
Abstract
Background:
Differentiated thyroid carcinoma (DTC) is occurring three times more frequently in females than in males. However, the underlying biological mechanisms driving this discrepancy remain poorly understood. To investigate the causal role of sex hormones and reproductive factors in the risk of DTC, we implemented a two-sample Mendelian randomization (MR) analysis.
Methods:
We utilized genome-wide association studies (GWAS) summary statistics to explore these associations. GWAS data on DTC were derived from a meta-analysis of six studies including 7705 cases and 963,612 controls of European ancestry. GWAS summary statistics on sex hormones, reproductive factors, and gynecological conditions were retrieved from publicly available sources. We used the inverse-variance weighted (IVW) method to estimate odds ratio (OR), with additional sensitivity analyses and conducted multivariable MR (MVMR) to account for potential confounding by body mass index (BMI) and thyrotropin (TSH).
Results:
We identified a positive association between sex hormone binding globulin (SHBG) and DTC (ORivw = 1.13, p = 0.046). After controlling for TSH and BMI in a MVMR analysis, the strength of this association remained similar but lost statistical significance. Bioavailable testosterone also showed a positive but marginally significant association with DTC after adjustment for BMI in the MVMR (ORivw = 1.13, p = 0.07). Putative causal association was observed with uterine fibroids in females under 50 years old (ORivw = 1.52, p = 0.017). Endometrial cancer was associated with DTC (ORivw = 1.15, p = 9.0 × 10−3); however, a genetic correlation of r2 = 13% suggested potential pleiotropy. No significant associations were observed for other investigated factors.
Conclusions:
Our study does not provide strong evidence for a causal role of reproductive and hormonal factors in DTC risk, despite the observed sex disparity in incidence rates. The associations observed with SHBG, bioavailable testosterone, uterine fibroids, and endometrial cancer indicate potential risk factors, but further investigation is required.
Introduction
Thyroid cancer is the most common endocrine malignancy and the ninth most common cancer in the world. 1 Its incidence has increased in recent decades, across all geographic locations and ethnicities. 2,3 Part of this increase is attributed to the improvement of advanced diagnostic medical imaging tools, such as thyroid ultrasonography, ultimately leading to overdiagnosis; 4 moreover, environmental and lifestyle factors may also play a role. 5,6 Differentiated thyroid carcinomas (DTC), which include papillary (PTC) and follicular types, represent about 90% of all thyroid cancers. Females display a notably higher incidence of DTC, with rates three times higher than in males, 7 leading to the hypothesis that female sex hormones and reproductive factors may contribute to the risk of developing DTC. Laboratory research utilizing thyroid cancer cell lines has demonstrated that estrogen stimulation enhances thyroid cancer proliferation through multiple mechanisms. 8 Epidemiological studies have reported associations between DTC and various reproductive factors, including age at menarche and menopause, age at first pregnancy, artificial menopause, miscarriage, and abortions. 9,10 Furthermore, postmenopausal females with gynecological disorders are at higher risk of developing thyroid cancer after hysterectomy. 11 Nonetheless, epidemiological studies on the association of sex hormones and reproductive factors with DTC in females remain inconclusive.
Most evidence on potential risk factors of DTC is derived from observational epidemiological studies, which are susceptible to biases such as confounding and reverse causality. 12 Sex differences in incidence rates could be attributed to detection bias, owing to increased medical surveillance during pregnancy and menstruation periods in females. Mendelian randomization (MR) 13 may address these limitations by using genetic variants associated with risk factors as instrumental variables (IVs) to estimate the causal association between these risk factors and DTC.
We used a two-sample MR approach to investigate whether sex hormones levels, reproductive traits and gynecological disorders, whether occurring either before or as a complication from hysterectomy, are causally associated with DTC, using the largest-scale meta-analysis of genome-wide association studies (GWAS), consisting of a total of 7,705 cases and 963,612 controls of European descent.
Methods
Two sample MR analysis uses genetic variants, typically single-nucleotide polymorphisms (SNPs), from GWAS summary statistics to estimate the causal association of risk factors with outcomes. 13 For SNPs to be valid IVs, three assumptions must be verified: (1) SNPs should be strongly associated with the exposure; (2) SNPs should not be directly associated with the outcome other than through the exposure (i.e., no directional horizontal pleiotropy); and (3) SNPs should not be associated with confounders of the exposure-outcome association.
Data sources and genetic instrument variables
We focused on sex hormones, reproductive traits, and gynecological disorders. The corresponding GWAS datasets were acquired from a variety of consortia and studies, predominantly consisting of European populations (Table 1). The summary statistics of DTC GWAS were gathered from several consortia and studies that included participants (male and female) of European ancestry: Epidemiology of thyroid cancer (EPITHYR), 14 European Prospective Investigation into Cancer and Nutrition (EPIC), 15 UK Biobank (UKBB), deCODE genetics, 16 Italian Study, 17 and FinnGen. 18 We performed a random-effect meta-analysis using the GWAMA software. 19 Detailed information on study design, quality control procedures, selection criteria, and statistical methods for exposures and outcome DTC GWAS is provided in Supplementary Data S1.
Description of the Genome-Wide Association Study Data Sets and Instrumental Variables Used in Our Study
Sample size: number of samples used in the genome-wide association study (GWAS); Number of SNPs: the number of single-nucleotide polymorphisms used in the study; Sample sex: males/females; SNPs at 5.0 × 10−8: number of genome-wide significant SNPs; PMID: PubMedID; Population: European population; Clumped SNPs: Clumped single-nucleotide polymorphism, keeping the most significant (lowest p value); R 2: the proportion of variance explained by instrumental variables; F-statistics: the correlation strengths of exposures; DTC: Differentiated thyroid carcinoma; SHBG: sex hormone binding globulin.
IEU GWAS pipeline is used for processing UK Biobank data (https://gwas.mrcieu.ac.uk/), which is the MRC-IEU-based platform that provides summary statistics of publicly available GWAS studies.
The IVs were selected based on the following criteria 20 : (1) association with the exposure at a genome-wide significance threshold of p ≤ 5.0 × 10−8; (2) SNP independence to prevent interference from linkage disequilibrium (LD) analysis, ensuring the correlation r2 value is under 0.001 in a 10,000 kb window through the European 1000 Genomes Project reference panel; (3) exclusion of noninferable palindromic variants with a minor allele frequency (MAF) greater than 0.30; and (4) exclusion of variants with MAF ≤ 0.01. We also excluded variants associated with both the exposure of interest and DTC (p ≤ 5.0 × 10−8) to prevent pleiotropy. The bias due to weak IVs can be avoided by the strength of the association of each SNP with the exposure through the F-statistic ≥10 (Supplementary Data S1). We computed MR power for binary outcomes 21 using a two-sided p-value of 0.05 (Supplementary Table S1).
Statistical analyses
We estimated the overall causal effect (odds-ratio [OR] and confidence intervals [CI]) of each exposure on DTC using the random-effects inverse-variance weighted method (IVW) 22 for the primary analysis; in sensitivity analyses, we used the weighted median (WM), 23 weighted mode-based 24 estimators, MR-Egger regression, 25 MR-Pleiotropy RESidual Sum and Outlier approach (MR-PRESSO), 26 and multivariable MR (MVMR) 27 (Supplementary Data S1). Lastly, we used the Causal Analysis using Summary Effect Estimates (CAUSE) method, which uses Bayesian modeling that accounts for correlated and uncorrelated horizontal pleiotropic effects to avoid false positives. 28 To investigate the possibility of reverse causation, we conducted a reverse MR analysis, particularly focused on variables significantly associated with DTC in our forward MR analysis. The heterogeneity among the genetic variants was quantified using Cochran’s Q-statistic. Multiple testing correction was not applied as the exposures in our analysis are dependent.
Bias due to overlap participants
Overlapped samples between the exposure and outcome GWAS can lead to overfitting bias. The derivation of analytic formulae for the expected bias under the null and type 1 error rate has been previously estimated by Burgess et al. 29 GWAS on circulating levels of sex hormones, age at first birth, and gynecological disorders include the UKBB, so does our DTC GWAS. We estimated that the expected type 1 error rate is approximately 0.05 for all examined exposures, suggesting minimal bias despite this overlap (Supplementary Table S2).
Ethics
Ethics approvals were obtained by the respective GWAS studies.
Results
Table 2 displays the sex distribution of cases and controls, with additional subcategorization based on age and histology of PTC cases among female. The F-statistic and R 2 for each SNP, as well as for the IVs for both exposures and DTC, can be found in Supplementary Tables S3 and S4.
Descriptive Characteristics Table of the Differentiated Thyroid Carcinoma Genome-Wide Association Study on Sex by Studies
Sex hormones and DTC
Genetically predicted sex hormone binding globulin (SHBG) levels were positively associated with DTC using the IVW method (ORivw per one SD-increase = 1.13 [1.00–1.28], p = 0.046) (Fig. 1 and Supplementary Table S5). MR-Egger indicated directional pleiotropy (PEgger-intercept = 0.058) and a significant positive association (OREgger = 1.35 [1.09–1.67], p = 0.008). The association between SHBG and DTC remained consistent in sensitivity analyses when excluding UKBB (ORivw = 1.15 [1.01–1.31], Supplementary Table S6). In females, the association was consistent, but no longer significant (ORivw = 1.13 [0.91–1.38], Supplementary Table S7), possibly due to a smaller sample size. Obesity and thyrotropin (TSH) are associated with both sex steroid hormones across sex 30 –32 and DTC risk. In MVMR, the association between SHGB and DTC increased after adjustment for TSH (ORMVMR = 1.27 [1.00–1.63]) and decreased in analyses adjusted for obesity-related factors (BMI, ORMVMR = 1.04 [0.94–1.16]; WHR, ORMVMR = 1.05 [0.94–1.17]; WHRadjBMI, ORMVMR = 1.05 [0.93–1.20]) (Fig. 2 and Supplementary Table S8). These results suggest that both TSH and BMI confound the association between SHBG and DTC. When adjusting for both TSH and BMI simultaneously, the association was similar in magnitude to the univariate IVW estimate but nonsignificant with a wider CI (ORMVMR = 1.12 [0.95–1.32]). This suggests a possible independent association between SHBG and DTC.

Causal effects from genetically predicted sex steroid hormones, reproductive traits, and gynecological disorders on the differentiated thyroid carcinomas. N, number of SNPs; OR, odds ratio; CI, confidence interval; IVW, inverse-variance weighted. *Outliers removed: rs149934734, rs547025, rs78378222), p-distortion test = 0.0055.
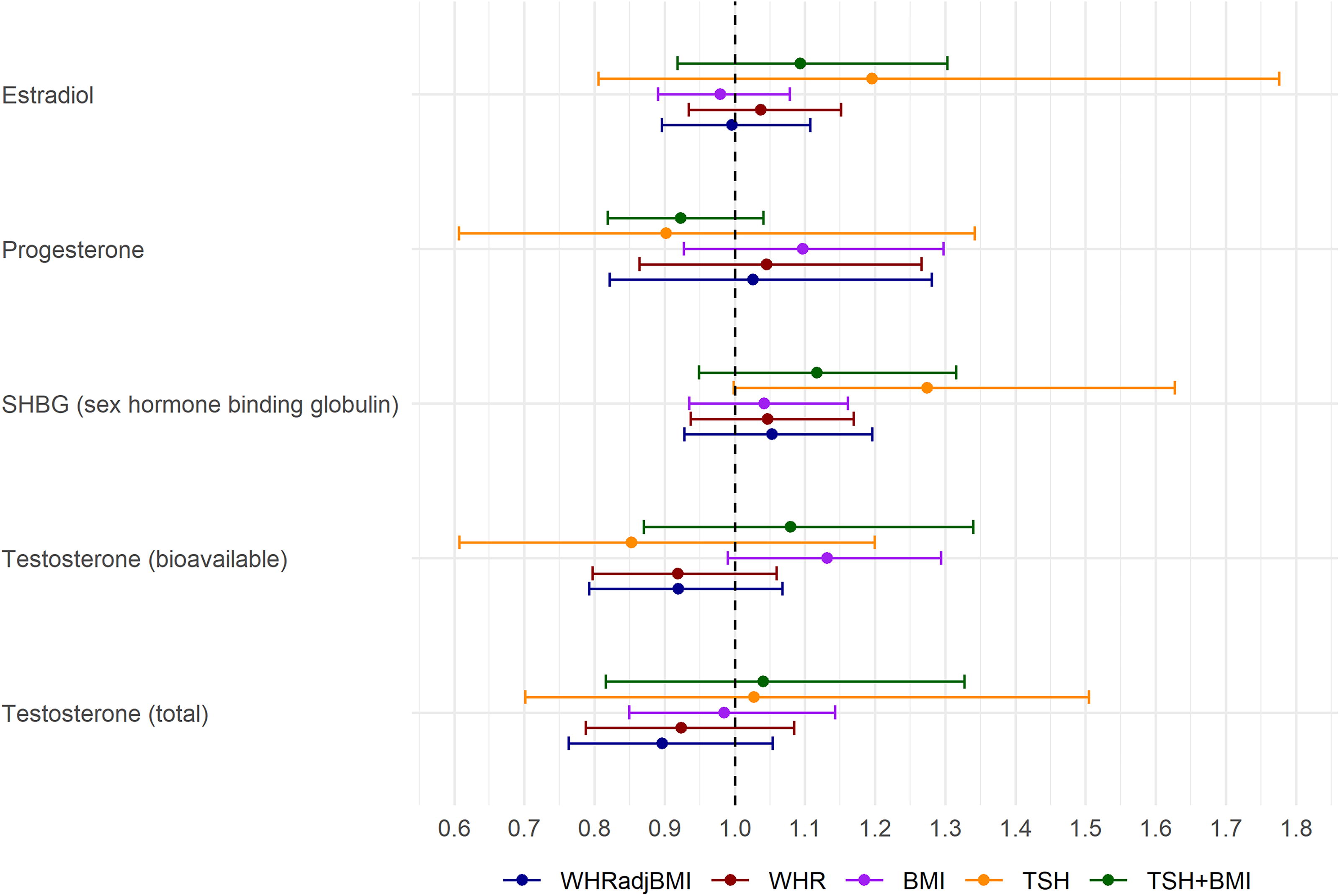
Multivariable Mendelian randomization (MVMR) analyzing the direct effect of sex steroid hormones on differentiated thyroid carcinoma controlling for thyrotropin and obesity-related factors. The MVMR models adjust for TSH (thyrotropin), body mass index (BMI), waist-to-hip ratio (WHR), and waist-to-hip ratio adjusted on body mass index (WHRadjBMI). Bars depict confidence intervals surrounding the direct effect point estimates obtained from MVMR analysis.
A putative inverse association was observed between genetically predicted bioavailable testosterone and DTC risk (ORivw per one SD-increase = 0.88 [0.75–1.03]), with a stronger effect according to the WM method (ORWM = 0.80 [0.62–1.01], p = 0.06). The results were generally consistent after adjusting for TSH, WHR, and WHRadjBMI; however, a putative positive association was found after adjustment for BMI (OR = 1.13 [0.99–1.29], p = 0.07) (Supplementary Table S8).
Reproductive traits and DTC
Genetically predicted age at first birth showed a nonsignificant inverse association with DTC (ORivw per one SD-increase = 0.89 [0.79–1.01], p = 0.078); this finding was supported by other MR sensitivity analyses. Interestingly, after excluding the FinnGen study from the DTC GWAS, this association became stronger (ORivw = 0.86 [0.75–0.98], p = 0.021) (Supplementary Table S10). No association was observed for age at menarche or menopause regardless of the method and stratification.
Gynecological disorders and DTC
Genetically predicted endometrial cancer was significantly associated with an increased risk of DTC (ORivw per one SD-increase = 1.15 [1.04–1.28], p = 0.009); this finding was consistent using additional MR sensitivity analyses and in females (Supplementary Tables S6, S7, and Table S10). The genetic correlation between DTC and endometrial cancer was estimated to be r2 = 0.13 (p = 0.167), indicating a modest positive relationship between the genetic factors influencing the two cancers. Although this result suggests some shared genetic etiology, the MR-Egger did not detect significant horizontal pleiotropy (PEgger intercept = 0.43).
For uterine fibroids, MR-Egger revealed evidence of horizontal pleiotropy (PEgger intercept = 0.038) and a significant positive association with DTC (OREgger per one SD-increase = 1.87 [1.23–2.87], p = 0.009). However, these findings were not supported by other MR sensitivity analyses that showed an inverse association according to the median and mode methods (Supplementary Fig. S1 and S2). Moreover, the CAUSE method showed found no evidence of bias due to correlated pleiotropy (ORCAUSE = 1.04 [0.98–1.10]) (Supplementary Fig. S3). There was substantial heterogeneity across variants for uterine fibroids (Cochran’s Q-statistic, p = 2.14 × 10−6) (Supplementary Table S5). Upon the removal of outliers (rs149934734 in C11orf65 gene, rs547025 in SIRT3 gene, and rs78378222 in TP53 gene), the MR-PRESSO showed an inverse association (ORPRESSO = 0.94 [0.83–1.07]) (Fig. 1), with a significant difference between estimates before and after their removal (distortion test, p = 0.0055). The association was stronger and more consistent across MR sensitivity analyses in females under 50 than over 50 (females below 50 years: ORIVW = 1.52 [1.08–2.15], p = 0.017 Supplementary Table S11 and S12). No significant association was observed between endometriosis, pelvic organ prolapse, and polycystic ovary syndrome.
Reverse MR analyses (Table 3) showed no significant causal association for SHBG, endometrial cancer, or uterine fibroids, supporting the validity of our primary hypothesis.
Reverse Mendelian Randomization of Significant Exposure in Mendelian Randomization
Estimates: for binary exposures: OR [CI]; for continuous exposures: beta [CI].
IVW, inverse-variance weighted (random-effect); CI, confidence interval; SHBG, sex hormone binding globulin; SNP, single-nucleotide polymorphism; p-het. p for heterogeneity (IVW); p-pleio., p for pleiotropy(MR-Egger).
Discussion
Numerous observational studies have been conducted to examine the causal association of sex hormones and reproductive factors with DTC risk; however, the findings were weak and inconsistent across studies 33,34 due to varying study design, population, and potential detection bias caused by increased surveillance. We identified a potential link between SHBG levels and DTC risk. After adjusting for confounding factors such as TSH and BMI, the association persisted with a similar magnitude but was no longer statistically significant. Bioavailable testosterone showed a positive but nonsignificant association with DTC in MVMR adjusted for BMI. Moreover, females with a history of endometrial cancer or uterine fibroids (under 50) had an increased DTC risk, but this analysis could not be adjusted for BMI, due to restriction of accessing to the overall GWAS data.
Sex hormones
Genetically predicted SHBG was observed to be potentially associated with a 13% increased risk of DTC for one SD increase. SHBG, primarily produced by the liver and released into the bloodstream, serves as glycoprotein for transporting sex hormones, such as estrogen and testosterone. Testosterone has a stronger binding affinity to SHBG than estrogen, leading to lower SHBG levels in males than females. 35 Regulation of serum SHBG levels involves not only androgens (testosterone and dihydrotestosterone) and estrogens but also thyroid hormones. 36 Thyroid hormones indirectly enhance SHBG production by modulating HNF-4α gene expression and decreasing cellular palmitate levels in hepatocytes. 32 A study involving 72,167 individuals of European descent revealed an inverse causal association between genetically predicted TSH levels and thyroid cancer, with an odds ratio of 0.47. 37 Additionally, a recent MR study showed that a 1 SD decrease in TSH was associated with a 1.332 nmol/L increase in SHBG. 38 These findings suggest that TSH may act as a confounder of the association between SHBG and DTC. Interestingly, in our study, SHBG adjusted on TSH exhibited a stronger association with DTC (ORMVMR = 1.27 [1.00–1.63], p = 0.052). Obesity, known as a risk factor of DTC, 39 is highly correlated with several sex hormones; 40 in particular, obesity has been associated with reduced levels of SHBG. 41 Therefore, obesity was considered as a potential confounder of the association between SHBG and DTC; we observed a weakened association between SHBG and DTC when adjusting for obesity related factors through MVMR. After adjusting for both BMI and TSH, SHBG remains a potential risk factor for DTC, though nonsignificant, indicating a possible independent association between SHBG and DTC. We also considered other thyroid hormones and conditions such as hypothyroidism and hyperthyroidism, as potential confounders; however, we did not find any significant association with DTC.
In contrast to a recent MR study 42 that demonstrated an inverse association with testosterone and risk of DTC without accounting for BMI, our MVMR analysis reveals a positive association between bioavailable testosterone and DTC risk after adjustment for BMI, highlighting that SNPs used as IVs for bioavailable testosterone were also associated to BMI and a confounding role of BMI in the association. Adjusting for WHR instead of BMI showed opposing effects between bioavailable testosterone and DTC risk, suggesting that BMI and WHR may measure different aspects of obesity. This is in accordance with previous studies that reported that BMI is negatively associated with testosterone, 43 whereas WHR is positively associated. 44
Reproductive traits
Previous epidemiological studies have explored the relationship between reproductive factors as potential risk factors for DTC 10 , 33 , 45 , 46 including age at menarche, age at menopause, and parity. Studies have reported conflicting findings regarding the impact of age at menarche on thyroid cancer risk. A twofold higher risk of thyroid cancer was observed in association with higher number of reproductive years, calculated as the difference between age at menopause and age at menarche. Considering the duration of reproductive years would be of interest but we did not have access to these GWAS data.
We found an inverse association between age at first birth and risk of DTC when excluding FinnGen study. The underlying biological mechanisms could be attributed to the potential decline in sex hormone levels in older females, 47 which may influence thyroid function. Moreover, early sexual behavior and teenage pregnancy is strongly related to lower socioeconomic status. 48 Behavioral traits exhibit significant genetic correlations, particularly in female, where age at first birth and educational attainment have a correlation of 0.74 ± 0.01. 49 We remain cautious in interpreting the estimates from the age at first birth GWAS, as they may be influenced by socioeconomic and behavioral biases.
Gynecological disorder
Hysterectomy, the surgical removal of a female uterus, is often recommended for conditions such as endometrial cancer, endometriosis, and uterine fibroids. Some epidemiological studies have suggested that undergoing a hysterectomy may be associated with an increased risk of DTC. 11 We reported a positive association between uterine fibroids and DTC risk in females younger than 50 years of age. This result aligns with a previous epidemiological study that reported that female with uterine fibroids were associated with a significantly increased risk of thyroid cancer. 50 However, we could not perform a MVMR analysis to account for obesity, a known risk factor for uterine fibroids, 51 due to limited access to the GWAS summary statistics. Without this adjustment, the observed positive association may be influenced by the effects of obesity rather than solely attributable to uterine fibroids.
We revealed a significant positive association with endometrial cancer, a condition for which obesity and high levels of estrogen are known risk factors. 52 Given the potential shared risk factors between endometrial cancer and thyroid cancer, investigating the presence of pleiotropy, where the same genetic variants influence both cancers, is necessary. It should be noted that the presence of horizontal pleiotropy was tested using the MR-Egger test which may lack power. 25 We estimated the genetic correlation between these two traits and reported a modest correlation of r2 = 0.13. However, it still remains uncertain whether pleiotropy genuinely plays a role or if the observed correlation is a result of random variation. Further research using methods such as colocalization analysis and fine-mapping could provide valuable insights into the genetic relationship between these conditions.
Strengths and limitations
The strength of our study is the use of the most extensive GWAS summary statistics of DTC in the European population conducted to date. Leveraging individual-level data allowed us to perform stratified analyses based on sex, age, and histology; moreover, we could implement rigorous quality controls by removing the relatedness among participants, which enhances the precision and accuracy of our GWAS before conducting MR studies. The MR approach addresses the potential confounding factors and reverse causation commonly issued in observational studies. Moreover, in our two-sample setting, we anticipated that any bias arising from dataset overlap would tend to be toward a null effect. Given potential risk of pleiotropy due to the diverse or uncertain biological pathway (i.e., confounding factors) of many selected SNPs, we conducted multiple sensitivity analyses through MR-Egger, WM, mode methods, MR-PRESSO, and MVMR.
Our study has several limitations that should be considered. Due to restrictions in the publicly available dataset, we were unable to perform the stratified analyses including all DTC GWAS while restricting this analysis to female. Even though a majority (76.2%) of our DTC cases were female, our results may be biased since our exposure dataset primarily focused on female hormone and reproductive factors derived solely from female participants. We attempted stratification analysis on datasets with individual information available, which allowed us to assess the consistency of associations but inevitably leads to reduced statistical power along with a smaller sample size. Secondly, certain reproductive risk factors, such as age at first birth, menarche, and menopause relied on self-reported data. This may introduce potential recall bias and measurement errors. Moreover, a few exposures exhibit low power due to insufficient genetic variation in the IVs or a small sample size, limiting our ability to draw reliable causal inference. We were also unable to rule out that some SNPs used as IVs were also associated with BMI and TSH, violating the second assumption of MR analysis. Lastly, our analysis was restricted to individuals of European ancestry, which limit the generalizability of our findings to nonEuropean populations.
In conclusion, our MR study highlights that SHBG emerged as a possible causal risk factor, while the association between bioavailable testosterone and DTC became positive after adjusting for BMI, hinting at a confounding effect of BMI. A significant association was observed between a history of endometrial cancer and increased risk of DTC; however, a possible shared genetic predisposition between the two cancers is suspected. Uterine fibroids showed a plausible causal association with DTC, particularly in younger female under 50 years. We could not provide definitive evidence regarding the potential role of sex hormones and reproductive factors in observed sex discrepancies of DTC risk. Future research on female population is essential to clarify the underlying biological mechanisms of contributing to the risk of DTC.
Footnotes
Acknowledgments
The authors acknowledge the use of data and biological samples from the EPIC-Ragusa cohort, principal investigator Rosario Tumino; EPIC-Asturias, principal investigator José Ramón Quirós García; EPIC-Bilthoven, principal investigator Monique Verschuren, EPIC-Utrecht, principal investigator Roel Vermeulen.
The authors acknowledge deCODE Genetics, FinnGen, ReproGen consortium and LIFE-HEART for sharing their GWAS summary statistics. This research has been conducted using the UK Biobank Resource under application number 92392. This work is also part of the Inserm Cross-Cutting Project GOLD.
Authors’ Contributions
S.H.P.: Study design, analyses, and writing the original draft. P.-E.S.: Interpretation of results, editing, and approval. Y.A.: Interpretation of results, editing, and approval. M.K.: Interpretation of results, editing, and approval. R.K.: Data acquisition, interpretation of results, editing, and approval. R.T.F.: Data acquisition, interpretation of results, editing, and approval. M.S.: Data acquisition, interpretation of results, editing, and approval. C.A.: Data acquisition, interpretation of results, editing, and approval. F.P.: Data acquisition, interpretation of results, editing, and approval. C.S.: Data acquisition, interpretation of results, editing, and approval. M.R.-B.: Data acquisition, interpretation of results, editing, and approval. A.A.: Data acquisition, interpretation of results, editing, and approval. N.C.C.: Data acquisition, interpretation of results, editing, and approval. M.G.: Data acquisition, interpretation of results, editing, and approval. S.T.T.: Interpretation of results, editing, and approval. E.W.: Interpretation of results, editing, and approval. F.d.V.: Data acquisition, interpretation of results, editing, and approval. F.L.: Interpretation of results, editing, and approval. P.G.: Data acquisition, interpretation of results, editing, and approval. C.M.: Data acquisition, interpretation of results, editing, and approval. P.L.-P.: Data acquisition, interpretation of results, editing, and approval. E.O.: Data acquisition, interpretation of results, editing, and approval. A.B.-A.: Data acquisition, interpretation of results, editing, and approval. J.-F.D.: Data acquisition, interpretation of results, editing, and approval. H.T.: Data acquisition, interpretation of results, editing, and approval. A.F.: Data acquisition, interpretation of results, editing, and approval. R.E.: Data acquisition, interpretation of results, editing, and approval. F.G.: Data acquisition, interpretation of results, editing, and approval. S.L.: Data acquisition, interpretation of results, editing, and approval. Sabina Rinaldi: Interpretation of results, editing, and approval. A.E.: Interpretation of results, editing, and approval. C.D.: Study design, interpretation of results, editing, and approval. T.T.: Funding acquisition, study design, supervision and interpretation of results, editing, and approval.
Disclaimer
Where authors are identified as personnel of the International Agency for Research on Cancer/World Health Organization, the authors alone are responsible for the views expressed in this article and they do not necessarily represent the decisions, policy, or views of the International Agency for Research on Cancer/World Health Organization.
Data Availability Statement
The data supporting this study are available as follows: EPIC, EPITHYR, and the Italian Study data can be requested from the corresponding author, T.T., but are not publicly available due to General Data Protection Regulation restrictions. GWAS summary statistics from the UK Biobank can be accessed through the UK Biobank website upon request (https://www.ukbiobank.ac.uk/), FinnGen GWAS data are publicly available at https://www.finngen.fi/en and summary statistics from deCODE genetics can be accessed at ![]() . The R scripts used for MR analyses can be provided upon reasonable request to the corresponding author, and the SNPs utilized for this analyses are available in Supplementary Table S3.
. The R scripts used for MR analyses can be provided upon reasonable request to the corresponding author, and the SNPs utilized for this analyses are available in Supplementary Table S3.
Author Disclosure Statement
None of the authors have any disclosure to report or any competing financial interest.
Funding Information
S.H.P. was the recipient of a PhD fellowship from Paris-Saclay University. The coordination of EPIC is financially supported by International Agency for Research on Cancer and also by the Department of Epidemiology and Biostatistics, School of Public Health, Imperial College London which has additional infrastructure support provided by the National Institute for Health and Care Research (NIHR) Imperial Biomedical Research Centre. The national cohorts from EPIC are supported by: Danish Cancer Society (Denmark); Ligue Contre le Cancer, Institut Gustave Roussy, Mutuelle Générale de l’Education Nationale, Institut National de la Santé et de la Recherche Médicale (France); German Cancer Aid, German Cancer Research Center (DKFZ), German Institute of Human Nutrition Potsdam-Rehbruecke, Federal Ministry of Education and Research (Germany); Associazione Italiana per la Ricerca sul Cancro-AIRC-Italy, Compagnia di SanPaolo and National Research Council (Italy); Dutch Ministry of Public Health, Welfare and Sports, Netherlands Cancer Registry, the National Institute for Public Health and the Environment (The Netherlands), LK Research Funds, Dutch Prevention Funds, Dutch ZON (Zorg Onderzoek Nederland), World Cancer Research Fund, Statistics Netherlands (The Netherlands); Health Research Fund-Instituto de Salud Carlos III), Regional Governments of Andalucía, Asturias, Basque Country, Murcia and Navarra, and the Catalan Institute of Oncology-ICO (Spain); Cancer Research UK (14136 to EPIC-Norfolk; C8221/A29017 to EPIC-Oxford), Medical Research Council (1000143 to EPIC-Norfolk; MR/M012190/1 to EPIC-Oxford). (United Kingdom). The genotyping of EPIC samples was supported by the Association pour la Recherche sur le Cancer (ARC) (#RF20180207126). The EPITHYR genome-wide association study was funded by INCA (#9533) and ARC (#PGA120150202302). The funders had no role in the design, analysis, or writing of this article. T.T.: Funding acquisition. The rest of the authors have no funding information available.
Supplementary Material
Supplementary Data S1
Supplementary Data S2
Supplementary Data S3
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
Supplementary Table S9
Supplementary Table S10
Supplementary Table S11
Supplementary Table S12
Supplementary Table S13
Supplementary Table S14