
Abstract
To characterize the potential transmission sites of sleeping sickness in Kinshasa, two entomologic surveys were carried out during the dry and the rainy seasons in rural and periurban areas of Kinshasa in 2005. About 610 pyramidal traps were set up, and 897 Glossina fuscipes quanzensis were captured. Environmental and biologic factors were reported, and relationships between these factors were evaluated using logistic regression and multiple correspondence analysis. The biologic factors (the presence of tsetse flies, human blood meals, and teneral flies) were progressively accumulated at each capture site to permit the characterization of the sleeping sickness transmission risk. The dry season was found to be a more favorable period for the disease transmission than the rainy season. Moreover, the landscapes characterized by the presence of argillaceous soils, raised ground cover with forest residues and rivers, were identified as types of environments with greater risk of sleeping sickness transmission. Pig breeding appeared as an important factor increasing the disease transmission. If vector control is continuously performed along rivers segments at high risk, the transmission of sleeping sickness in rural and periurban areas of Kinshasa will considerably decrease.
Introduction
The area of Kinshasa, well known as HAT focus since the beginning of the 20th century, has been considered as an extinct focus after the independence of the country in 1960 (Mulumba et al. 1993). Due to sociopolitical instabilities and the alteration of health services following the independence, an increase in the number of HAT cases was observed in all districts of Kinshasa with a peak of 912 new cases in 1999 (Bilengue et al. 2001). Seventy-four percent of the patients detected in 1999 were from the active part of the population; mostly farmers and fishermen (Kande, unpublished data). To clarify the epidemiologic situation of HAT in Kinshasa, several studies were conducted. In a case control study, Robays et al. (2004) concluded that living in the endemic province of Bandundu and in the rural Kinshasa increased the risk of contacting HAT. On the basis of epidemiologic data, Ebeja et al. (2003) reported that most urban resident affected people were in the first stage of infection, suggesting that they were recently or probably locally infected. This hypothesis was reinforced by De Deken et al. (2005), who detected suitable habitats for tsetse flies in the urban, periurban, and rural areas of the city using remote sensed data. Further, Simo et al. (2006) identified Trypanosoma brucei gambiense in tsetse flies and reported a predominant man/fly contact in the periurban and rural areas of Kinshasa, confirming thus the local transmission of HAT in Kinshasa.
The present study attempts to determine the characteristics of the environment that could favor HAT transmission in Kinshasa. The approach aims at linking the biologic risk factors reported by Simo et al. (2006) and environmental parameters of the periurban and rural areas, and at determining the landscape associated with a higher risk for HAT transmission.
Material and Methods
Study zone
Kinshasa is the capital of the République Démocratique du Congo and lies at 4S16, 15E17. It is also the capital of the “Kinshasa Province,” with the urban area covering 4.5% of the 9.965 km2 of the whole province. Two main HAT endemic provinces surround the province of Kinshasa, the Bas-Congo in the south and the Bandundu in the north and in the east. One part of the city is bordered by the Congo River and lies on an alluvial plain up to the hills of the southern, western, and eastern sites. An important hydrographical network crosses the mountains sites. The soils are sandy, sandy-argillaceous, to argillaceous. In the periphery, the vegetation types include steppes, semideciduous and riverine forest islands, and wooded and grassy savannah. The mean annual rainfall varies between 1200 and 1500 mm. For annual temperature and relative humidity, the annual means are around 25°C and 80%, respectively. Kinshasa has an Atlantic littoral climate due to the southwestern cold air stream of the Benguela. For this reason, Kinshasa experiences low rainfall events, low temperature, a minimum sunshine, and a maximum cloud mass.
The population is distributed in 24 districts and has been estimated upon simulations between 4.5 and 6.5 millions inhabitants. The mean density in the suburbs can be estimated at around 500 inhabitants/km2 (Pain 1984). With such a population, the association of unemployment and food precariousness led many people to produce their own food in the periurban and rural areas of Kinshasa.
Entomologic surveys
During the dry and rainy seasons, 610 traps were set up for 4 consecutive days in six capture sites around rural and periurban areas of Kinshasa (Simo et al. 2006). Flies were collected twice a day. Each fly was identified and dissected, and the midgut was put on microscope slides for trypanosomes identification and then collected on Whatman® paper for further polymerase chain reaction (PCR) analysis. The detailed description of the trapping procedure and the identification blood meals and trypanosomes are reported in Simo et al. (2006).
Biologic risk factors
Most of the biologic risk factors defined by Laveissière et al. (1994) to calculate an index of the risk transmission were used. These factors include the presence of tsetse flies (quantified through the apparent density per trap and per day [ADT]), the teneral status of the flies (new hatched flies that never had a blood meal), the percentage of blood meals taken on human (detected by heteroduplex PCR based method [Njiokou et al. 2004] and reflecting the interface between human and flies). We did not look for physiologic age of females because of the low added value compared with technical constraint (Laveissière and Grébaut 2003); on the other hand, teneral flies were identified as an essential biologic risk factor for the transmission of HAT due to the fact that they can be easily infected during their first blood meal on an infected host. Salivary glands were not examined for subspecific identification of trypanosomes because of the low infection rate of Trypanosoma brucei gambiense in these organs. Nevertheless, this has been compensated further by PCR analysis of tsetse fly midguts, sufficient to reflect the circulation of the parasite in the vector population.
Evaluation of environmental factors
Each trap location was georeferenced with a global positioning system and environmental parameters such as the human activities and the characteristics of the environment (Table 1) around the trap in a radius of 50 m were recorded (in Excel file by a geographer) in a standardized way. Biologic risk factors identified from the flies at each trapping sites were then added to the database. The link between the biologic risk factors and environmental parameters was investigated using the logistic regression. Statistical analysis was performed using XLSTAT (ADDINSOFT®) and R free program (© R Foundation for Statistical Computing). The biologic risk factors were compared by seasons, with the trapping site as statistical unit. The capture point and the variables were discrete and dichotomized (presence or absence).
The levels of transmission risk
In each capture site, a gradient of transmission risk was established by cumulating progressively the biologic risk factors characterizing the transmission risk index (Laveissière et al. 1994, Laveissière and Grébaut 2003).
RISK0 for traps with no tsetse fly,
RISK1 for traps that caught at least one tsetse fly,
RISK2 for traps with tsetse flies including at least one teneral fly or a fly with human blood meal,
RISK3 for traps with tsetse flies, including both a teneral fly and a fly with human blood meal, and
RISK4 for traps having all the biologic factors above plus Trypanosoma brucei gambiense midgut infection.
The landscape at risk
A multiple correspondence analysis (MCA) was performed to explore the parameters and modalities associated with the different levels of risk and to determine the landscape at risk prevailing in the study zone. Using the ArcGis 9.1 ® ESRI program, a geographic information system, was built to visualize the distribution of the results. The projection system references were: Universal Transverse Mercator Zone 33 South (UTM); Central meridian, 15°E; Scale factor, 0.99960; unity, the meter; referential ellipsoid and datum, WGS84. The design of the map shown in this paper (Fig. 1) was finalized with Adobe Illustrator 11.0 ®.
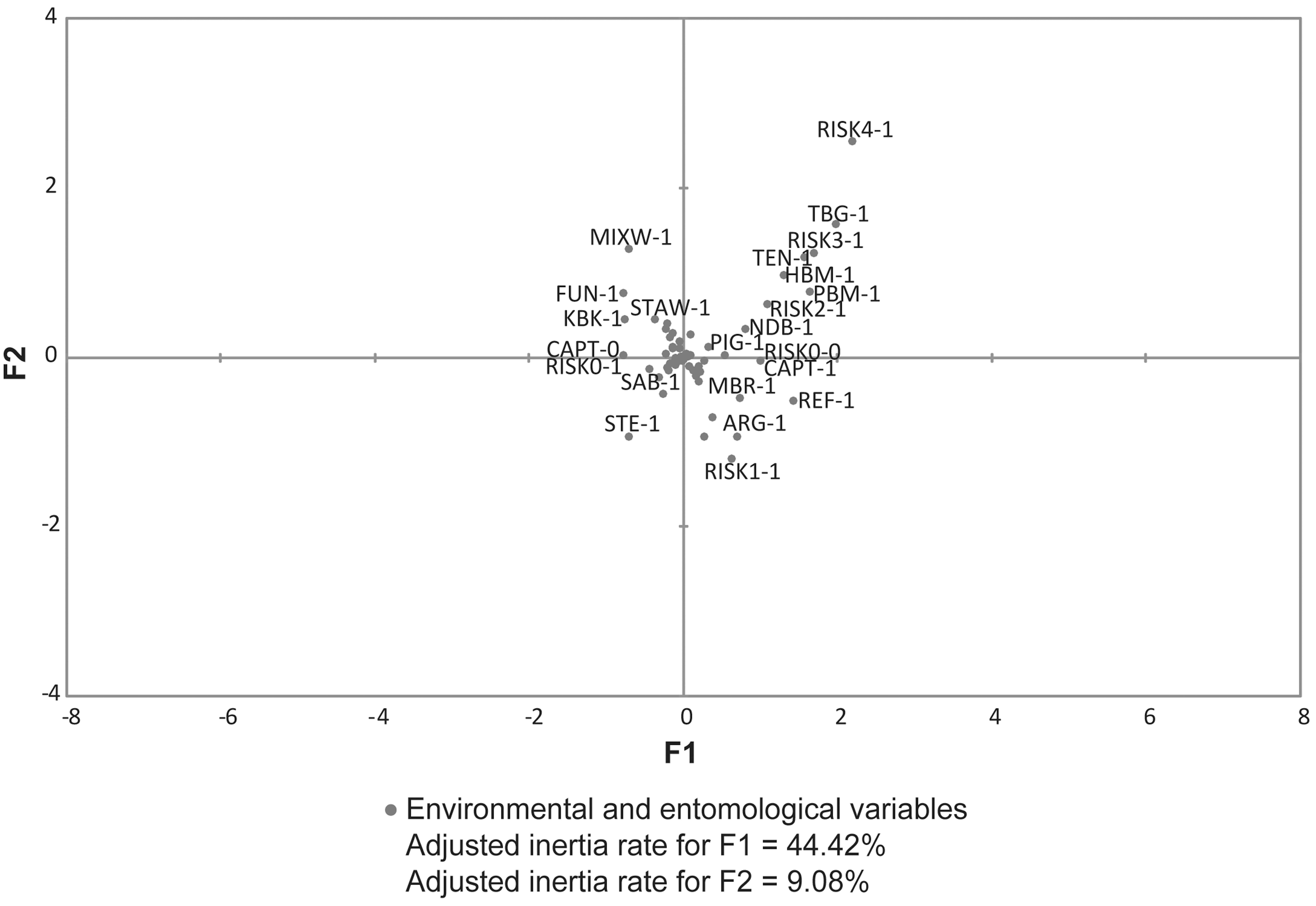
Multiple correspondence analysis graphical display of biologic and environmental factors of risks of sleeping sickness in the rural and periurban areas of Kinshasa. 1, presence; 0, absence; ARG, argillaceous soil; CAPT, tsetse flies captured; FUN, Funa site; HBM, human blood meal; KBK, Kimbanseke site; MBR, Mambre site; MIXW, mixed water (stagnant and flowing); NDB, Ndjili Brasserie site; PBM, pig blood meal; REF, forest residues; RISK, level of risk of transmission; SAB, sandy soil; STAGW, stagnant water; STE, steppe; TBG, T. b. gambiense in the midgut of tsetse fly; TEN, teneral flies.
Results
Entomologic surveys
During the two entomologic surveys, the 610 traps captured 897 Glossina fuscipes quanzensis (624 and 273 in the rainy and dry seasons, respectively). The annual mean ADT (0.37) is very low and stayed below 1 what ever the season (Table 2). Out of these, 570 tsetse flies (65.5%) were dissected: 376 and 194 in the rainy and dry seasons, respectively (Table 2). The detailed entomologic results for tsetse flies collected during the dry and rainy seasons are reported in Simo et al. (2006).
ADT, apparent density of flies per trap; No., number; TBG, Trypanosoma brucei gambiense.
Among the flies dissected, a relatively higher number of blood meals was obtained during the dry season campaign compared to the rainy season (64 versus 53; chi-square = 27.96, DF = 1, p<.000). The rate of human blood meals among the analyzed meals was comparable whatever the season (66% and 67% in the rainy and dry seasons, respectively).
Environmental risk factors
Several environmental parameters were used for statistical analysis (Table 1). Table 3 only reports the 11 parameters associated with a significant risk for HAT transmission. Tsetse flies are found mostly along rivers and in raised ground cover with forest residues and argillaceous soils. Human blood meals are found in flies captured in traps located in raised ground cover with forest residues and around pigsties. Herbaceous cover and stagnant water are unfavorable for teneral flies.
OR, odds ratio; CI, confidence interval.
p < 0.05.
p > .0.05 (not significant).
Unfavorable environmental unit for the presence of teneral flies.
Landscape exposing to HAT transmission risk
The five levels of risk of transmission defined above were associated with environmental parameters performing the MCA. The highest inertia (44.42%) was included in the F1 axis (Fig. 2). Taking into account the contribution of each variable, we tried to put out the variables that contributed the most to the F1 axis. The projections on this axe help to highlight a specific landscape with RISK0 that is observed in the periurban area of Kinshasa in Kimbanseke and Funa and is described as herbaceous cover (steppe and grassy savannah) lying on a sandy soil and in presence of stagnant water. On the opposite, the landscape associated with high biologic risk (RISK3 and 4) was projected on the other side of the F1 axis. This landscape was found in capture sites like Mambre and Ndjili Brasserie, and it was characterized by the presence of raised ground cover with forest residues, argillaceous soil, and the presence of pigsties.
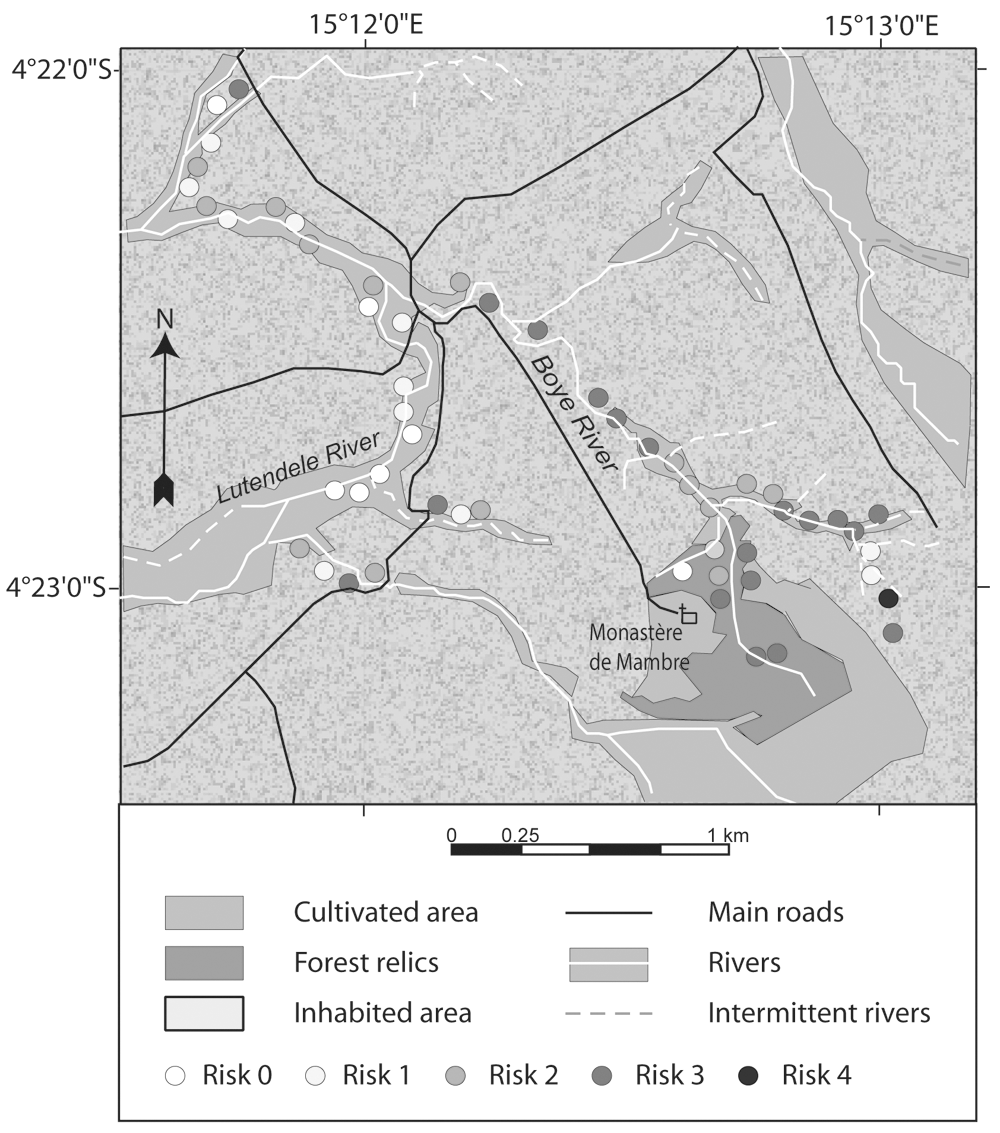
Example of capture site (Mambre) of rural and periurban areas of Kinshasa showing landscapes with the capture points and the gradient of transmission risk of HAT.
Discussion
This study attempted to identify and characterize potential transmission sites of HAT in rural and periurban areas of Kinshasa by putting together biologic and environmental data into a geographic information system in order to determine the most suitable environments where human populations are exposed to the transmission risks of HAT. It confirmed the very low density (ADP = 0.37) of Glossina fuscipes quanzensis in the periurban area of Kinshasa as reported by De Deken et al. (2005). This low fly population combined with the important man/fly contact (66.7% human blood meals) in Kinshasa can lead to high prevalence of HAT in human population. This is in line with the conclusions of Gouteux and Artzrouni (2000) who reported similar observations using biomathematics investigations to model the resurgence of HAT.
The argillaceous soils, the raised ground cover with forest residues, and the presence of rivers and pigsties are specific niches where biologic factors characterizing the transmission risk index of HAT were found during the dry and rainy seasons. This illustrates how the biologic factors are directly related to environmental units such as the type of soil and the vegetal cover. Indeed, the type of soil is directly related to tsetse flies larvae, which need a very soft ground to be buried. To enable larvae and pupae to complete their development before releasing teneral flies, local environmental factors such as the ground temperature that must not be neither too high nor too low, and the humidity neither too dry nor too humid (Schwetz 1919). All these environmental conditions partly depend of the vegetal cover that protects the soil from warming, drying, and eroding. In Kinshasa, argillaceous soils are generally associated with raised ground cover with forest residues (Flouriot et al. 1978) that provides environmental conditions where tsetse flies can find resting places, humidity, and branches to settle their larvae. In these types of environment, larvae find favorable places for their protection (humus and dead leaves cover), their development, and their maturation (dead wooden trunks) (Schwetz 1919, Laveissière 1977, Gouteux 1985). In the contrary, dense human encroachment, grassy landscapes, and areas with stagnant water appeared as unfavorable habitats for tsetse flies since no fly was captured in these types of environment. This can be explained by the lack of vegetal cover that can lead to mechanical and chemical degradations of the soil, thus providing thin elements and low percentage of clay found most often in the shallows (Van Caillie, 1989). In this context, the abiotic conditions, especially humidity and temperature, are unfavorable for tsetse flies.
Results of the MCA between environmental and biologic factors enable us to identify particular landscapes where the transmission risk is high (RISK3 and RISK4). Characterized by the presence of argillaceous soil and raised ground cover with forest residues, these landscapes appeared as zones where vector control operations should be prioritized to decrease the transmission of HAT in rural and periurban areas of Kinshasa. The capture points with low transmission risks are clustered in landscapes (same axis using MCA) characterized by herbaceous cover and sandy soil. Results of MCA confirm those obtained by cumulating, in the same capture point, most biologic factors characterizing the transmission risk index. They confirm also the usefulness of geographic information system as powerful tools for epidemiologic studies and especially to plan vector control operations as reported previously (Odiit et al. 2006, Bouyer et al. 2006, Symeonakis et al. 2007).
The dry season appeared as the most favorable period for HAT transmission because several environmental factors presenting a significant risk (Table 3) for the transmission of HAT are found during this season. Moreover, significant higher number of blood meals was taken during the dry season. These results can be explained by the fat reserves, which are fundamental for the survival of tsetse flies and play a major role in the water regulation. In tsetse flies, blood meals constitute the only way to supply these flies with water and energy (Bursell 1963). During the dry season, more water reserves in tsetse organisms are requested due to high temperature; more tsetse flies feed on mammals and, consequently, are more aggressive.
Whatever the season, the important contact frequency between man and tsetse flies (66% and 67% during the rainy and the dry seasons, respectively) can be explained by the absence of several alternative blood meals sources like wild animals for example. Indeed, the high pressure exerted by Kinshasa inhabitants on their environments did not enable wild fauna to survive in rural and periurban areas. The other types of breeding like goats and sheep are underrepresented in most capture sites. Compared to other HAT foci where tsetse flies can feed also on domestic and wild animals (Grebaut et al. 2004, Njiokou et al. 2004, Simo et al. 2008), only few alternative blood meal sources are available in Kinshasa. In such conditions, the aggressiveness of tsetse flies for human population becomes more important. The considerable percentage (27.3%) of pig blood meals identified in Kinshasa by Simo et al. (2006) suggests pigs as the second blood meal source for tsetse flies. The identification of one pig blood meal with Trypanosoma brucei gambiense infection (Simo et al. 2006) suggests that pigsties are an important risk factor for HAT transmission in Kinshasa. This must be put in perspective considering the significant number (odds ratio = 3.81; Confidence Interval 95% = 1.72–8.44; p = 0.002) of human blood meals found in tsetse flies captured in traps located close to pigsties. This important man–fly contacts around pigsties results from the particular way of pig breeding in Kinshasa: in most cases, owners confined their pigs in a room closed to their homes.
Our results provided additional elements enabling us to optimize vector control in Kinshasa and improved our knowledge on the bioecology of Glossina fuscipes quanzensis, a tsetse fly species that has been poorly studied. Complementarily to a previous study (De Deken et al. 2005) that showed the spatial distribution of Glossina fuscipes quanzensis in Kinshasa and proposed a map localizing suitable areas where these flies can be found, our study provided landscapes maps showing high transmission risk for HAT. The type of environmental patterns cumulating significant biologic risk was established.
Footnotes
Acknowledgments
We thank Mr. E. Elguero from IRD for his precious help during statistical analysis and Dr. C. Laveissière for critical reading of the manuscript.
Disclosure Statement
This study received financial support from the “Service de Coopération et d’Action Culturelle” of the French Embassy in RDC and “Institut de Recherche pour le Developpement (IRD, UR 177).”