
Abstract
Background:
Mice lacking the thyroid hormone (TH) transporters monocarboxylate transporter 8 (Mct8) and organic anion transporter 1c1 (Oatp1c1) exhibit a strongly perturbed brain maturation, thereby replicating symptoms of patients with MCT8 deficiency. Mct8/Oatp1c1 double knockout (DKO) mice show a strongly diminished TH brain content, indicating a compromised TH passage across blood–brain barrier (BBB) endothelial cells that may represent a major pathogenic event causing CNS abnormalities. Here, we tested this hypothesis by generating mice that lack Mct8 and Oatp1c1 in endothelial cells only.
Methods:
Adult conditional Mct8/Oatp1c1 mice expressing a constitutively active Tek-driven Cre recombinase (Endo del mice) and control littermates were characterized regarding their hypothalamus–pituitary–thyroid axis, brain morphology as well as peripheral and central TH signaling. For comparison, age-matched DKO mice were included. Immunofluorescence (IF) studies were conducted to examine neural maturation. Fluorescence in situ hybridization (FISH) experiments and qPCR analysis were performed to determine transcript levels of TH-regulated genes in different brain regions. TH tissue content in dissected brain areas was quantified by LC-MS/MS analysis.
Results:
Analysis of different brain parameters by IF staining revealed a compromised maturation of cortical GABAergic interneurons and hypomyelination in Endo del mice, although the observed alterations were less profound than in DKO mice. TH content determination, FISH, and qPCR studies disclosed significantly reduced TH concentrations and, consequently, decreased TH signaling in several brain areas. Surprisingly, hippocampal T3 content and transcript levels of TH-regulated genes were found to be only mildly altered in Endo del mice compared with DKO animals.
Conclusions:
Deficiency of Mct8 and Oatp1c1 in endothelial cells results in reduced murine brain TH content and TH action, thereby underscoring the major function of BBB endothelial Mct8/Oatp1c1 in facilitating TH uptake into the CNS. Yet, the degree of central TH deficiency and neural impairments in Endo del mice are not as profound as in DKO mice. Particularly, unaltered hippocampal T3 signaling suggests a sufficient T3 supply of this brain area, possibly via the cerebrospinal fluid (CSF). These findings infer that apart from the BBB, additional Mct8/Oatp1c1-facilitated TH transmembrane passages (for instance, TH transport across the blood–CSF barrier) are required for proper mouse brain development and function.
Introduction
The monocarboxylate transporter 8 (MCT8) represents a highly specific thyroid hormone (TH) transporter facilitating the passage of both T4 and T3 across cell membranes. 1 Patients with inactivating mutations in the X-linked SLC16A2 gene encoding MCT8 display severe neurocognitive and movement impairments (Allan–Herndon–Dudley syndrome, AHDS). 2 –4 Apart from neurological symptoms, affected patients present abnormal serum TH parameters with highly elevated T3 that in turn causes a thyrotoxic state in liver, kidney, heart, and skeletal muscle. 5 In contrast, hypomyelination and an impeded neuronal differentiation, both also persistently seen in hypothyroid mouse models, are indicative for a profound TH-deficient state in the patients’ brain. 6,7 As MCT8 is expressed in human blood–brain barrier (BBB) endothelial cells and in blood–cerebrospinal fluid barrier (BCSFB) choroid plexus epithelial cells, a diminished TH transport across these brain barrier cells and, consequently, an insufficient TH supply during brain development have been suggested to be pivotal pathogenic mechanisms underlying the neurological symptoms. 8,9
Mice with global Mct8 deficiency (Mct8 KO mice) replicate the abnormal serum TH profile but exhibit only a mild CNS phenotype. 10,11 This finding can be explained by the presence of the T4-transporting organic anion transporter 1c1 (Oatp1c1) in murine brain barrier endothelial and choroid plexus cells where Oatp1c1 (encoded by Slco1c1) can partially compensate for the absence of Mct8. 12 –14 Thus, only Mct8/Oatp1c1 double knockout (DKO) mice show a strongly diminished TH transport into the CNS, leading to highly decreased TH brain concentrations and histomorphological alterations in neural development similar to those seen in MCT8-deficient patients. 15 Yet, which Mct8/Oatp1c1-dependent cellular TH passage is most critical for TH entering the CNS has still not been addressed.
Here, we tested the hypothesis that Mct8 and Oatp1c1 in BBB endothelial cells are primarily responsible for providing the CNS with TH. To this end, we crossed conditional Mct8/Oatp1c1 mutant animals with mice expressing a constitutively active Tek-driven Cre recombinase in order to inactivate both TH transporters in endothelial cells only. These so-called Endo del mice indeed exhibit reduced brain TH levels and TH signaling in distinct (albeit not all) brain areas but display a less disturbed neural maturation compared with global DKO mice. Altogether, our findings not only confirm a pivotal function of Mct8/Oatp1c1 in facilitating TH passage across BBB endothelial cells but also suggest a relevant contribution of Mct8/Oatp1c1 in the BCSFB and/or other CNS cell types to central TH homeostasis.
Material and Methods
All animal studies were executed in compliance with local regulations by the Animal Welfare Committee of the Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen (Recklinghausen, Germany; approval code AZ81-02.04.2022.A156). Mct8 fl/fl (or Mct8 fl/y) and Oatp1c1 fl/fl mice were generated and interbred as described elsewhere. 16,17 Mct8/Oatp1c1 double floxed (dfl) mice were mated with transgenic animals expressing a constitutively active Cre-recombinase under the direction of the receptor tyrosine kinase Tek promoter/enhancer (Cg-Tg(Tek-cre)1Ywa; JAX stock #008863) 18 in order to delete the loxP site flanked (floxed) exons 3 of the Mct8 encoding Slc16a2 gene and the Oatp1c1 encoding Slco1c1 gene in endothelial cells (Endo del mice). For monitoring Cre-recombinase activity, mice carrying an EYFP reporter construct were crossed in as described before. 19,20 Notably, the Tek-Cre transgene was transmitted only through the paternal lineage, and experimental animals (all on C57BL/6 background) were monitored for any undesirable germ-line deletion of the floxed alleles by PCR. As controls, Mct8/Oatp1c1 dfl littermates without a Tek-Cre transgene were used. For comparison, Mct8/Oatp1c1 DKO mice of similar age and sex were included as well.
For immunofluorescence (IF) studies, 4-month-old mice were subjected to intracardial perfusion-fixation with 4% paraformaldehyde in PBS (4% PFA) under deep ketamine/xylazine-induced anesthesia. Isolated tissues were post-fixed in 4% PFA for 24 hours. IF analysis was conducted using 50-µm-thick brain vibratome sections or 20-µm-thick cryosections from liver and kidney as described in Supplementary Data. For fluorescence in situ hybridization (FISH) and qPCR analyses and determination of serum parameters, 2–4-month-old male mice were killed by cervical dislocation. Dissected cerebral cortex, hippocampus, striatum, and hypothalamus for qPCR were immediately frozen on dry ice, while isolated brains for FISH were frozen in isopentane on dry ice. qPCR was conducted as described in Supplementary Data. Transcript levels were normalized to expression levels of Cyclophilin D as house-keeping gene and relative gene expression was calculated with the delta–delta Ct method. Control values were set as 1.0. FISH was performed as described in Supplementary Data to assess transcript levels of TH-target genes on 20-µm-thick coronal brain cryosections. Serum samples were used to quantify TSH concentrations by MILLIPLEX MAP Mouse Pituitary Magnetic Bead Panel (MPTMAG-49K; Merck) on a Luminex system (Millipore) according to the manufacturer’s instruction. Circulating T4, T3, and rT3 levels were quantified in serum samples by LC-MS/MS as described before. 21 For TH tissue content determination by LC-MS/MS, cerebral cortex, striatum, and hippocampus were dissected from 2–4-month-old female mice. The clean-up procedure for TH tissue extraction was performed as previously published. 22 For TH detection and quantification, a Sciex QTrap 5500 mass spectrometer was interfaced with Agilent 1290 Infinity II LC system. Elution and MS/MS parameters are reported in Supplementary Data. Brain capillaries were isolated from fresh-frozen cortices of 2–3-month-old male mice following a published protocol 23 and used for qPCR analysis as further detailed in Supplementary Data.
All data are represented as mean ± standard deviation. Statistical significance between control, Endo del, and DKO mice was determined by one-way analysis of variance followed by a Tukey’s multiple comparison test. Differences were considered significant when p < 0.05 and marked as follows: *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Results
Global absence of Mct8 and Oatp1c1 in DKO mice results in a TH-deficient CNS due to an abolished TH transport across brain barriers. 15 To explore the cell-specific function of both TH transporters in BBB endothelial cells, we generated conditional Mct8/Oatp1c1 dfl mice that additionally expressed a constitutively active Tek-driven Cre recombinase. The phenotype of these so-called Endo del mice was compared with their Cre-negative littermates that served as controls (Fig. 1A). By taking advantage of an EYFP reporter knock-in, Cre-recombinase activity could be detected in around 97% of all brain vessels (Supplementary Fig. S1). To validate Cre-mediated deletion of the loxP flanked exons 3 in the Slc16a2 and Slco1c1 genes, brain vessel preparations containing an enriched fraction of endothelial cells were obtained from adult male control, Endo del, and DKO animals, and isolated mRNA was subjected to exon 3-specific qPCR analyses. In Endo del mice, Mct8 (Slc16a2) mRNA was strongly reduced to approximately 20% of the respective control values, whereas Oatp1c1 (Slco1c1) mRNA levels were even close to the detection limit (Fig. 1B). These findings point to an efficient endothelial transporter elimination in Endo del mice, while the residual capillary Mct8 mRNA levels may reflect an undisturbed Mct8 expression in pericytes, smooth muscle cells, and/or other vessel-attached cells. To further confirm an endothelial-specific Mct8 inactivation, vibratome sections of perfusion-fixed adult mice were coimmunostained with antibodies against Mct8 and the endothelial cell marker CD31 (Fig. 1C; Supplementary Figs. S2 and Figs. S3). Only in control but not in Endo del mice, Mct8-specific immunostaining was visible in CD31-positive endothelial cells. To exclude any off-target inactivation in other brain barrier cell types, Mct8 protein expression was also examined in choroid plexus epithelial cells comprising the BCSFB as well as in tanycytes of the third ventricle where Mct8 may be involved in hypothalamic TH supply. 14,24 –26 In both cell types, a normal Mct8 expression pattern was sustained in Endo del mice, indicating that Mct8-mediated TH transport in tanycytes and choroid plexus is not impeded (Fig. 1C).
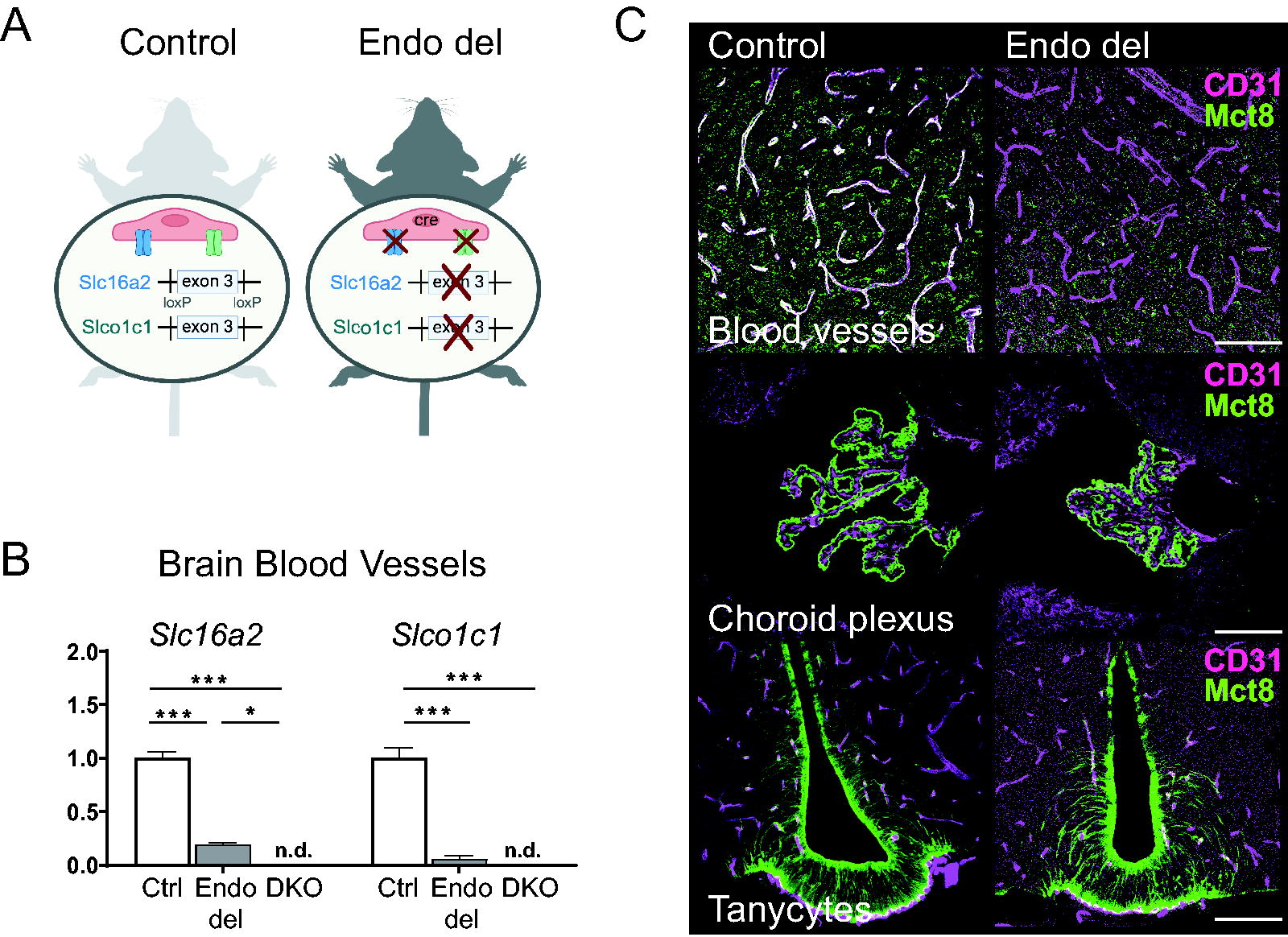
Cell-specific inactivation of Mct8/Oatp1c1 in murine brain endothelial cells. Schematic illustration of genomic manipulation in Endo del and control mice
As Mct8 is also strongly expressed in liver and kidneys, we compared Mct8 protein localization in control and Endo del mice but did not detect any obvious alterations (Fig. 2A, B). We also failed to detect Mct8 in hepatic or renal CD31-positive cells in control mice, indicating that Mct8 is not involved in endothelial TH transport in these tissues. Since in global Mct8-deficient and DKO mice, hepatic and renal deiodinase type 1 (Dio1) expression is highly upregulated, 15 we also quantified Dio1 transcript levels by qPCR but again failed to observe any significant differences between control and Endo del mice (Fig. 2C, D). These findings point to a rather normal TH status in liver and kidneys of Endo del animals.
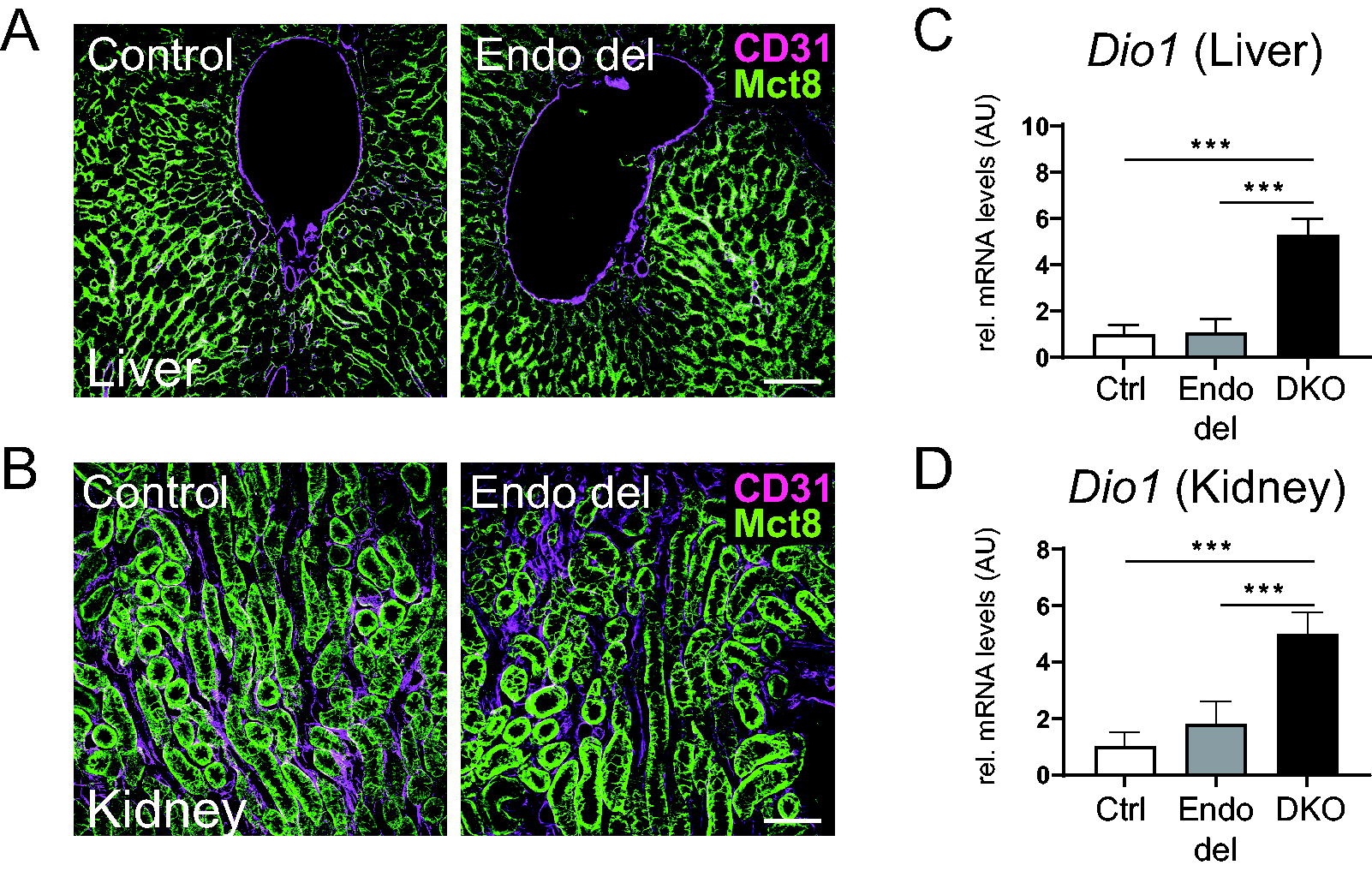
Analysis of Mct8 and Dio1 expression in liver and kidney of Endo del mice. In liver and kidney sections of control mice, immunofluorescence staining did not reveal an overlapping expression pattern for Mct8 (green) and the capillary marker CD31 (violet). Likewise, hepatic and renal Mct8 protein expression was not visibly altered in Endo del mice
A hallmark of human and murine Mct8 deficiency is a highly disturbed hypothalamus–pituitary–thyroid (HPT) axis with abnormal serum TH parameters. 10,11,15,27 We therefore studied hypothalamic Trh mRNA expression in the hypothalamic paraventricular nucleus (PVN) by FISH and found strongly elevated Trh levels in Endo del mice, almost reaching those seen in DKO animals (Fig. 3A, B). Further, quantification of type 2 deiodinase Dio2 mRNA expression in dissected hypothalami by qPCR revealed increased levels in Endo del and DKO mice (Fig. 3C), pointing to a hypothyroid state in the hypothalamus. In the pituitary, qPCR analysis disclosed increased Tshb transcripts in Endo del mice and DKO mice (Fig. 3D). Measurement of serum TSH levels, however, revealed normal values in Endo del mice (Fig. 3E). In the thyroid, Mct8 is present in thyrocytes but not in CD31-positive vessels. Consequently, thyroidal Mct8 expression was visibly not altered in Endo del mice (Fig. 3F). Likewise, determination of serum TH values revealed only a small increase in T4, T3, and rT3 concentrations, whereas DKO mice exhibited the expected abnormal pattern with elevated T3 and decreased T4 and rT3 levels (Fig. 3G). In sum, in contrast to DKO mice, Endo del mice exhibit only mild alterations in serum TSH and TH parameters.
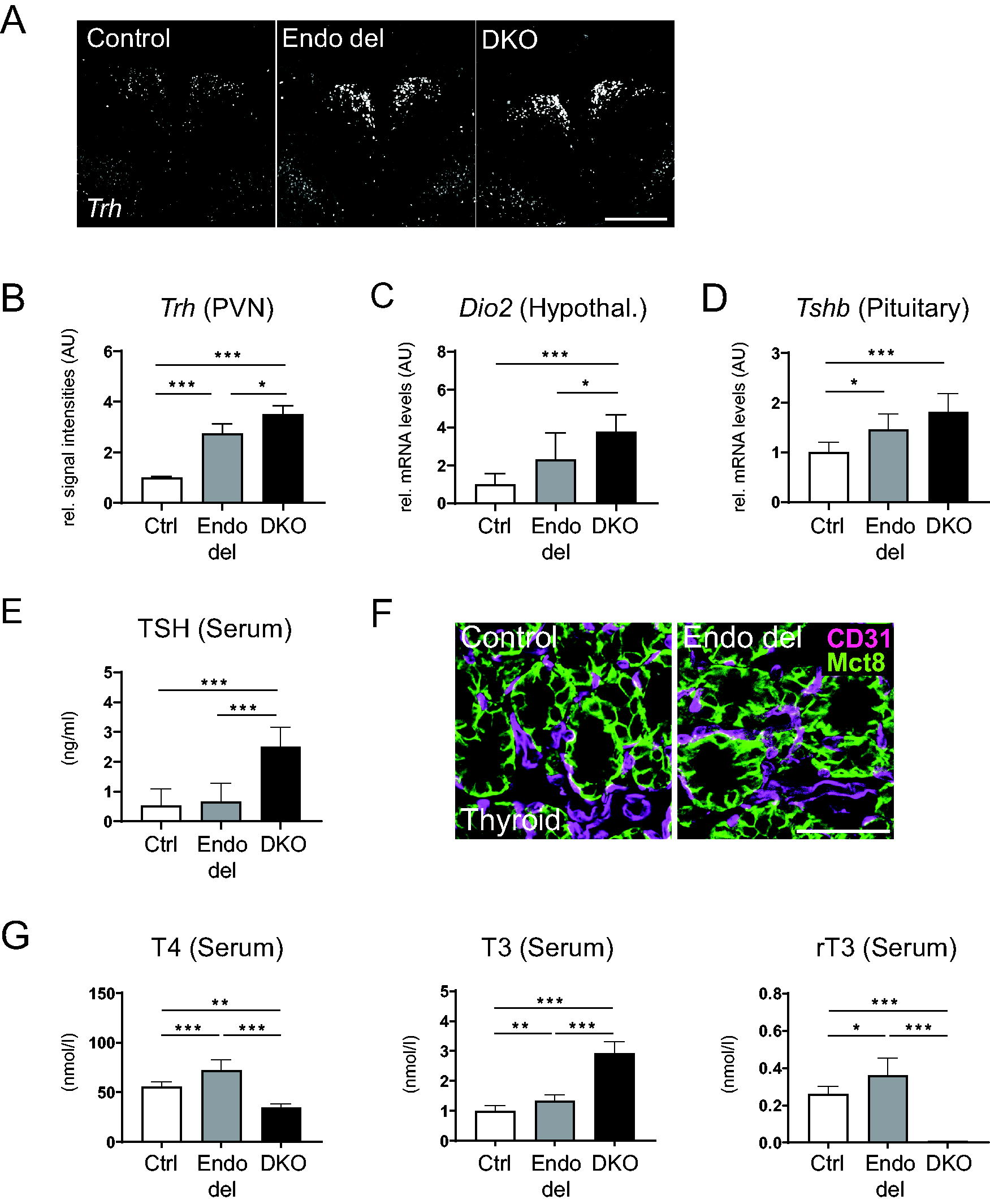
Activity of the hypothalamus–pituitary–thyroid axis in Endo del mice. Trh mRNA expression in the PVN was visualized in male control, Endo del, and DKO mice by FISH
We further analyzed several histomorphological parameters known to be altered in the CNS of AHDS mouse models. 15,28,29 A key hallmark represents a compromised cortical GABAergic interneuron maturation in the somatosensory cortex as evidenced by a reduced number of Parvalbumin immunopositive neurons, a rise in Calretinin expressing neurons, and a reduced immunoreactivity of the GABA-producing enzyme GAD67 in DKO mice 15,30 (Fig. 4A). Similar (albeit less profound) changes could also be detected in Endo del mice (Fig. 4A). We further examined myelination by quantifying cortical myelin basic protein immunoreactivity and fluoromyelin staining in the corpus callosum and detected a decrease in Endo del mice though less drastic compared with DKO animals 15,31,32 (Fig. 4B). Altogether, our IF studies confirm an altered cortical maturation and myelination in Endo del mice. Yet, the degree of impairment in Endo del mice is lower than in DKO animals.
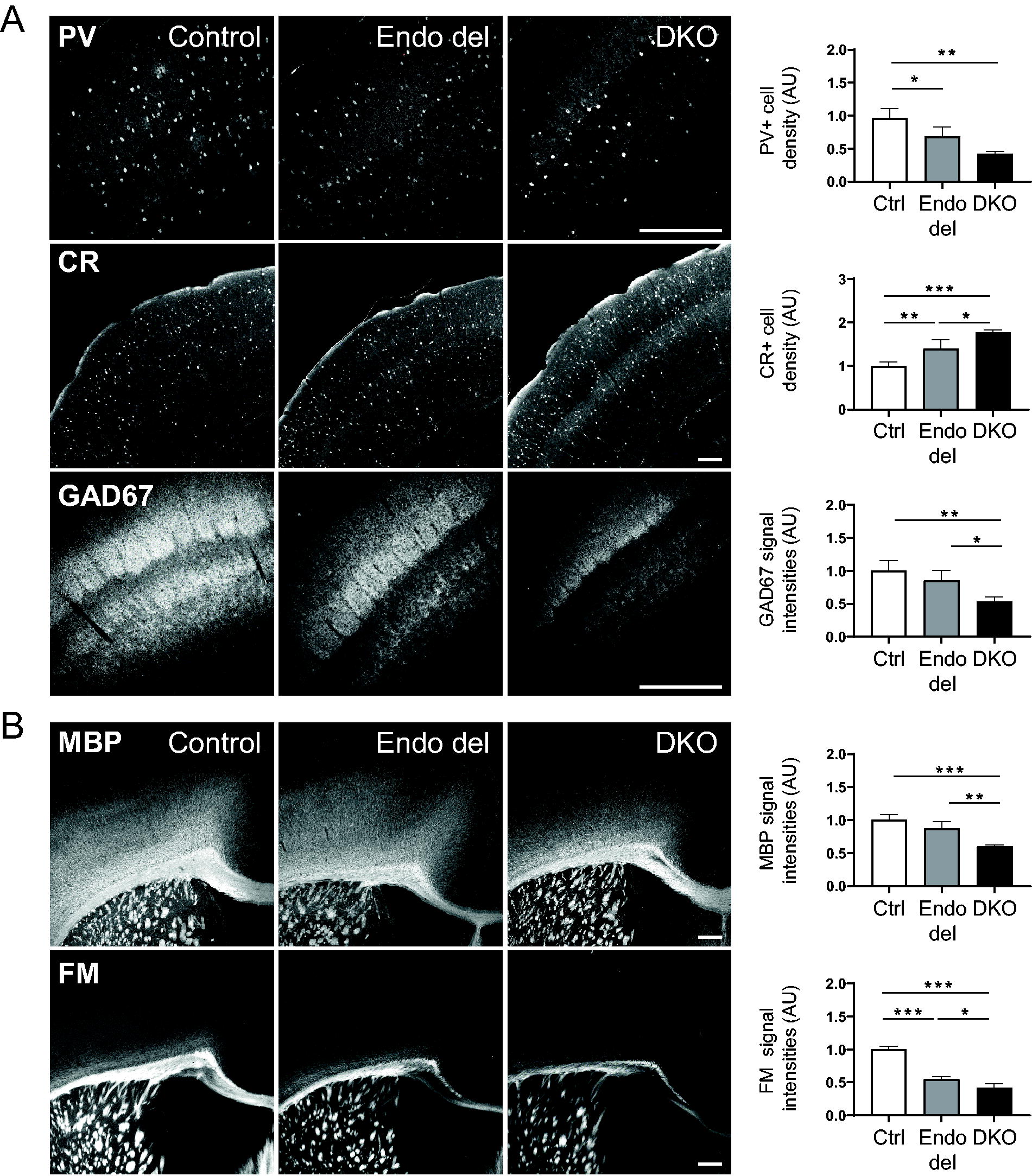
Analysis of cortical GABAergic interneuron maturation and myelination in Endo del mice. Immunofluorescence staining of coronal brain vibratome sections was conducted to study GABAergic interneuron marker expression in the somatosensory cortex of adult control, Endo del, and DKO mice (3 sections/animal and 4–5 males/genotype). Endo del mice showed a reduced number of PV immunopositive cells and an increased number of CR immunopositive cells, but changes were less prominent than in DKO animals. Likewise, a significant reduction in GAD67 immunoreactivity was only observed in DKO mice
We speculated that distinct brain areas might be more dependent on endothelial Mct8/Oatp1c1-mediated TH transport than others and therefore collected cerebral cortices, hippocampi, and striatal tissues from control, Endo del, and DKO mice for tissue TH content determination by LC-MS/MS (Fig. 5A). In line with previous results, 27,33,34 DKO mice showed in all three brain regions strongly reduced T3 values, while T4 and rT3 concentrations were even below the detection limit. In Endo del mice, T3 concentrations were reduced to 45% of control values in striatum, 78% in hippocampus, and 37% in cortex, whereas T4 concentrations were even further decreased in all three brain areas (Fig. 5B). Of note, rT3 and T2 tissue concentrations could be quantified in all three brain areas of control animals, whereas both TH metabolites were consistently below the limit of detection in DKO mice and only occasionally detected in Endo del mice (Supplementary Table S1). Together, these data not only confirm a reduced TH content in the CNS of Endo del mice but also indicate a much less affected brain TH import in these animals compared with DKO mice.
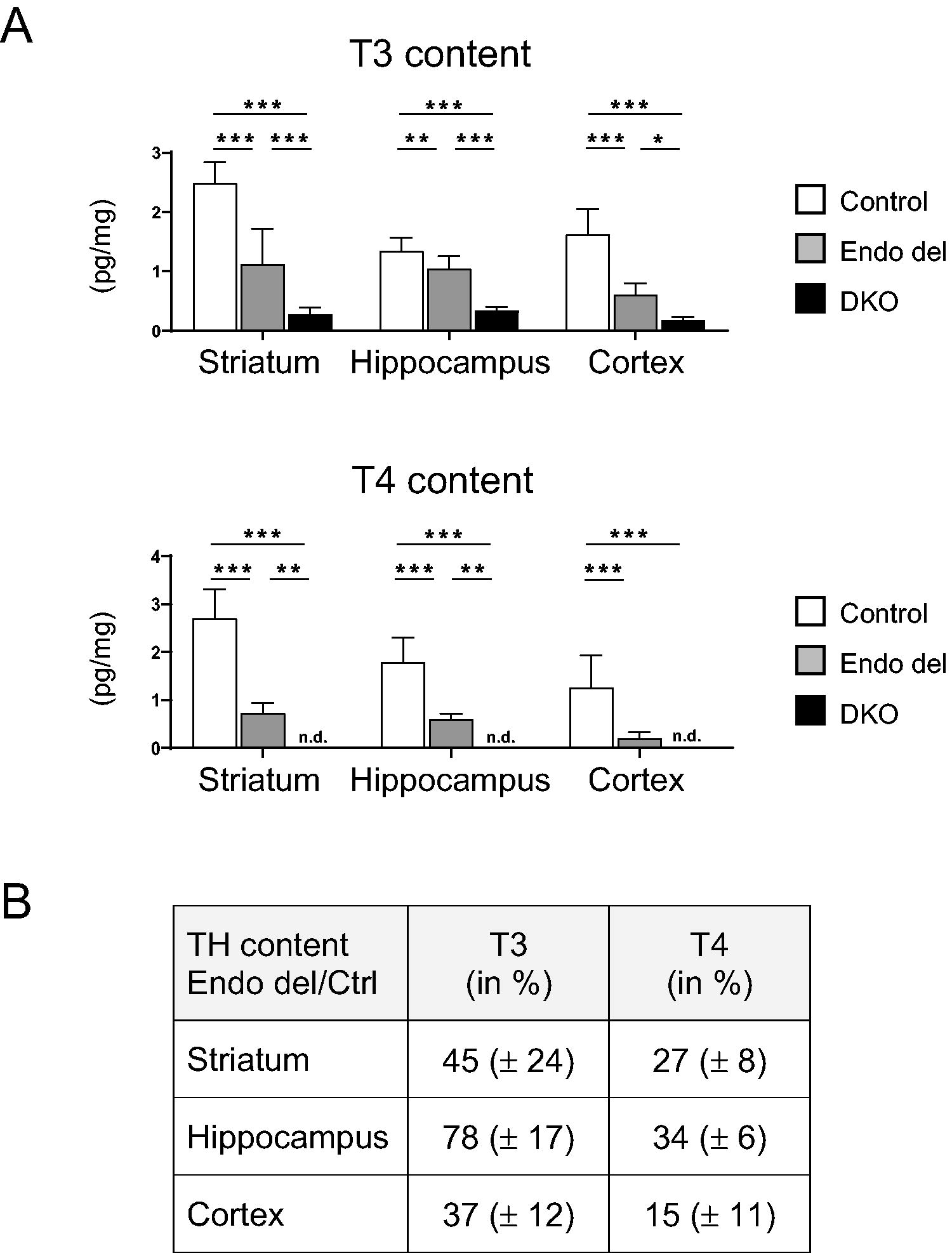
Determination of brain TH content in Endo del mice. Tissue T3 and T4 content was measured by LC-MS/MS in isolated striatum, hippocampus, and cerebral cortex samples of 2–4-month-old female control, Endo del, and DKO mice (8–9 females/genotype). Values are presented as pg/mg tissue. n.d., not detected
In order to assess the impact of altered TH content on T3 signaling in striatum, hippocampus, and cortex, we conducted qPCR analysis for Rc3 (Fig. 6A), Hairless (Hr) (Fig. 6B), and Klf9 (Fig. 6C) as well-established T3-regulated genes. 35 As expected, a stark drop in mRNA expression was found in DKO mice for all three genes. Endo del mice exhibited similarly reduced striatal Rc3, Hr, and Klf9 transcript levels and decreased cortical Rc3 and Hr mRNA expression. Intriguingly, hippocampal expression of all three TH-regulated genes in Endo del mice was found to be close to normal.
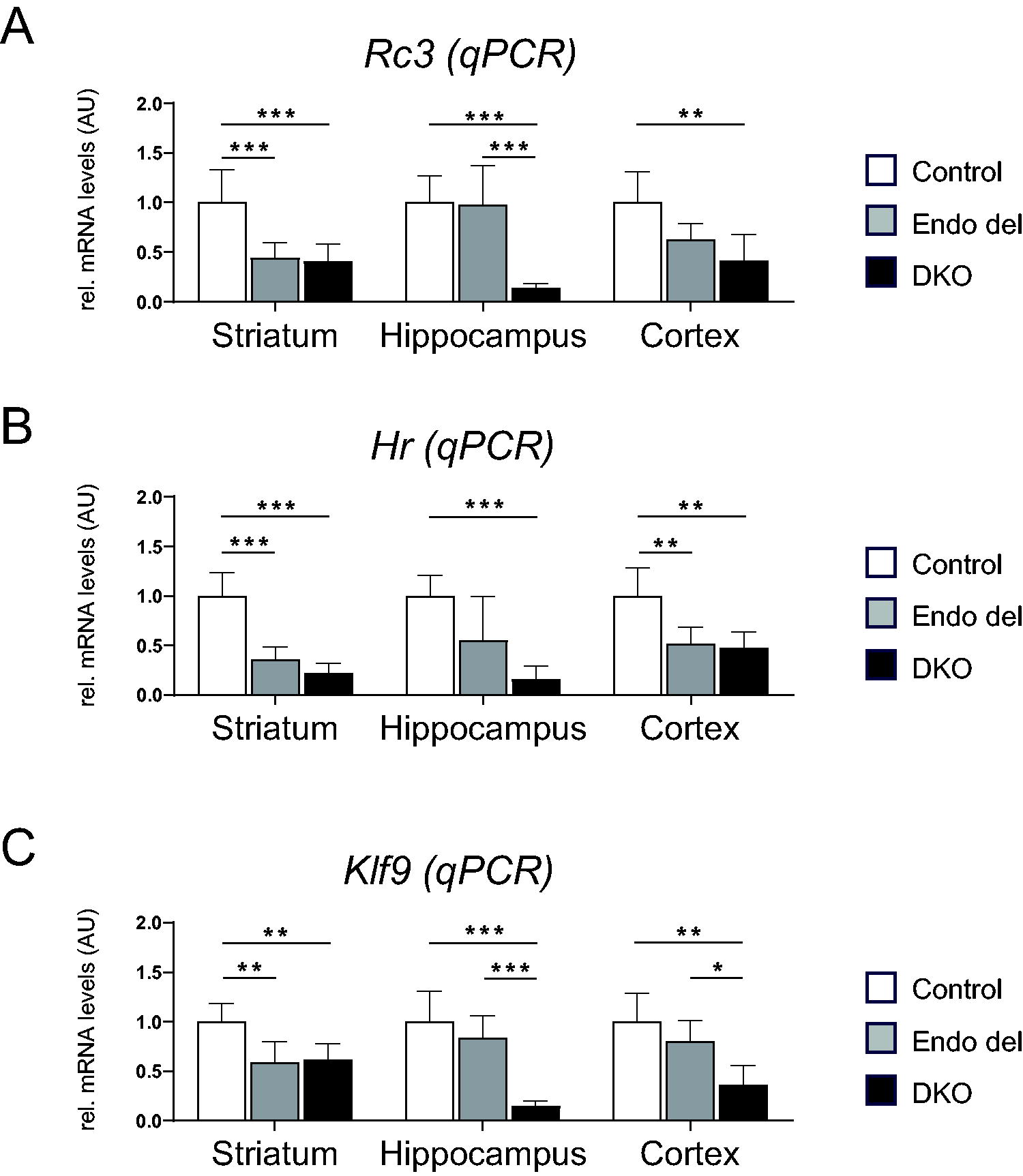
Analysis of T3-regulated target gene expression in dissected brain areas by qPCR. Striatum, hippocampus, and cerebral cortex samples were dissected from 2–4-month-old control, Endo del, and DKO mice (6–8 males/genotype) and subjected to RNA isolation. qPCR analysis was conducted to quantify mRNA levels of Rc3
To further assess mRNA changes of TH-responsive genes with spatial resolution, we conducted FISH experiments and quantified hybridization signal intensities in different brain areas (Fig. 7A). In agreement with our qPCR results, Rc3 signal intensities were decreased in cortex and striatum of Endo del and DKO mice (Fig. 7B). Klf9 signal intensities were also low in the cortex of Endo del animals but normal in the hippocampal CA1 area. As a gene negatively regulated by TH, we analyzed mu crystallin (Crym) mRNA expression and detected elevated cortical levels but normal expression in the hippocampal CA3 region of Endo del mice. Altogether, our studies indicate that Endo del mice display a reduced T3 action in the striatum and cortex but a rather normal T3 signaling in the hippocampal formation.
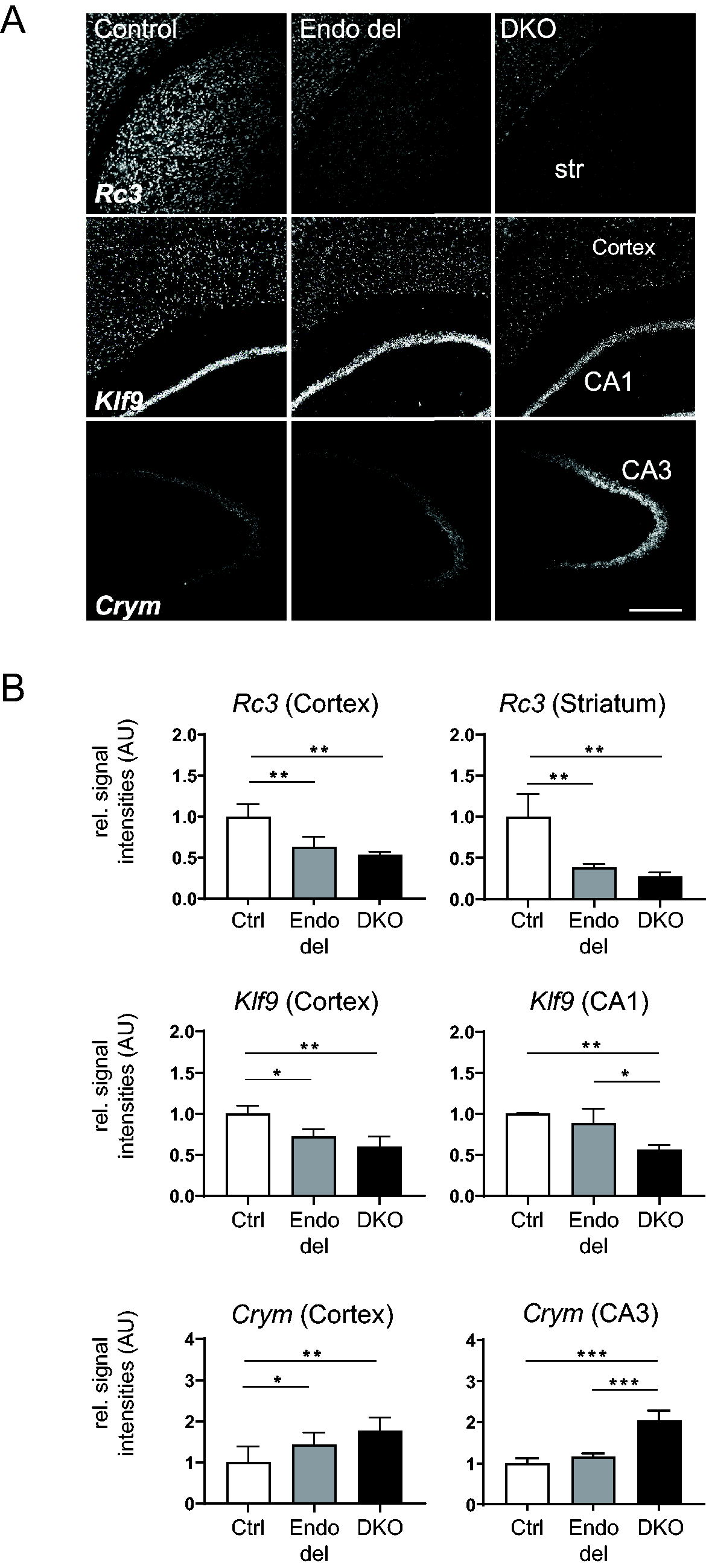
Assessment of T3-regulated gene expression in brain sections by FISH. FISH studies were conducted to visualize transcript levels of the T3-regulated targets Rc3, Klf9, and Crym in brain sections of control, Endo del, and DKO mice
Discussion
Despite having highly elevated serum T3 levels, DKO mice were repeatedly shown to exhibit a profound TH deprivation in the CNS that has been linked to a diminished TH transport into the brain. 15,22,27,33,34 As Mct8 and Oatp1c1 are strongly expressed in murine brain capillary endothelial cells, an impaired TH passage across the BBB was suggested to be the primary cause for the central TH deficiency of DKO mice. In order to corroborate this hypothesis, we generated Endo del mice by crossing Mct8/Oatp1c1 floxed mutants with Tek-driven Cre animals exhibiting a pan-endothelial cell Cre-recombinase activity. By this approach, Mct8/Oatp1c1 expression was efficiently abolished in brain endothelial cells but remained unaffected in choroid plexus epithelial cells or hypothalamic tanycytes, which represent alternative routes for TH to reach the CNS. Since both TH transporters are not present in endothelial cells outside the CNS, Mct8 localization in peripheral organs was preserved. Normal renal and hepatic Dio1 expression further denotes that TH metabolism in these tissues is not disturbed.
Notably, Endo del animals exhibited a small but significant rise in serum TH. This increase points to an altered set point of the HPT axis particularly as hypothalamic Trh expression was highly elevated. It is thus reasonable to assume that in response to a strong TRH stimulus, pituitary thyrotrophs produce TSH with a higher biological activity, which in turn leads to an enhanced thyroidal T4 and T3 secretion. Yet, in contrast to DKO mice, serum TSH levels in Endo del mice are in the normal range, implying that at least on the pituitary level, the negative feedback mechanisms are largely intact in these mutants.
The strong rise in hypothalamic Trh, however, indicates that circulating TH concentrations are not properly sensed by hypophysiotropic PVN neurons in Endo del mice. This observation is intriguing since tanycytes lining the third ventricle were suggested to control hypothalamic TH availability. As specialized ependymal cells, tanycytes indeed express TH transporters as well as Dio2 and their processes are in close contact with axonal endings of PVN neurons. 26 Yet, as TH transport via tanycytes is expected to be undisturbed in Endo del mice, one has to assume that hypophysiotropic PVN neurons are more dependent on TH traversing the BBB for proper feedback regulation.
A disturbed maturation of the cortical GABAergic system and hypomyelination are prominent alterations consistently described in AHDS mouse models. 6,28,29 Both maturation processes are clearly affected in Endo del mice as well, yet to a much lesser extent compared with DKO mice. This discrepancy underscores the relevance of additional Mct8/Oatp1c1-mediated TH transmembrane transport that is abolished in DKO animals but preserved in Endo del mice. Prominent examples are TH transport processes into oligodendrocytes 31,32 or into GABAergic progenitor cells. 30 Furthermore, the rather moderate CNS phenotype of Endo del mice suggests the presence of alternative routes by which TH can enter the CNS apart from the BBB.
The latter notion is supported by brain T4 content measurements. Endo del mice indeed exhibited a strong reduction with T4 values ranging from 15% in cortex to 34% in hippocampus of control values. Yet, in DKO mice, brain T4 concentrations are below the limit of detection. Obviously, limited amounts of T4 still reach the brain parenchyma of Endo del mice, possibly via the BCSFB. Compared with T4, brain T3 concentrations were even less drastically reduced in Endo del mice ranging from 37% to 78% of the respective controls. Possibly, a strong rise in brain Dio2 activities leads to an enhanced local T3 production in Endo del mice, whereas this compensatory mechanism is less effective in DKO mice due to the overall strongly reduced brain T4 concentrations.
Notably, highest T4 and T3 tissue concentrations were found in the hippocampus of Endo del mice. Likewise, qPCR studies and FISH analysis revealed close to normal hippocampal transcript levels of T3-regulated genes, indicating that hippocampal T3 signaling is almost retained in Endo del mice. It is thus tempting to speculate that due to its vicinity to the ventricles, the hippocampus might be supplied via the BCSFB with sufficient TH if its BBB passage is impeded. Such a scenario is also in agreement with autoradiographic studies by Dratman and colleagues who compared the CNS distribution of radiolabeled TH upon either i.v. or intrathecal injection. 36 In contrast to the rather uniform signal distribution pattern observed in i.v. injected rats, a predominant hippocampal distribution pattern was seen upon intrathecal injection. By which transport processes, however, CSF-derived TH can pass ventricle ependymal cells and enter the brain parenchyma is still an open question and a matter of future studies.
In sum, the CNS phenotype of Endo del mice with an overall decreased brain TH content and TH signaling substantiates the notion that Mct8/Oatp1c1 represents critical TH transport systems in murine brain endothelial cells and also suggests a physiologically relevant contribution of Mct8/Oatp1c1-facilitated TH transport at the BCSFB. Our findings are in line with recent preclinical studies aiming to restore functional MCT8 expression at brain barrier cells by adeno-associated virus (AAV)-mediated gene therapy approach. Of note, treatment of neonatal DKO mice with an AAV-BR1-MCT8 vector construct that selectively transduce BBB endothelial cells significantly improved CNS maturation and brain T3 content, although normal values could not be reached. 33 Possibly, one might have to consider to restore MCT8 expression in both BBB endothelial cells and BCSFB structures simultaneously to refine future treatment strategies for AHDS.
Footnotes
Acknowledgments
Authors’ Contributions
Conceptualization: B.M., M.S., D.F., S.M., and H.H.; investigation: A.A., B.M., M.D.A., A.C., and A.B.; data analysis: A.A., B.M., M.D.A., T.D.M., A.C., H.M.-F., A.B., S.M., and H.H.; resources: M.S., H.M.-F., T.D.M., A.B., D.F., S.M., and H.H.; writing—original draft: A.A., B.M., S.M., and H.H.; writing—review and editing: all authors; supervision: T.D.M., H.M.-F., A.B., S.M., and H.H.; funding acquisition: M.S., H.M.-F., T.D.M., A.B., D.F., S.M., and H.H.
Author Disclosure Statement
All authors declare that they have no interests, financial resources, or employment relationships that improperly influence or affect the integrity of the submission.
Funding Information
This study was supported by funding from the Deutsche Forschungsgemeinschaft (DFG; German Research Foundation; Project-ID 424957847 CRC/TR296; P01 (H.H.; M.S.); P02 (H.M.-F.); P05 (T.D.M.); P10 (D.F.); P19 (S.M.)).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.