
Abstract
Background:
The 14q13.3 has been identified as a genetic locus associated with a genetically increased risk of papillary thyroid cancer (PTC) in several cohorts, yet its underlying regulatory mechanisms remain poorly understood.
Methods:
The full-length sequence of expressed sequence tag fragment AA632637 in the thyroid was obtained by rapid amplification of complementary DNA ends assay. Quantitative Reverse Transcription PCR (qRT-PCR) assays were utilized to examine the expression levels of the long noncoding RNA (lncRNA) in clinical thyroid tissues and cell lines. Functional assays, including cell proliferation, migration, invasion, and apoptosis assays, were conducted both in vitro and in vivo. Furthermore, RNA-seq analysis, actinomycin D assay, RNA pull-down, RNA immunoprecipitation, and dual-luciferase reporter assays were performed to identify the long noncoding RNA (lncRNA) binding targets and reveal the underlying regulatory mechanism.
Results:
We identified a previously unannotated lncRNA gene, named papillary thyroid carcinoma susceptibility candidate 3 antisense 1 (PTCSC3-AS1), within 14q13.3. The expression of PTCSC3-AS1 was strongly downregulated in PTC tumor tissues, and restoration of PTCSC3-AS1 expression in PTC cells inhibited tumorigenesis and promoted cell apoptosis. Moreover, PTCSC3-AS1 and PTCSC3, two lncRNAs located on the opposite strands at 14q13.3, were revealed to synergistically interact with their shared binding protein vimentin. Forced overexpression of PTCSC3 and PTCSC3-AS1 revealed that ZC3H12A, a gene validated as a PTC suppressor, was the shared downstream target of the two lncRNAs. Vimentin significantly reduced the mRNA stability of ZC3H12A, while the upregulation of PTCSC3 and PTCSC3-AS1 suppressed the mRNA degradation of ZC3H12A. In addition, rs944289 and rs116909374 were identified as two potential causative variants with distinct regulatory roles in the 14q13.3 locus. Mechanistically, PTCSC3-AS1 and PTCSC3 protected ZC3H12A from vimentin-mediated mRNA degradation by targeting the ZC3H12A 3′ untranslated region (3′UTR) during PTC initiation and progression.
Conclusion:
Our results suggest a novel dual-lncRNA regulatory model in the 14q13.3 risk locus and provide a comprehensive annotation of the PTCSC3-AS1/PTCSC3-vimentin-ZC3H12A signaling network in PTC genetic predisposition.
Introduction
The genetic predisposition to papillary thyroid cancer (PTC) is determined by multiple genomic variants in typical loci, such as 14q13.3, which exhibits a strong association with PTC risk. The 14q13.3 predisposition locus has been identified across European, American, Chinese, and Japanese populations by genome-wide association studies (GWAS). Two genetic polymorphism variants, rs944289 and rs116909374 were identified as the representative tag single nucleotide polymorphisms (SNPs) within this locus. A subsequent study validated that rs944289, a causative promoter variant, can affect the binding affinities of CCAAT enhancer binding protein alpha (C/EBPα) and CCAAT enhancer binding protein beta (C/EBPβ) transcription factors thereby regulating the expression of long noncoding RNA (lncRNA) gene papillary thyroid carcinoma susceptibility candidate 3 (PTCSC3). 5 However, the functional variants and target gene networks in the other linkage disequilibrium (LD) block containing rs116909374 have not been clearly annotated yet. Furthermore, the precise mechanism by which the 14q13.3 locus confers PTC predisposition remains elusive.
LncRNAs are RNA molecules with more than 200 nucleotides in length. Recent studies have characterized lncRNAs as molecular decoys, guides, and scaffolds that participate in multiple biological processes through transcriptional or posttranscriptional regulation mechanisms. 6 Most nuclear lncRNAs function in chromatin organization, RNA transcription, and splicing, while cytoplasmic lncRNAs have been reported to regulate mRNA stability, transportation, and translation. 7 The functions of lncRNAs are closely linked to their interactions with molecules, such as DNA elements, RNA, and RNA-binding proteins. 8 Dysregulation of lncRNAs significantly impacts cancer hallmarks, acting as either oncogenes or tumor suppressors. 9 Certain tissue-specific lncRNAs identified in thyroid cancer risk loci, such as lncRNAs PTCSC2 (at 9q22.33) 10 and PTCSC3 (at 14q13.3) 5 have been implicated in PTC malignancy. For instance, PTCSC2 participates in the forkhead box E1 (FOXE1)-mediated p53 signaling pathway via its interaction with RNA-binding protein myosin heavy chain 9 (MYH9). 11 Similarly, PTCSC3 downregulates S100A4, leading to a reduction of vascular endothelial growth factor A (VEGF) and matrix metallopeptidase 9 (MMP-9), which in turn decreases PTC cell motility and invasiveness. 12
Here, in an effort to investigate the underlying regulatory mechanism in PTC predisposition, we provide a detailed annotation of the featured genomic variants/elements and the functional molecules in the 14q13.3 PTC risk locus. Notably, a previously uncharacterized lncRNA named PTCSC3 antisense 1 (PTCSC3-AS1) was identified and functionally characterized. In particular, our study highlights that the crosstalk between lncRNAs PTCSC3 and PTCSC3-AS1 modulates the expression of ZC3H12A through a competitive binding system mediated with vimentin at the 14q13.3 PTC risk locus.
Materials and Methods
Detailed information regarding experimental materials, protocols, data analysis methods, and data availability statement is provided in the Supplementary Data.
Results
Identification and characterization of an unannotated lncRNA in the 14q13.3 locus
Previous studies mainly focused on the lncRNA PTCSC3 and its causative promoter variant rs944289 to explain the PTC genetic susceptibility of the 14q13.3 locus. However, considering the complexity of this risk locus, the regulatory model relying on a single causative variant and a single susceptibility gene could not provide a complete annotation of the underlying mechanism of PTC risk. Thus, we performed a detailed screening across a reported 40 kb LD block containing GWAS tag SNP rs944289, especially in the adjacent genomic region of PTCSC3. Consequently, AA632637, an expressed sequence tag fragment derived from thyroid tissue and located on the opposite strand of PTCSC3, was highlighted (Fig. 1A). First, we confirmed that AA632637 is expressed in thyroid tissue rather than PTC cell lines using RT-PCR assays (Supplementary Fig. S1A). After performing 5′ and 3′ rapid amplification of complementary DNA ends assays (Supplementary Fig. S1B), the 281 bp full-length sequence was obtained and designated as PTCSC3-AS1. Furthermore, PTCSC3-AS1 was predicted to have a potential open reading frame (ORF) with only 31 amino acids using the ORF Finder tool (Supplementary Fig. S1C). The coding probability prediction analysis was performed using the Coding Potential Assessment Tool and we found that the output value of PTCSC3-AS1 was only 0.0026, which was much lower than those of other tested protein-coding genes and lncRNAs (Fig. 1B). It suggested that PTCSC3-AS1 is more likely a noncoding gene. Furthermore, we found that the expression level of PTCSC3-AS1 was significantly higher in rs944289 wildtype homozygous samples than in mutant homozygous samples (Fig. 1C). We subsequently examined its expression in 50 PTC tumor pairs using Quantitative Reverse Transcription PCR (qRT-PCR). Similar to PTCSC3, the expression of PTCSC3-AS1 was significantly lower in the PTC tumors than in the adjacent nontumorous tissues (Fig. 1D). RNA fluorescence in situ hybridization analysis with PTCSC3-AS1 probe in PTC tissue pairs confirmed this differential expression. It also indicated that PTCSC3-AS1 was predominantly localized in the cytoplasmic region of thyroid cells (Fig. 1E).
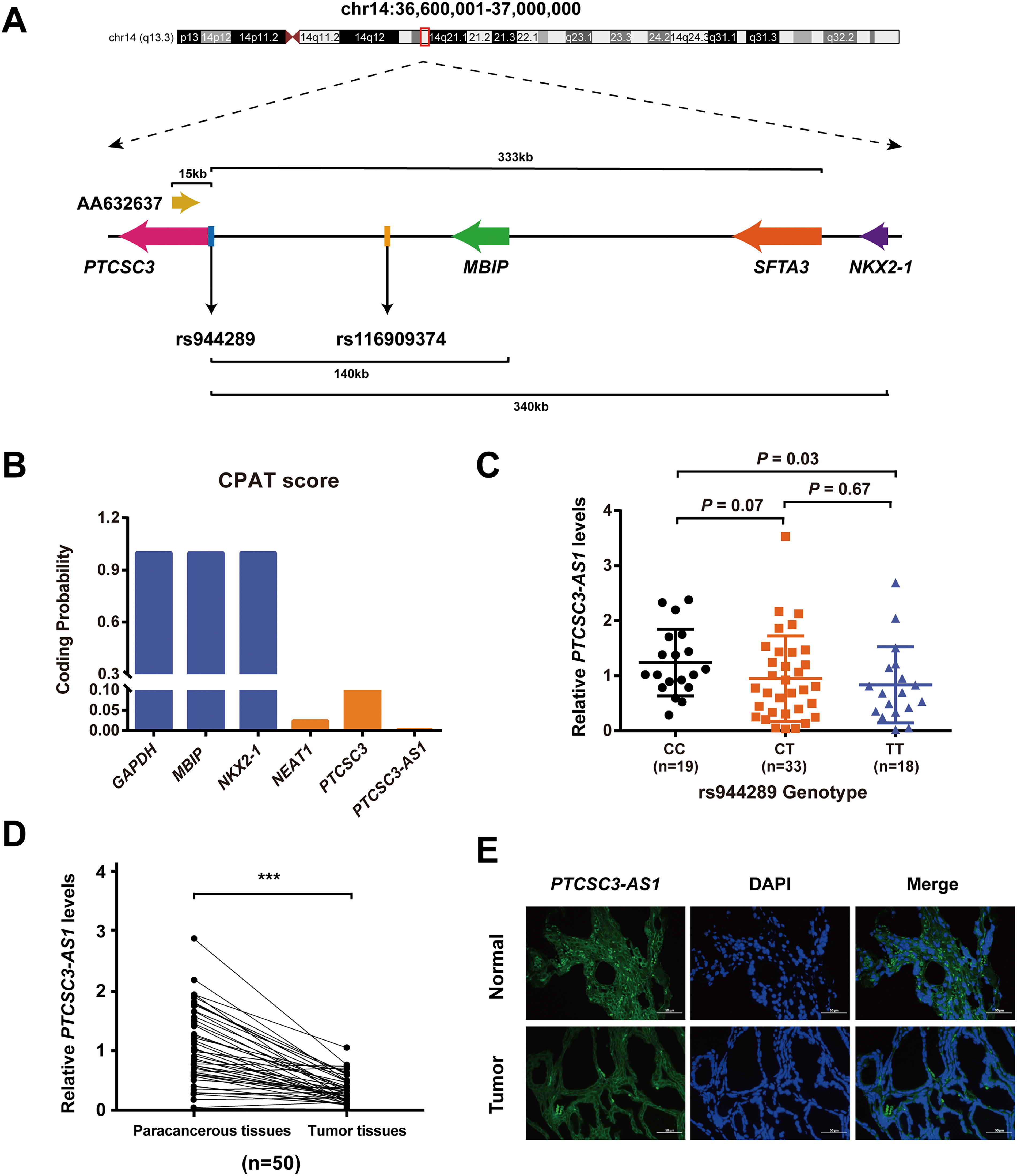
Characterization of lncRNA gene PTCSC3-AS1 in PTC.
PTCSC3-AS1 is a PTC tumor suppressor gene and can promote cell apoptosis
Since PTCSC3-AS1 is nearly undetectable in most PTC cell lines, stable overexpression cell models with forcing expression of PTCSC3-AS1 were established. It showed that PTCSC3-AS1 overexpression significantly inhibited cell proliferative capacity in both Cell Counting Kit-8 (CCK-8) (Fig. 2A) and colony formation assays (Fig. 2B). Furthermore, cell cycle analysis revealed a remarkable delay in the S phase of PTCSC3-AS1 overexpression cells (Fig. 2C). Terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) staining demonstrated a higher proportion of apoptotic cells in cells with PTCSC3-AS1 overexpression compared to the controls (Fig. 2D). In addition, PTCSC3-AS1 overexpression downregulated the antiapoptotic protein BCL2 apoptosis regulator (BCL-2) and upregulated the pro-apoptotic protein BCL2 associated X, apoptosis regulator (BAX) (Supplementary Fig. S2A). Cell migration and invasion capacities were also significantly decreased in PTCSC3-AS1 overexpression cells (Fig. 2E and F, Supplementary Fig. S2B). Consistent with the in vitro findings, overexpressing PTCSC3-AS1 significantly reduced both tumor size and weight of xenograft models and suppressed the expression of proliferation marker Ki67 (Fig. 2G–J). Taken together, PTCSC3-AS1 functions as a candidate PTC suppressor gene, capable of inhibiting PTC malignancy and inducing cell apoptosis.
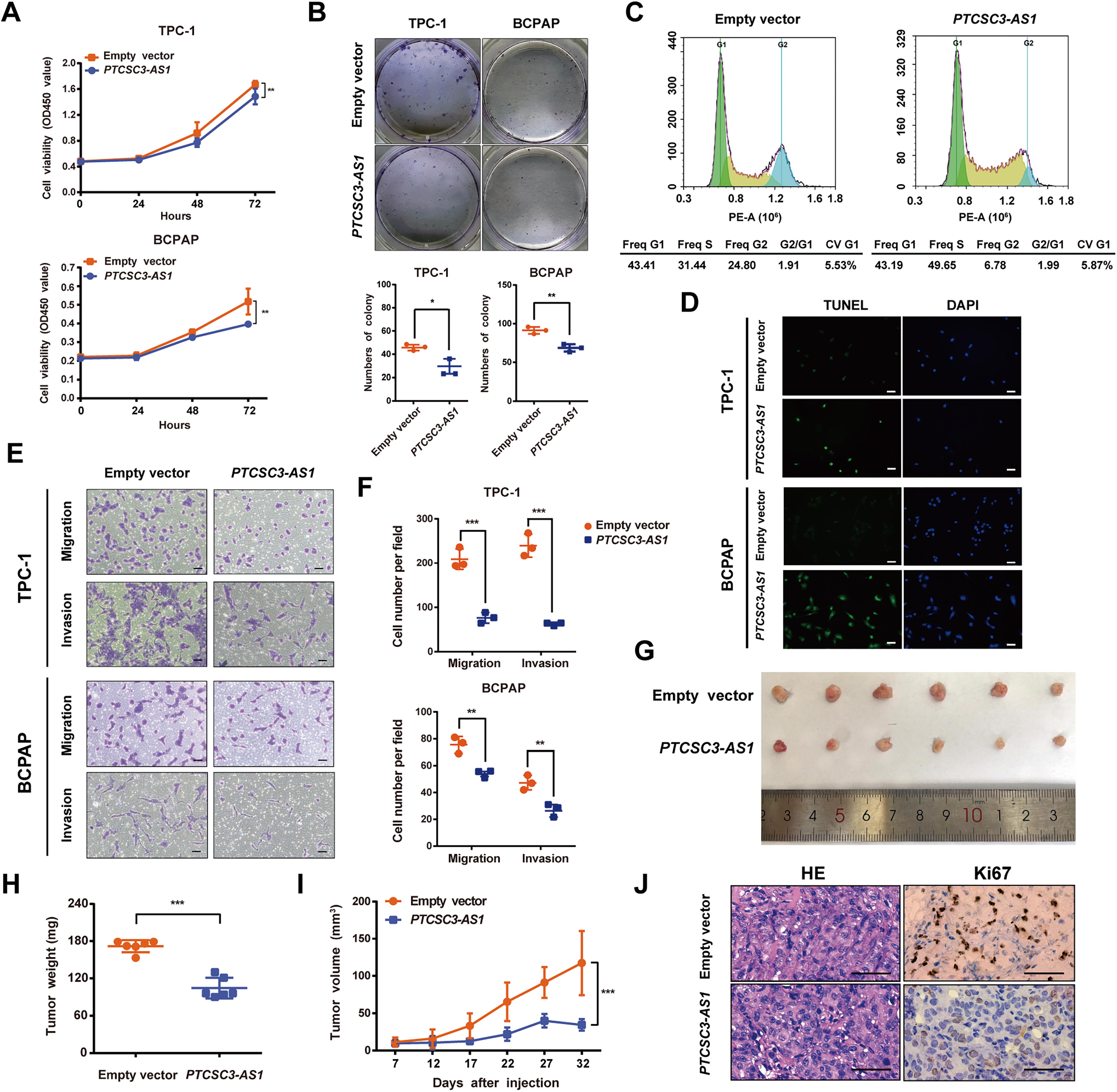
PTCSC3-AS1 functions as a tumor suppressor to inhibit PTC malignancy.
ZC3H12A is an important target synergistically upregulated by both lncRNAs PTCSC3 and PTCSC3-AS1
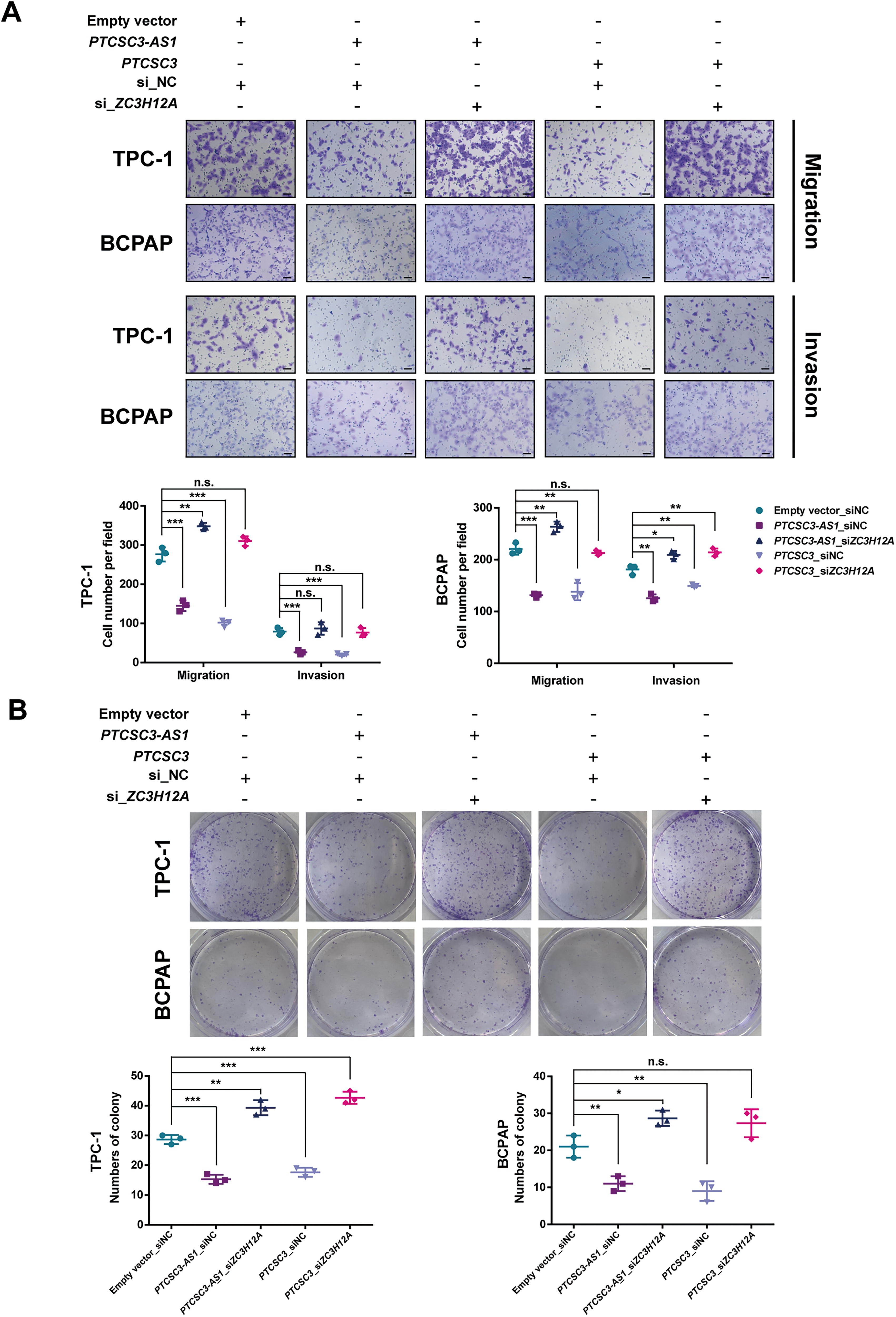
We conducted RNA-seq analysis to investigate the downstream signaling network of PTCSC3-AS1. Subsequently, differential gene expression analysis identified a total of 1012 dysregulated genes (p < 0.05). Gene Ontology enrichment analysis revealed that PTCSC3-AS1 was involved in protein binding, signal transduction, mRNA 3′-end processing, and regulation of transcription processes (Supplementary Fig. S3). Furthermore, a total of 669 dysregulated targets, including 205 upregulated genes and 464 downregulated genes, were identified with more stringent screening parameters (Fig. 3A). Notably, the expression level of PTCSC3-AS1 was positively correlated with PTCSC3 expression level in 15 PTC tumors (R = 0.3905, p = 0.0128) and nontumorous thyroid tissues (R = 0.8960, p < 0.001) (Fig. 3B). It suggests that PTCSC3 and PTCSC3-AS1 may function synergistically in downstream regulation. Since a previous study had reported 467 dysregulated genes caused by PTCSC3 overexpression in TPC-1 cells, 12 a comparison analysis was performed, and two shared target genes, ZC3H12A and CDK5R2, were discovered (Fig. 3C). Between them, ZC3H12A was highlighted due to its higher p-value. In addition, we analyzed gene expression data of thyroid tumors and normal tissue samples from the The Cancer Genome Atlas (TCGA) database. ZC3H12A expression was significantly decreased in thyroid tumor tissues, while CDK5R2 showed no expression difference (Supplementary Fig. S4). The dysregulation of ZC3H12A was confirmed by qRT-PCR (Fig. 3D) and Western blotting assays (Fig. 3E), consistent with our RNA-seq results (Fig. 3F). Moreover, in vitro functional assays were performed with forced ZC3H12A expression. Consistent with its roles in other cancer types, 13,14 our results demonstrated that ZC3H12A functions as a tumor suppressor in thyroid carcinogenesis (Fig. 3G–I). In addition, knocking down ZC3H12A could rescue the inhibitory effects on PTC malignancy caused by PTCSC3 or PTCSC3-AS1 overexpression (Fig. 4A and B). It further confirms that ZC3H12A is a downstream target of PTCSC3/PTCSC3-AS1 in PTC initiation and progression.
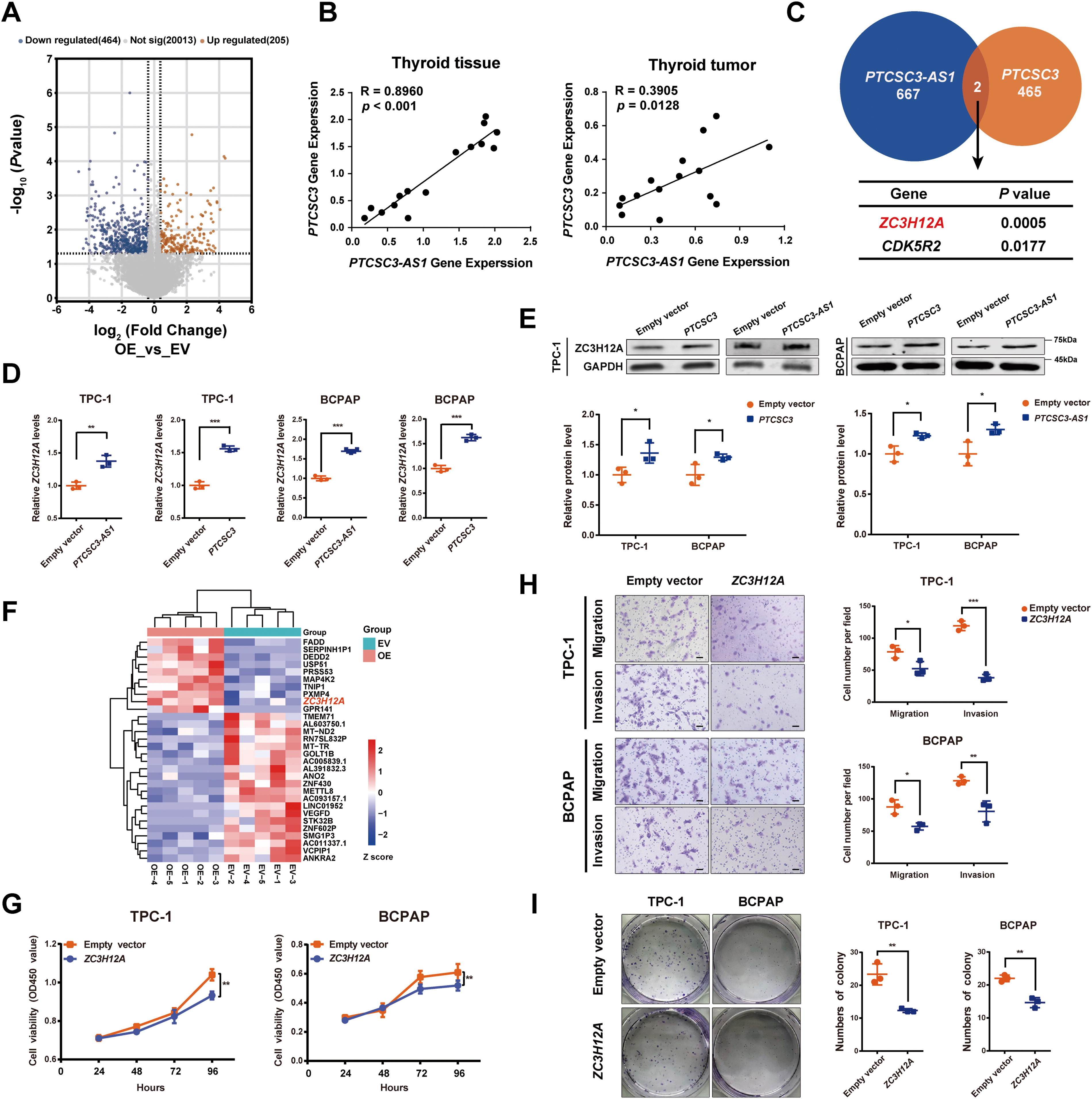
Identification of PTC suppressor gene ZC3H12A as a shared target of lncRNAs PTCSC3-AS1 and PTCSC3.
PTCSC3-AS1 and PTCSC3 can improve the mRNA stability of ZC3H12A
We found that both PTCSC3-AS1 and PTCSC3 were predominantly expressed in the cytoplasm rather than the nucleus in cells isolated from nontumorous thyroid tissues (Fig. 5A). Cytoplasmic lncRNAs are known to play regulatory roles through various molecular mechanisms, including modulating mRNA stability. 15 Since qRT-PCR analysis revealed that ZC3H12A mRNA levels were upregulated upon overexpression of PTCSC3 or PTCSC3-AS1, we hypothesized that ZC3H12A mRNA stability was influenced by dysregulation of these two lncRNAs at posttranscriptional level. Subsequent actinomycin D assays showed that ZC3H12A mRNA stability was significantly enhanced by overexpression of PTCSC3-AS1 or PTCSC3, respectively (Fig. 5B). It is well known that 3′ untranslated regions (3′UTRs) play key roles in mRNA metabolism, including the regulation of mRNA stability. Important cis-regulatory elements, which can be recognized by trans-factors such as RNA-binding proteins and microRNAs involved in mRNA stability, are enriched in 3′UTRs. 16,17 The 3′UTR is known to serve as a docking platform for regulatory proteins and RNAs. 18 Given this, we proposed that the 3′UTR of ZC3H12A could serve as a key target element for these two lncRNAs to maintain mRNA stability. Dual-luciferase reporter assay was performed using a reporter plasmid containing the 3′UTR of ZC3H12A. The results showed significantly increased relative luciferase activity upon PTCSC3-AS1 or PTCSC3 overexpression (Fig. 5C and D). Collectively, these results suggest that PTCSC3-AS1 and PTCSC3 enhance the mRNA stability of ZC3H12A by targeting its 3′UTR.

PTCSC3-AS1 and PTCSC3 promote ZC3H12A expression by targeting its 3′UTR region.
Vimentin serves as the lncRNA binding protein of PTCSC3-AS1 and PTCSC3 to regulate ZC3H12A via targeting its 3′UTR region
Since lncRNAs function as miRNA sponges in the competitive endogenous RNAs (ceRNAs) model, 19 we first attempted to identify a shared candidate miRNA that could potentially bind to PTCSC3, PTCSC3-AS1, and ZC3H12A. However, we were unable to find such a miRNA after screening the miRDB database (https://mirdb.org/download.html). On the contrary, we found that lncRNA PTCSC3-AS1 and the 3′UTR of ZC3H12A were predicted to have potential miR-4282 binding sites (Fig. 6A, Supplementary Tables S3 and S4), which might suggest an alternative model involving miR-4282. To validate this, we conducted dual-luciferase reporter assays using plasmids containing PTCSC3-AS1 and ZC3H12A-3′UTR, respectively. A significant decrease in luciferase activity was observed in the ZC3H12A-3′UTR reporter gene group treated with miR-4282 (Fig. 6B). However, the PTCSC3-AS1 reporter gene group did not exhibit any significant difference when comparing the values of miR-4282 treated samples with the negative controls. It suggested that miR-4282 might not participate in the upregulation of ZC3H12A mRNA stability caused by PTCSC3-AS1 overexpression. Therefore, we excluded the ceRNA regulatory model from our mechanism hypothesis. We also performed an RNA pull-down assay to isolate PTCSC3-associated proteins from thyroid tissue extraction. As a result, two separated protein bands were identified as DExH-box helicase 9 (DHX9) and vimentin by mass spectrometry analysis (Fig. 6C and D, Supplementary Fig. S5, Supplementary Tables S5 and S6). However, it showed that ZC3H12A expression was not significantly changed with DHX9 knocking-down (Fig. 6E). Moreover, there was no significant difference in transcriptional activity of ZC3H12A-3′UTR treated with DHX9 knocking-down (Fig. 6F). It indicated that DHX9 might not be the key protein involved in the regulation of ZC3H12A.

Identification of vimentin and DHX9 proteins as candidate PTCSC3-AS1/PTCSC3 binding partners.
To confirm the binding capacity of vimentin with PTCSC3, an RNA Immunoprecipitation (RIP) assay was performed. Significant enrichment of PTCSC3 was found in the vimentin antibody pull-down group (Fig. 7A). Furthermore, we discovered that vimentin significantly inhibited ZC3H12A expression (Fig. 7B). Consistently, the forced overexpression of vimentin can also significantly reduce the mRNA stability of ZC3H12A (Fig. 7C) and the transcriptional activity of luciferase reporter plasmids containing the ZC3H12A-3′UTR (Fig. 7D). These results suggest that vimentin may accelerate mRNA decay to inhibit ZC3H12A by targeting its 3′UTR region.
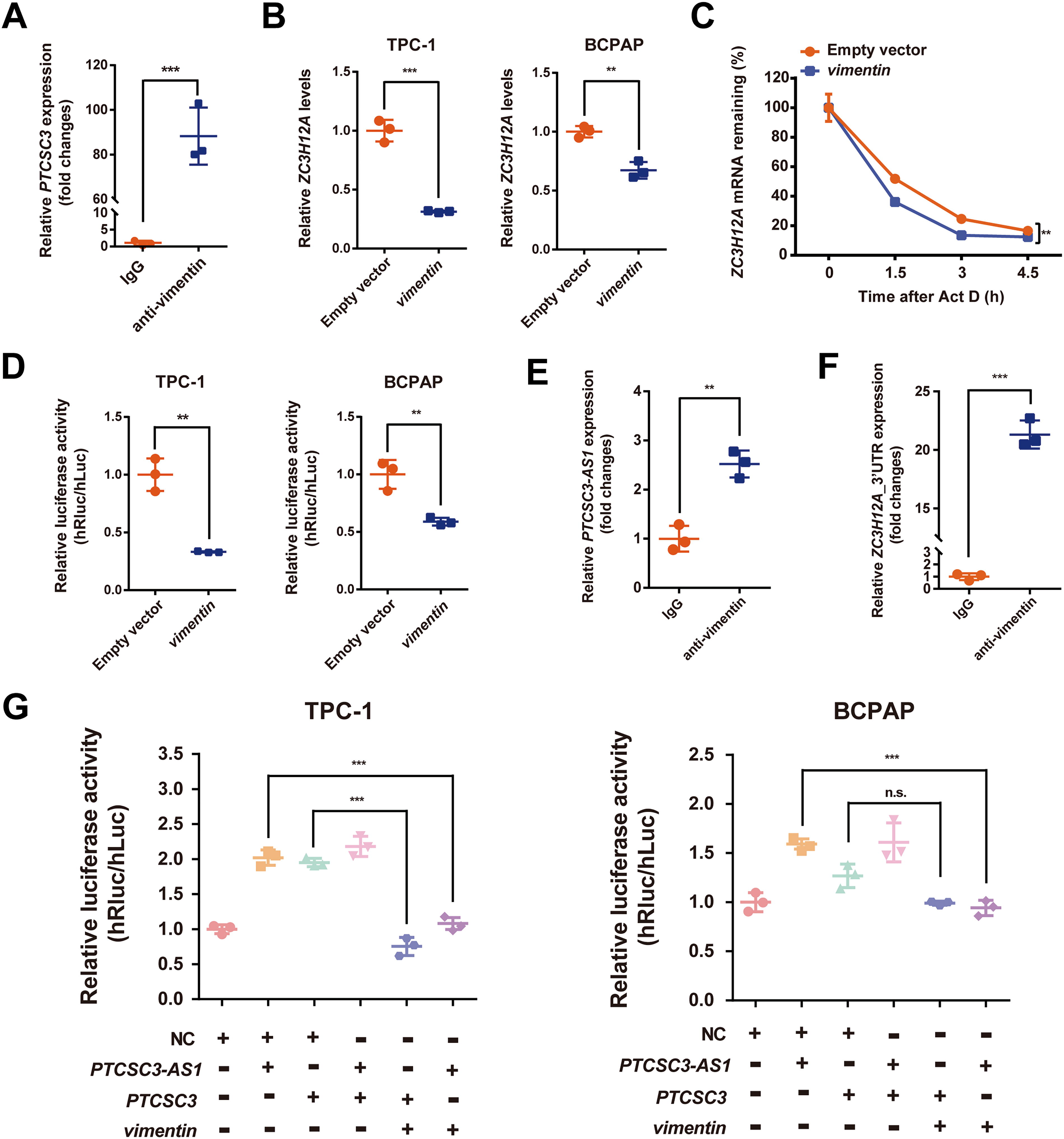
PTCSC3-AS1 and PTCSC3 promote ZC3H12A expression by sequestering vimentin protein from its 3′UTR region.
Furthermore, significant enrichment of PTCSC3-AS1 and 3′UTR of ZC3H12A was detected in the immunoprecipitation products by RIP assay using antibody against vimentin (Fig. 7E and F). A functional rescue experiment was conducted using luciferase reporter assay, which showed that the overexpression of vimentin significantly attenuated the luciferase activity of ZC3H12A-3′UTR induced by PTCSC3 or PTCSC3-AS1 (Fig. 7G). We also examined the expression of vimentin in response to PTCSC3 or PTCSC3-AS1 overexpression using Western blotting assays. The results showed that vimentin expression remained unaltered under the dysregulation of these two lncRNAs (Supplementary Fig. S6). It further confirms that PTCSC3 and PTCSC3-AS1 function as binding partners of vimentin protein, rather than affecting its expression levels. Taken together, our study discovers that PTCSC3-AS1 and PTCSC3 play as negative regulators of vimentin protein, limiting its inhibitory capacity to interact with ZC3H12A in PTC progression.
rs944289 and rs116909374 are two potential causative variants with distinct regulatory roles for the candidate targets in 14q13.3 locus
We also found that the two independent GWAS tag SNPs at 14q13.3, rs944289, and rs116909374, were in different LD regions in various populations (Fig. 8A and B, Supplementary Fig. S7), which implied they might play distinct regulatory roles in PTC predisposition. rs944289 had been previously shown to affect PTCSC3 promoter activation. Here, we identified that rs116909374 was located in an open chromatin region (Supplementary Fig. S8) and was not genetically linked to other nearby SNPs in the 1000 Genome datasets (https://pubs.broadinstitute.org/mammals/haploreg). In addition, based on the expression quantitative trait locus data of thyroid in 14q13.3 locus from the Genotype-Tissue Expression project (GTEx) database (www.gtexportal.org), significant associations with PTCSC3 and MBIP were observed for rs944289 (Fig. 8C). Moreover, significant associations with PTCSC3, MBIP, and surfactant associated 3 (SFTA3) were also observed for rs116909374. To further validate their potential long-range regulatory effects, dual-luciferase reporter assays were conducted using a reporter plasmid containing the E4TATA mini promoter. It indicated that the rs944289 fragment exhibited transcriptional activation function (Fig. 8D), while rs116909374 was in a transcriptional inhibitory element (Fig. 8E). Furthermore, the mutant risk (T) allele of rs944289 significantly downregulated transcriptional activity (Fig. 8D), whereas the mutant risk (T) allele of rs116909374 could significantly upregulated transcriptional activity (Fig. 8E).
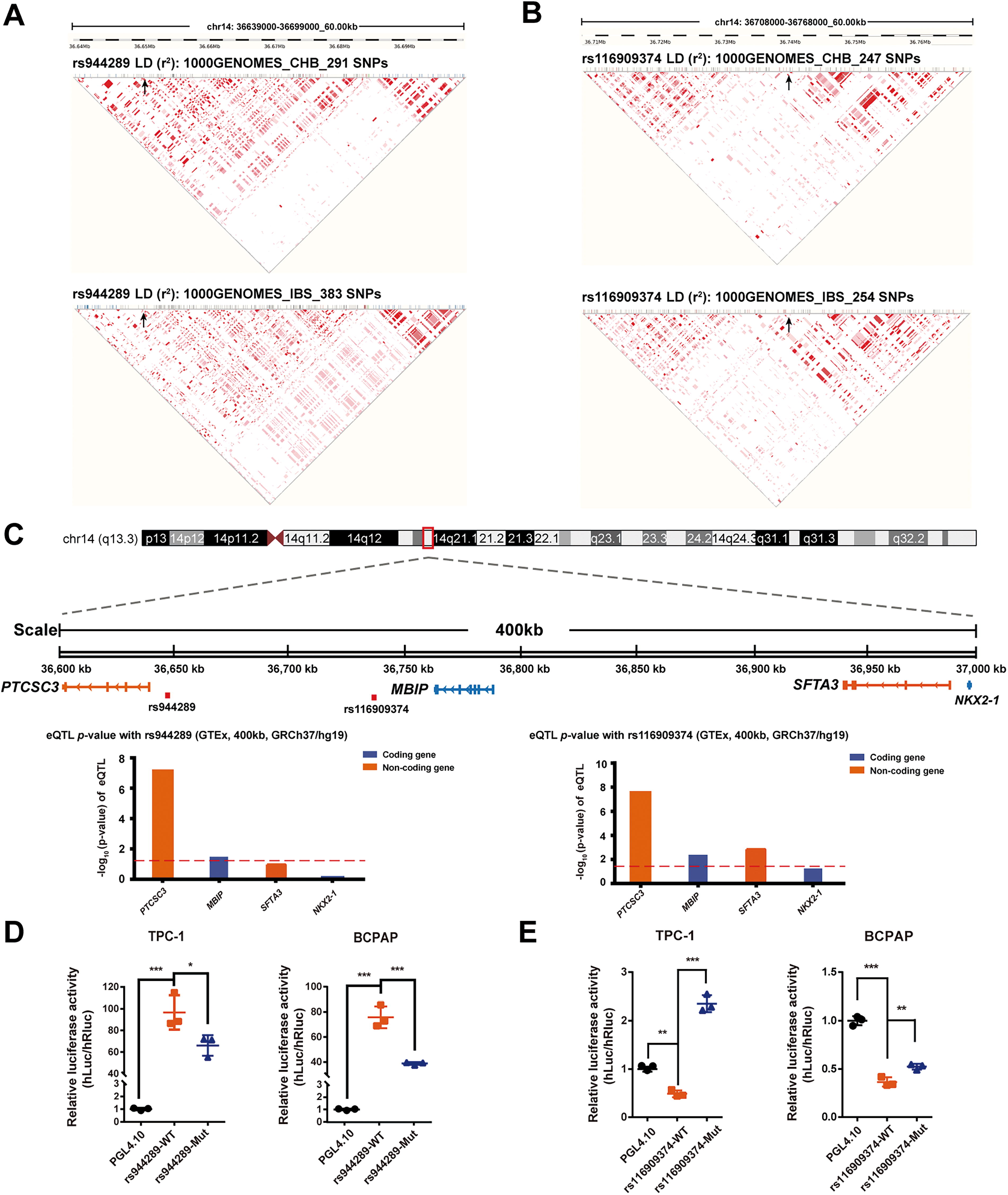
Distinct regulatory effects of rs944289 and rs116909374 on the candidate targets in the 14q13.3 locus. LD analysis results in 14q13.3 for rs944289
Moreover, it was found that MBIP and STFA3 showed significantly higher expression in the PTC tumors from the TCGA database (Supplementary Fig. S9). We hypothesized that the rs116909374 fragment might play a transcriptional inhibitory role by primarily targeting the two candidate oncogenes MBIP and SFTA3. Meanwhile, the rs944289 fragment may play a transcriptionally activating role by primarily targeting PTCSC3 and PTCSC3-AS1. Future work is required to investigate the physical interactions within the genomic network at this locus. In conclusion, our study has provided a comprehensive annotation of the 14q13.3 thyroid cancer risk locus and has proposed a dual-lncRNA regulatory model underlying vimentin-mediated regulation of ZC3H12A mRNA stability (Fig. 9).
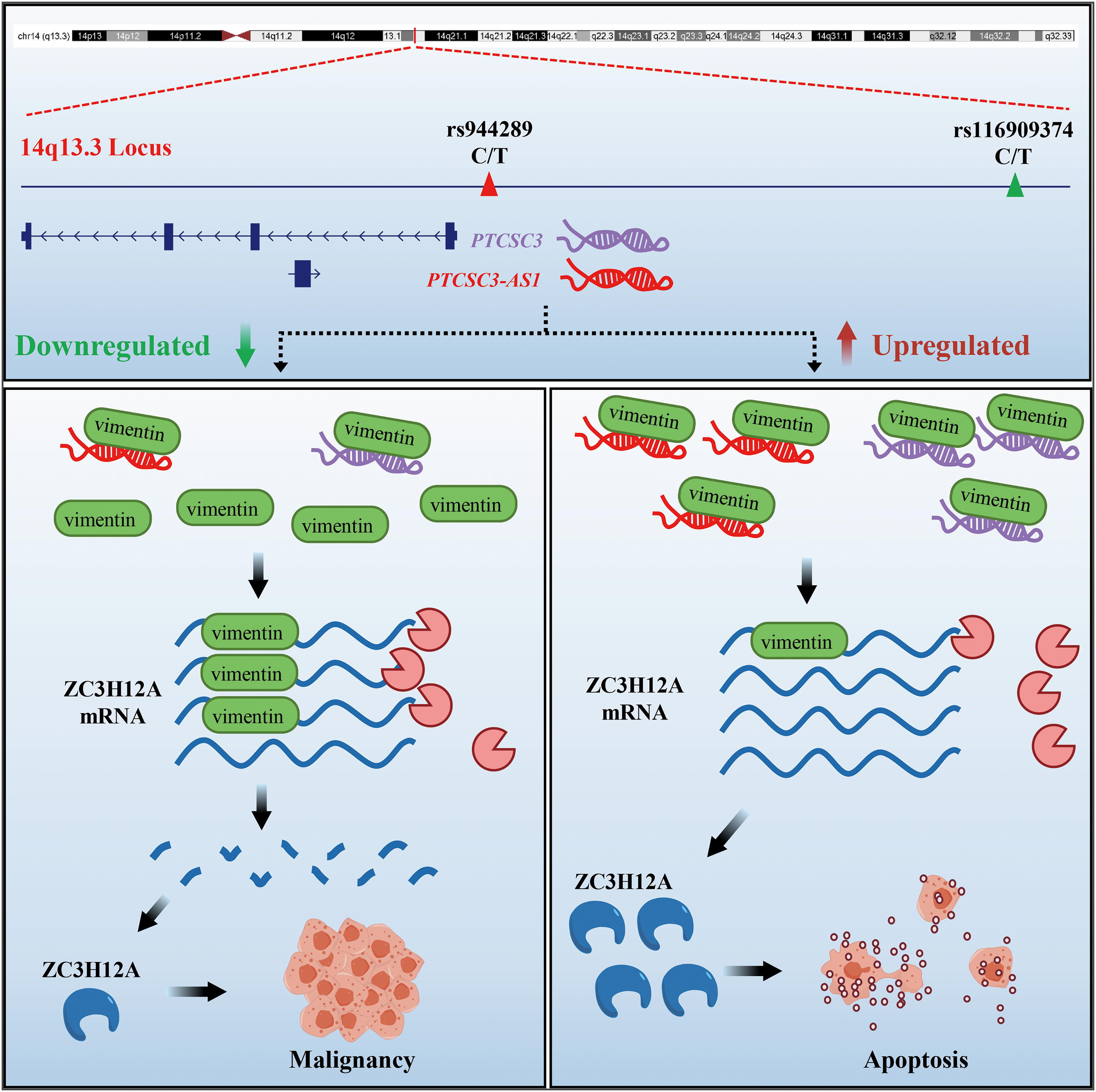
Schematic summary of the key elements and molecular mechanism presented in our study.
Discussion
14q13.3 locus has been reported as one of the top loci of PTC genetic predisposition. 20 In this study, we provided a comprehensive annotation of 14q13.3 locus, including characterization of an antisense lncRNA PTCSC3-AS1 and elucidation of the detailed regulatory mechanism of PTCSC3 and PTCSC3-AS1 in vimentin-mediated regulation of ZC3H12A.
PTCSC3 has been reported to have multiple roles in suppressing the oncogenic growth signals of PTC. 5 For instance, PTCSC3 was demonstrated to suppress thyroid cancer cell aggressiveness by modulating S100A4, VEGF, and MMP-9 expression. PTCSC3 also correlates with the expression of NRG1, another candidate PTC predisposition gene harboring GWAS tag SNP rs2439302. 4,21 Moreover, it was also reported that overexpression of PTCSC3 remarkably suppressed the Warburg effect by directly interacting with PGK1 in thyroid cancer cells. 22 In the present study, we identified an unannotated lncRNA PTCSC3-AS1 transcribed from the complementary strand of PTCSC3 and located within the same LD block. Antisense RNAs are well known to influence gene expression through various steps, including transcriptional initiation, cotranscriptional and posttranscriptional processes. 23 For example, antisense RNA Interferon Regulatory Factor 1 Antisense RNA (IRF1-AS) could form a complex with interleukin enhancer binding factor 3 (ILF3) and DHX9 proteins at the IRF1 promoter region to activate IRF1 transcription. 24 Notably, DHX9 was also identified as lncRNA PTCSC3 binding protein in our study. In addition, antisense lncRNA IGFBP7-AS1 forms an RNA-RNA double-strand complex with its adjacent gene IGFBP7 to enhance IGFBP7 RNA stability. 25 Some antisense lncRNAs further alter mRNA alternative splicing by reducing U2 snRNP recruitment to the target pre-mRNAs. 26 Here, unlike the previously reported models, lncRNAs PTCSC3 and PTCSC3-AS1 synergistically function as dual activators to enhance RNA stability and inhibit cancer development. Our findings elucidate a dual-lncRNA regulatory mechanism underlying PTC tumorigenesis, providing a framework for comprehensively understanding noncoding RNA interplay in cancer biology.
In our study, the canonical epithelial-mesenchymal transition (EMT) marker vimentin was identified as a newly discovered suppressor of ZC3H12A in thyroid cancer cells. Similar to that, some studies have also demonstrated the molecular regulatory function of vimentin, which is involved in direct protein–protein or protein–RNA interactions. It has been reported that vimentin can downregulate osteocalcin expression by suppressing the transcriptional activity of activating transcription factor 4 (ATF4), resulting in the inhibition effect of cell differentiation in osteoblasts. 27 In addition, vimentin has been shown to bind to the 5′UTR of collagen mRNAs in a La ribonucleoprotein 6, translational regulator (LARP6)-dependent manner and regulate tissue fibrosis development by stabilizing collagen mRNAs. 28 Furthermore, vimentin has been found to stabilize TF mRNA and interfere with miRNA-dependent regulation at 3′UTR during early metastasis of vimentin-expressing circulating tumor cells. 29 In comparison to previous findings, our study reveals that vimentin can act directly as a destabilizing factor at ZC3H12A 3′UTR. Further efforts were needed to delineate its precise mechanistic contributions to PTC tumorigenesis in the future.
In addition to the binding capacity with coding mRNAs, vimentin has also been shown to play a regulatory role by directly binding to noncoding RNAs. For instance, the vimentin-binding lncRNA VAL has been found to competitively inhibit Trim16-dependent vimentin polyubiquitination and degradation in lung adenocarcinoma. 30 Here, we identified that lncRNA PTCSC3 and its antisense RNA PTCSC3-AS1 directly bind to vimentin and competitively impact ZC3H12A mRNA degradation. It provides new evidence supporting the regulatory role of vimentin at the posttranscriptional level in carcinogenesis.
Previous studies have shown that ZC3H12A participates in regulating genes involved in oncogenic growth signaling networks. In renal cell carcinoma, ZC3H12A has been found to affect the inflammation process by degrading mRNAs of pro-inflammatory cytokines, such as IL6, IL1, and IL12. 31 Mechanistically, ZC3H12A mainly participates in the RNA degradation process by recognizing stem-loop structures presented in 3′UTR. It also acts as a suppressor of miRNA biogenesis by cleaving the terminal loops of precursor miRNAs. 32 Notably, a few studies have revealed that lncRNAs take part in ZC3H12A regulation in cancer development. For example, the upregulated Linc00936 inhibited tumor immune escape and aggressiveness of gastric cancer cells by elevating ZC3H12A, while the molecular mechanism remains unclear. 25 Here, our results show how lncRNAs participate in regulating ZC3H12A in tumorigenesis and how they interact with shared protein partners to perform regulatory functions. Future experiments, such as chromosome conformation capture (3C) assays, are needed to further elucidate the physical interactions between regulatory variants and candidate target genes at the 14q13.3 locus. In addition, visualization techniques, such as proximity ligation assay, could be attempted in optimized thyroid primary cells or tissue samples to directly observe the interactions between the two lncRNAs and vimentin in future studies.
Conclusions
In summary, our study describes a dual-lncRNA regulatory axis (PTCSC3-AS1/PTCSC3) at the 14q13.3 locus that stabilizes ZC3H12A by antagonizing vimentin-mediated mRNA degradation. Functional variants (rs944289/rs116909374) potentially regulate this axis, establishing a mechanistic connection between 14q13.3 and PTC predisposition through RNA stabilization pathways.
Footnotes
Acknowledgments
The authors thank Bo Yu and Weichao Li for their assistance in clinical sample collection.
Authors’ Contributions
Y.W. contributed to the conception and design of this study. Y.Z., X.S., S.C., J.S., Z.A., and J.Y. conducted the experiments. J.L. and M.G. contributed to clinical sample collection and administration. B.Z. contributed to the data analysis of RNA-seq data. Y.Z. wrote the article draft. Y.W. and X.C. revised the article. All authors read and approved the final article.
Author Disclosure Statement
The authors declare that no competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (82072955), the Research Project Supported by Shanxi Scholarship Council of China (2024-074), Shanxi Province Higher Education “Billion Project” Science and Technology Guidance Project (BYJL066), the Scientific Research Startup Fund from Shanxi Medical University to Y.W., and the Doctoral Startup Grant from Shanxi Province in China (SD2033).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.