioassay 9 as described in Supplementary Data S1. The results showed that none of the immunized animals developed TSHR stimulating or blocking antibodies (Supplementary Fig. S2). Overall, the hinge region TSHR peptide generated only epitope specific neutral antibodies to the TSHR.
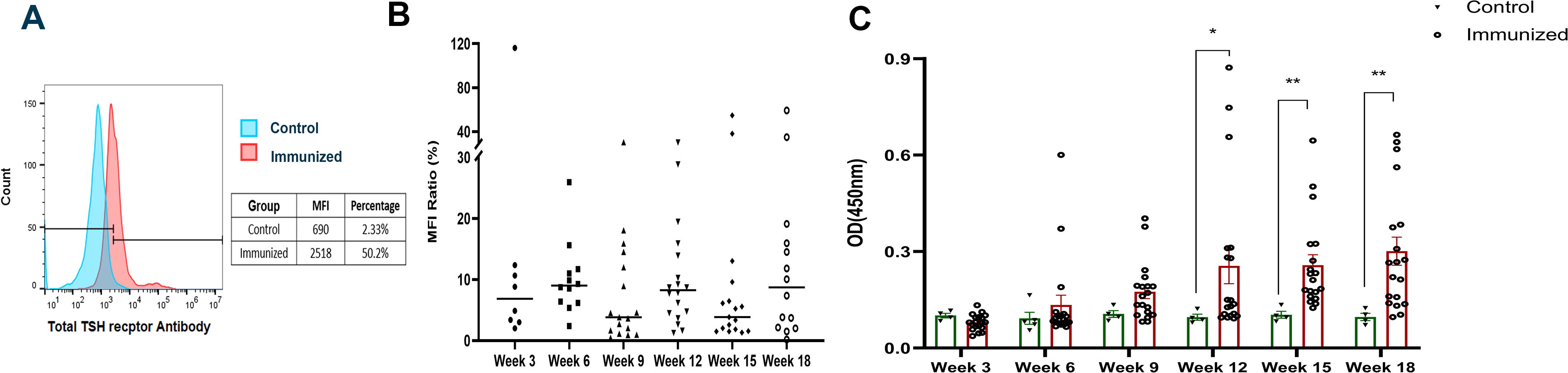
The hinge region TSHR peptide generates neutral TSHR antibodies in mice:
Endogenous neutral TSHR antibodies decrease T4 and elevate TSH in balb/c mice
To examine the effects of these endogenous N-TSHR-Abs on thyroid function, we initially found that serum T4 levels were lower in TSHR immunized mice compared to the control group, while serum TSH levels were higher than those in the control group (Fig. 3A and B). These data indicated that N-TSHR antibodies induced a state of hypothyroidism. To exclude TSH blockade by the immunizing peptide itself, we tested TSH stimulation in the presence of the peptide using the TSHR-Glo bioassay, which showed no evidence of blockade and thus confirmed that it was thyroid damage inhibiting thyroid function (Supplementary Fig. S3). Hence the increase in the TSH levels in the immunized mice could not be caused by the reactive antibodies interfering with TSH action but were evidence of directly induced thyroid hypofunction.
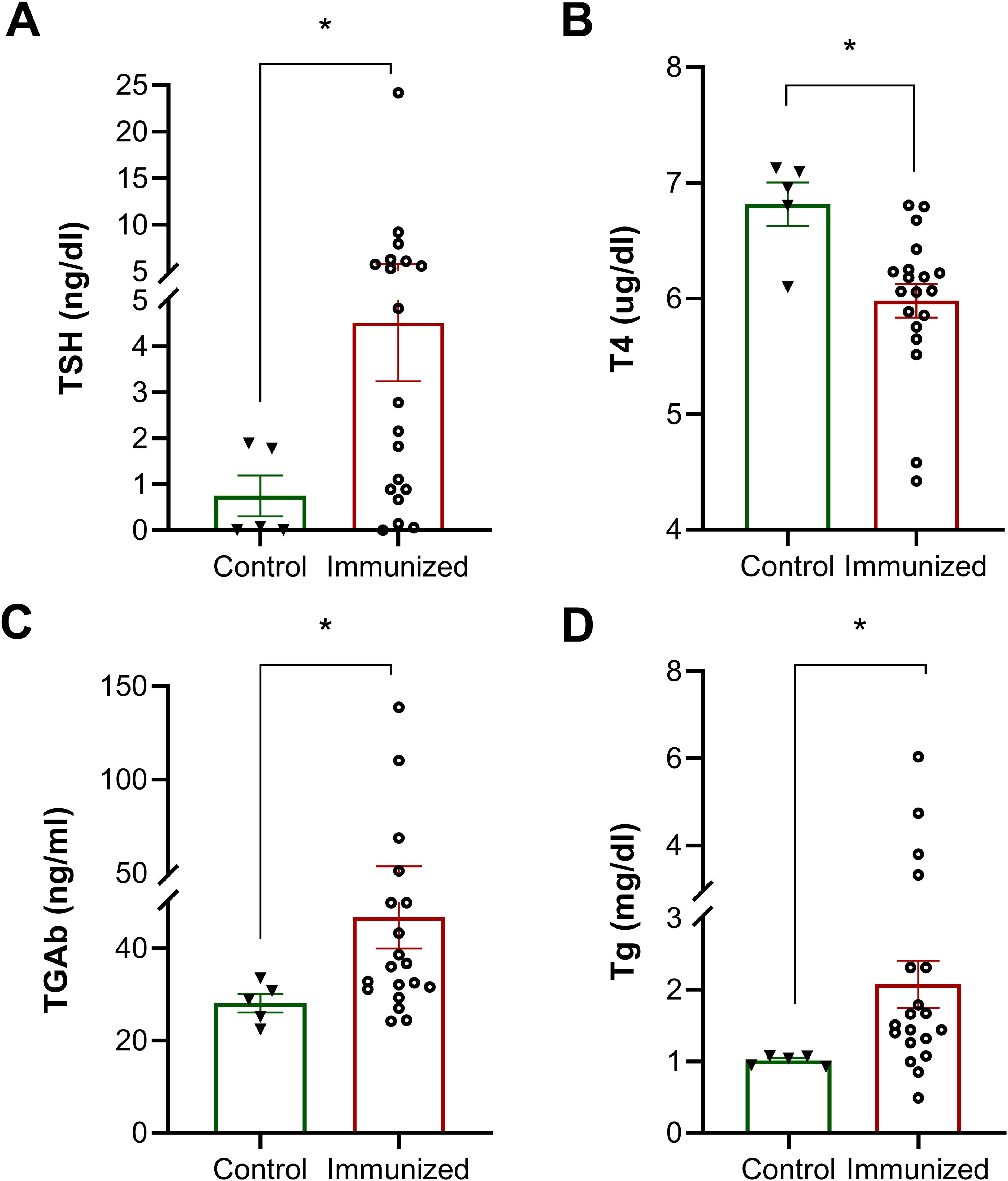
Effect of neutral TSHR antibodies on the thyroid
Endogenous Tg release and induced TgAb
Serum Tg was elevated in the peptide immunized mice consistent with thyroid damage. In addition, Tg-Ab appeared in the serum of the immunized mice secondary to thyroid cell death (Fig. 3C and D). The chronic presence of the N-TSHR antibodies in response to the immunization appeared to markedly compromise thyroid cell function.
Neutral TSHR antibodies directly induce thyrocyte apoptosis
In this 18-week model of induced neutral TSHR antibodies, we found destroyed follicles, irregularly shaped follicles empty of colloid, and some follicles with incomplete boundaries (Fig. 4A). In the control group, the sections showed variable sized but mostly large well-formed follicles with a relatively round shape, filled with homogeneous colloid. Quantitative image analyses confirmed the significant presence of destroyed follicles in the immunized mice (Fig. 4C). TUNEL assay analyses to investigate the degree of apoptosis in the thyroids revealed positive apoptotic cells only in thyroids of the peptide 21 immunized mice (Fig. 4B). There were many follicles which exhibited apoptotic cells, and there was a non-uniform distribution of these apoptotic cells throughout the section. Quantitative analysis of apoptosis showed that the tunnel positive cells were significantly higher in TSHR peptide-treated mice compared to controls, where virtually no apoptotic staining was observed (Fig. 4D).
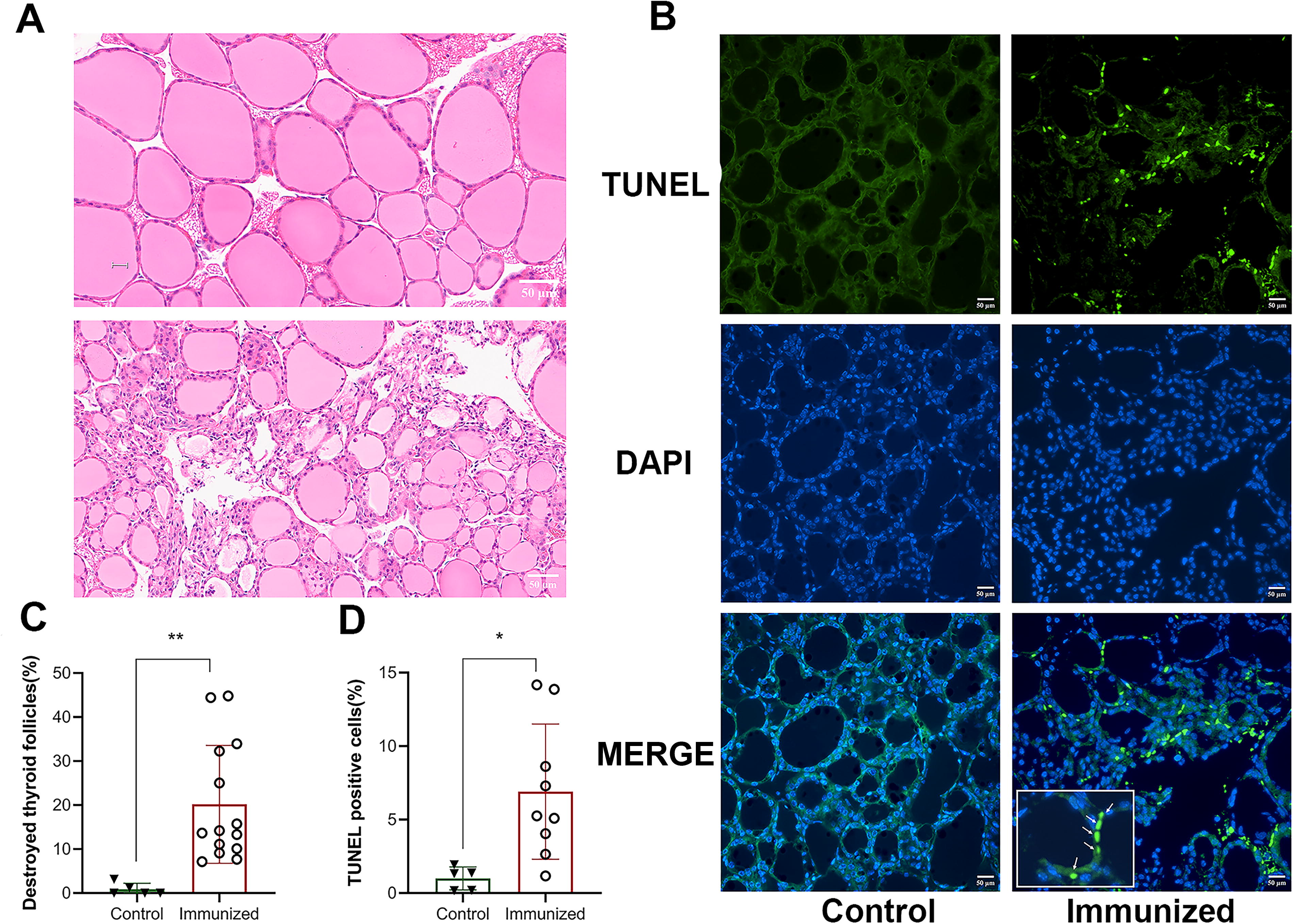
Neutral TSHR antibodies induce thyroid damage and thyrocyte apoptosis in mouse thyroid glands. TSHR peptide immunized mice show higher proportion of destroyed follicles (p < 0.05) compared to control mice, as shown in
Neutral TSHR antibodies induce M1 macrophages infiltration
Macrophages have an important role in antigen processing and in the clearance of dying and dead cells. They also play a crucial role in the onset and progression of autoimmune diseases. We detected macrophages by immunofluorescence staining and demonstrated that compared to control peptide treated mice, macrophages were detected in TSHR peptide immunized mice, especially, there was an infiltration of M1 macrophages in the thyroid gland (p < 0.01) (Fig. 5A, B, E, and F), whereas M2 macrophages did not appear to be involved (Fig. 5C, D, G, and H).
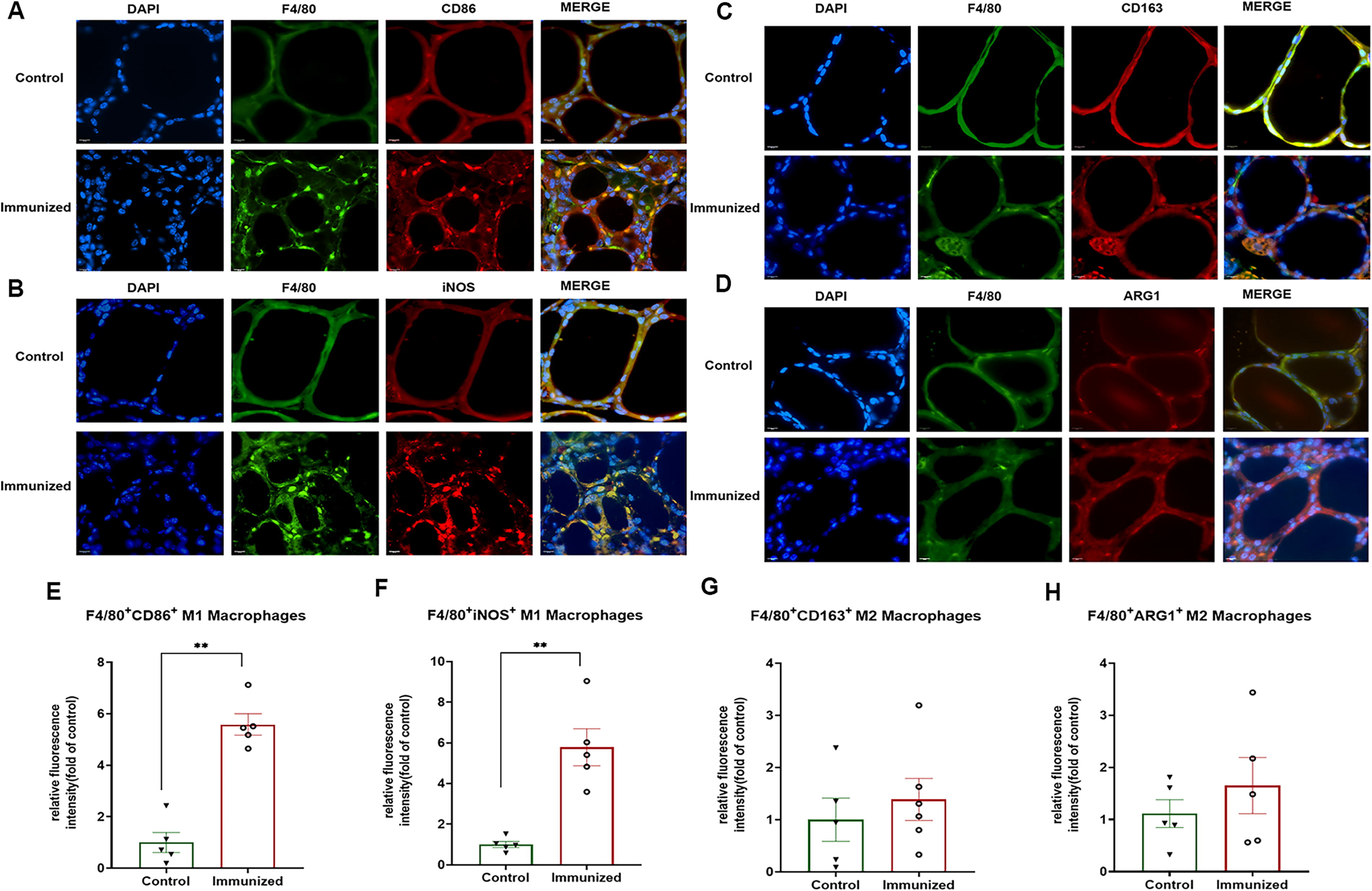
Neutral TSHR antibodies induce M1 macrophages infiltration in mouse thyroid glands. Compared to control peptide treated mice, the M1 macrophages (green stained cells, inset F4/80 with red stained cells, inset CD86 and iNOS) in TSHR peptide immunized mice were found to be elevated (p < 0.01), as shown in
CD4+ T cell, treg, macrophage, and CD8+ T cell profiles
CD4+T cells assist effector B cells in augmenting autoimmunity. Therefore, we measured the profile of B and T cells in the spleens of immunized and control mice. The data showed that overall there were no significant differences between the average percentage of CD45+CD3+T cells and CD45+CD19+B cells in the spleen in the immunized versus the control group (p > 0.05) (Supplementary Fig. S4A and S4B). On further subtyping these T and B cells into memory B cells, CD4+T cells and CD8+T cells and Treg cell, we found no difference among memory B cells (Supplementary Fig S4G and 4H), CD4+T cells and CD8+T cells (Supplementary Fig. S4C and S4D) and Treg cells (Supplementary Fig. S4E and S4F) in the 2 groups. Additionally, there was no difference between the percentage of spleen macrophages in the immunized group and the control group (Supplementary Fig. S4I and S4J).
Discussion
The TSHR serves as the principal regulator of thyrocyte function and growth 10 and it is a major autoantigen in autoimmune thyroid disease. 1,3 The TSHR is composed of a large ectodomain (ECD) and membrane-bound signal transducing transmembrane domain (TMD) 8,11 (Fig. 1B). The ECD can be subdivided into a leucine-rich domain, which takes on a curved structure, and is connected to the TMD through a 130-aa hinge (or linker region (aa 80-410). 12 Auto antibodies that are specific for the TSHR may be “stimulating,” “blocking,” or “neutral” in their activity at the TSHR. 2 Neutral TSHR Abs do not trigger a conventional signal, nor do they inhibit TSH binding. 13 These antibodies primarily, although not exclusively, bind to linear epitopes within the hinge region. 14 The variable signaling profile of these antibodies has now been well characterized by earlier studies. 3,5,15 The hinge region, which also acts as a scaffold, is an intrinsically disordered protein as recently shown by our modeling studies. 12 Our previous data also showed there is a substantial affinity between the hinge region and TSH and autoantibodies, 16 indicating that the hinge region plays an important part in activation of the receptor. 12 Furthermore, our studies with full-length TSHR immunization of mice have shown that the hinge region of the TSHR has many epitopes that are antigenic “hot spots” for neutral autoantibodies. 14
In this study, we have examined the effect of the long-term presence of neutral TSHR antibodies to a peptide corresponding to aa 322–340 of the TSHR hinge region (Figs. 1 and 2). We showed that the peptide immunization confined the immune response exclusively to the peptide and showed no epitope spreading. In addition, none of the immunized serum Abs showed any TSHR stimulating or blocking activity. Although it has been reported that thyroid antibodies may display epitope spreading, 17 this phenomenon may be restricted by the molecular size of the protein. 18 However, the peptide immunization did generate robust epitope-specific neutral antibodies allowing us to study the effects of chronic neutral TSHR antibody exposure on the thyroid.
Our previous studies of N-TSHR Abs have shown that they are not truly neutral. 4,19 Instead, they have the capacity to induce thyroid cell stress and trigger a series of events that ultimately result in programmed cell death by activating the intrinsic pathway of apoptosis. 3,5 The short-term injection of neutral TSHR mAb (MC1) also induced thyroid damage with decreased serum T4. 4 In the present studies, we found that endogenous neutral antibodies decreased serum T4 and elevated TSH levels indicating significant hypothyroidism in the immunized mice (Fig. 3). In agreement with the disturbed thyroid function, we found an increased serum thyroglobulin (Tg) and the appearance of thyroglobulin antibodies (Tg-Ab) in many of the mice. Thyroid follicular damage and inflammation of the thyroid gland is well known to induce release of Tg 20 as is the appearance of Tg-Ab following thyroid tissue damage. 21 Our results, therefore, were indicative of thyroid follicle destruction triggered by the immune response. In addition to damaging thyroid cells, we have previously shown that neutral TSHR antibodies are capable of releasing the TSHR splice variant V1.3 from cells. 22 In fact, it is not uncommon that antibodies can be generated to various tissue antigens which are capable of inducing site-specific damage such as seen with brain reactive antibodies, 23 antibodies to DNA in SLE, 24 and antibodies to motor neurons in myasthenia gravis. 25 The destructive mode of action can be complement mediated or non-complement mediated mechanisms such as stress induction as seen with N-TSHR antibodies. 5
Examination of the thyroid glands after 18 weeks of exposure to N-TSHR-Abs confirmed our serum analyses. We found many damaged thyroid follicles and multiple apoptotic cells as clear indications of the destructive nature of the immune response (Fig. 4). The mechanisms responsible for triggering such thyrocyte death have been shown to include the induction of thyroid cell stress and the collapse of the cells protective mechanisms including autophagy. 4 It is fair to speculate that similar mechanisms were in effect in the present study and the paucity of inflammatory cells within the thyroid suggested that these effects were indeed antibody mediated. This was substantiated by the normal T cell analyses. We did not observe a significant change in total T cells, B cells, and memory B cells between the immunized and control groups. For example, Treg lymphocytes play an important role in the development of autoimmune thyroid disease (AITD). 26 The initial assumption was that autoimmune diseases would exhibit a reduced presence of Treg lymphocytes, which play a role in suppressing the immune response 27 although, some studies have documented an elevated presence of Treg cells in patients with AITD. 28,29 However, our study showed that Treg cells were not increased in the spleen of the peptide immunized mice but their functional status was not determined. It is known that TSHRs are expressed in immune cells and it may be that neutral antibodies by increasing the oxidative state of these cells may have impaired their functional state as seen in negatively influenced Treg function due to dysregulated antioxidant mechanisms in liver disease. 30
Macrophages are known to remove dying and deceased cells. 31 They also play a vital role in the occurrence and development of autoimmune diseases. 32 In this study, macrophages were elevated in the immunized group. Studies have revealed a positive correlation between the extent of tissue damage and macrophage infiltration in rheumatic arthritis and systemic lupus erythematosus. 33,34 A recent study presents the diverse cell populations found in thyroid tissues and peripheral blood mononuclear cells obtained from patients with HT through single-cell RNA-sequencing and the results indicated that inflammation-associated macrophages were located within thyroid tissues in HT and may have significant involvement in thyrocyte damage. 35 In this study, M1 macrophage infiltration was found in the thyroid gland but not M2 macrophage infiltration. M1 macrophages are recruited and activated in the thyroid gland, where they produce pro-inflammatory cytokines and chemokines, which not only cause damage to thyroid cells but also attract additional immune cells to maintain the inflammatory milieu in the thyroid gland. A prior study indicated that in a mouse model of thyroiditis, the number of M1 macrophages was elevated compared to the control group. It was also suggested that reducing M1 macrophage polarization could have a therapeutic impact. 36 The increase in resident M1 macrophages in the thyroid of our immunized mice would suggest an active immune response to the immunization peptide thus driving proinflammatory cytokine release.
While these studies were confined to mice, there have been previous studies which have indicated M1 macrophages also infiltrate the thyroid gland in human AITD. Cai et al. 37 found a significant enrichment of M1 polarization-associated genes in the thyroid tissue and peripheral blood CD14+ monocytes/macrophages of patients with HT. Similarly, Tao et al. 38 demonstrated an increase in CD68+CD14+ indicating an increase in proinflammatory M1 macrophages. All these data indicate the important role of macrophages in causing thyroid cell damage and serve as a biomarker for disease activity in AITD. Such observations may aid in developing more targeted and effective treatments.
In conclusion, injection of a peptide from the hinge region of the TSHR over an 18-week period resulted in the production of N-TSHR Abs. These antibodies induced thyroid damage via apoptosis and M1 macrophage infiltration leading to hypothyroidism. The pathogenic role of N-TSHR-antibodies complicates our simplistic thinking of the role of antibodies and T cells in GD.
Footnotes
Acknowledgment
Authors’ Contributions
T.F.D. and S.M. led the project and designed the research. P.X. and R.L. conducted the experiments. P.X. acquired data, analyzed data, and drafted the article. R.L. and T.F.D. critically revised the article. All authors read and approved the final version of the article before submission.
Author Disclosure Statement
T.F.D. is a member of the Board of Kronus Inc, Starr, ID. The remaining authors have nothing to disclose. There is no conflict of interest, which can be perceived as prejudicing the impartiality of the research reported here.
Funding Information
This work was supported in part by a VA Merit Award to T.F.D.
Supplementary Material
Supplementary Data S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1