
Abstract
Europe was officially declared free from malaria in 1975; nevertheless, this disease remains a potential problem related to the presence of former vectors, belonging to the Anopheles maculipennis complex. Autochthonous-introduced malaria cases, recently reported in European countries, together with the predicted climatic and environmental changes, have increased the concern of health authorities over the possible resurgence of this disease in the Mediterranean Basin. In Italy, to study the distribution and bionomics of indigenous anopheline populations and to assess environmental parameters that could influence their dynamics, an entomological study was carried out in 2005–2006 in an at-risk study area. This model area is represented by the geographical region named the Maremma, a Tyrrhenian costal plain in Central Italy, where malaria was hyperendemic up to the 1950s. Fortnightly, entomological surveys (April–October) were carried out in four selected sites with different ecological features. Morphological and molecular characterization, blood meal identification, and parity rate assessment of the anophelines were performed. In total, 8274 mosquitoes were collected, 7691 of which were anophelines. Six Anopheles species were recorded, the most abundant of which were Anopheles labranchiae and An. maculipennis s.s. An. labranchiae is predominant in the coastal plain, where it is present in scattered foci. However, this species exhibits a wider than expected range: in fact it has been recorded, for the first time, inland where An. maculipennis s.s. is the most abundant species. Both species fed on a wide range of animal hosts, also showing a marked aggressiveness on humans, when available. Our findings demonstrated the high receptivity of the Maremma area, where the former malaria vector, An. labranchiae, occurs at different densities related to the kind of environment, climatic parameters, and anthropic activities.
Introduction
In Italy malaria was eradicated between the end of the 1940s and the beginning of the 1950s (Soper et al. 1947, Canaperia and Patrissi 1948, Missiroli 1948), with the exception of two isolated foci of Plasmodium vivax that occurred in Sicily in the first half of the 1960s (Cefalù and Gullotta 1959, Lazzara et al. 1967). At the time, the main malaria vectors were Anopheles labranchiae Falleroni, 1926, and Anopheles sacharovi Favre, 1903, predominant along the coastal plains of central and southern regions. Nevertheless, other species belonging to the An. maculipennis complex contributed to maintain low levels of endemicity in some internal areas of the country, where the two main vectors were absent. After the 5-year national campaign against malaria, launched in 1947, anopheline vectors apparently disappeared or were dramatically reduced in abundance (Raffaele 1964). In some historical hyperendemic areas, located in the Tuscany region (Central Italy) and on the island of Sardinia, where the powerful vector An. labranchiae was very abundant, malaria control operations lasted 10 years more (Aitken and Casini 1966, Majori et al. 1970). In particular, during periodical surveys conducted in the Maremma, a coastal plain in the Tuscany region, this species was not found until the late 1960s, while in natural breeding sites other anopheline species were recorded (Coluzzi and Finizio 1966, Majori et al. 1970). Starting in the 1970s, despite the warning launched by the local health authorities, intensive rice cultivation was introduced in the Grosseto Province. In the following years, the regular surveillance for anopheline mosquitoes showed a constant rise of An. labranchiae presence, which in 1994 reached 100% of the whole anopheline sample collected in the ricefields areas (Bettini et al. 1978, Romi et al. 1992). After that, surveillance and control measures were implemented to reduce larval development in ricefields, and regular surveys were extended to other ricefields located inland (Siena Province), where remarkable densities of Anopheles atroparvus Van Thiel, 1927, (80%) and An. maculipennis s.s. Meigen, 1818, (20%) were recorded (Romi et al. 1992). An extensive survey on malaria vector status in Italy, carried out in the mid-1990s, identified the Maremma as a major at-risk area for malaria reintroduction in the country (Romi et al. 1997a). In 1997 an autochthonous-introduced malaria case, reasonably transmitted by indigenous An. labranchiae, occurred in the Grosseto Province (Baldari et al. 1998, Romi et al. 2001). As a consequence of this event, surveillance and control activities of anopheline foci were increased (Regione Toscana 1998).
To evaluate the present risk of malaria resurgence in this former hyperendemic area, where the potential vector is still present at epidemiologically relevant densities, a study on the bionomics of anophelines was implemented. The results of a 2-year longitudinal study, carried out with regular surveys, from 2005 to 2006, are shown and discussed, also by the light of the recent existing literature on potential malaria vectors in Europe (Zamburlini and Cagnus 1998, Merdić and Boca 2004, Bietolini et al. 2006, Patsoula et al. 2007, Ponçon et al. 2007).
Materials and Methods
Study area
The study area represents a large part of a physical region named the Maremma, a coastal plain of Central Italy that extends along the Tyrrhenian Sea, from South Tuscany to North Latium Regions and inland to the Apennine foothills (Fig. 1). The area is shared out among the administrative provinces of Grosseto, Siena, and Viterbo, with a population density of about 52 inhabitants/km2. It is characterized by a Mediterranean climate with a yearly average temperature of 14.5°C, ranging from a minimum of 7.1°C in January to a maximum of 23.1°C in August and a yearly average rainfall of 66.7 mm (from 16.9 mm in July to 99.9 mm in November). A large variety of biotopes (sandy coasts, dunes, Mediterranean bush, woodland, pasture, and cultivation) and of natural water bodies (marshes, lagoons, ponds, lakes, rivers, and streams) characterize this area. The major human activities are tourism (camping, hotels, and farm-holidays), intensive farming, and livestock breeding.
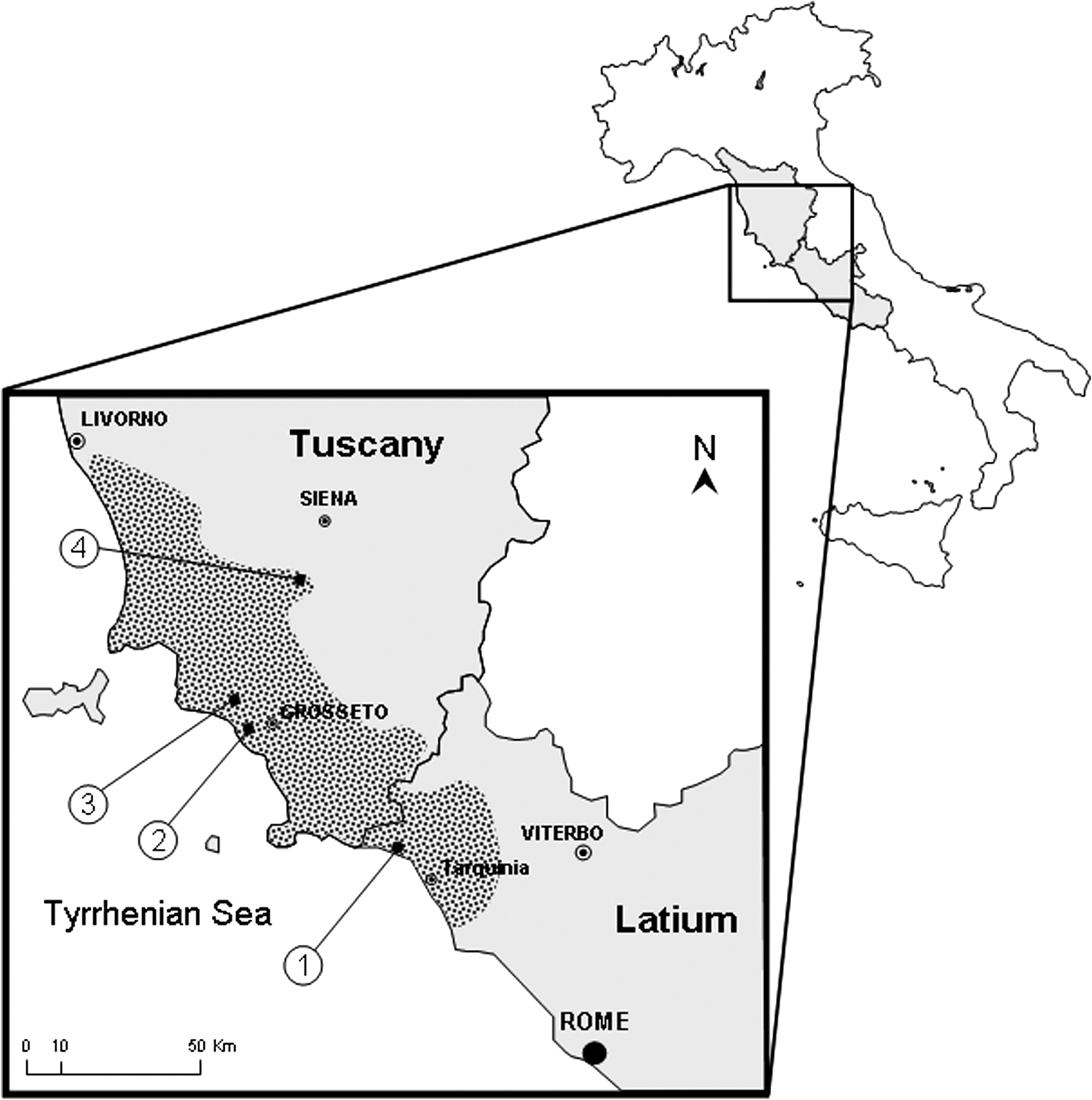
Location of study area in the Maremma (dotted), indicating the four mosquito collection sites.
Four sites, represented by farms, were selected for longitudinal surveillance based on their ecological features and location. As potential breeding sites we considered those located not more than 500 m from the farms (ricefields, irrigation canals, drainage canals, and a permanent stream):
Site 1—Valfragida (42°18′26″N and 11°39′27″E), located along the coastal plain (Viterbo Province) in the southern limit of the Maremma and characterized by cattle rearing (for milk and dairy products) and extensive tomato and wheat cultivation.
Site 2—Principina (42°43′29″N and 11°02′28″E) located near to the coastal part of the Grosseto Province, characterized by sheep, horse, and swine rearing and extensive ricefields (250 ha).
Site 3—Ampio (42°47′30″N and 10°56′51″E) situated in the interior of the Grosseto Province, characterized by extensive sunflower and maize cultivation and horse rearing. In 1997, near to this farm, the autochthonous-introduced malaria case occurred.
Site 4—Val di Merse (43°09′11″N and 11°17′18″E) situated along the northeastern border of the Maremma in the Siena Province, in a range of low hills (at 300–400 m a.s.l.), considered as the northern limit of the An. labranchiae range in Italy. The site is characterized by swine rearing and rice (70 ha) and maize cultivation.
Mosquito collections
Two-day entomological surveys (adult and larval collection) were carried out every 15 days, from April to October in both 2005 and 2006, to investigate the presence and abundance of anopheline species. Catches were performed with different methods.
Adult collection
In each site, adult resting females were collected in selected premises: most of them (4–6 by site) corresponded to animal shelters (cowshed, horse stables, pigsty, sheep pen, henhouse, rabbit hutch, and dog's bed), while a minor fraction (1–2 by site) to human dwellings (milking room, hayloft, garage, and veranda). Resting females were always exhaustively collected, early in the morning (7:00–10:00 a.m.) by oral or battery powered aspirators.
CDC-light traps, with and without CO2, were hung in selected places from sunset to 9:00 a.m.
Human bait collections were performed in all four sites in 2005, and in sites 2 and 4 only in 2006. Monthly night catches were carried out between June and September, from 20:00 p.m. to 2:00 a.m. (Romi et al. 1997a), by two couples located in selected places.
Larval collection
Larvae were sampled using a standard 500 mL enamel dipper. In the ricefields 100 dips were taken along the perimeter of selected paddies for the study (about 2 ha of surface and about 400–600 m of perimeter). Stream, irrigation, and drainage canals were sampled by 10–20 dips, depending on their size.
Laboratory processing
Adult and larval specimens were identified according to morphological keys (Rioux 1958, Romi et al. 1997b) and stored at −20°C. For each sample of each survey, a fraction of An. maculipennis s.l. gravid females was induced to lay eggs to identify species by observation of exochorion ornamentation (Angelucci 1955). Mosquitoes with ambiguous ovipositions and a representative number of An. maculipennis s.l. adults and larvae were processed for species identification by multiplex PCR according to Proft et al. (1999). To confirm the reliability of species identification by egg morphology, several specimens were also analyzed by multiplex PCR. Besides, to assess intraspecific heterogeneity and to confirm the results of multiplex PCRs, a random sample from each site per year was further processed, yielding ITS2 sequences, according to a previously described procedure (Collins and Paskewitz 1996, Marinucci et al. 1999). The PCR products were purified using Microcon-PCR (Millipore, Billerica, MA) and directly sequenced at MWG Biotech AG (Ebersberg, Germany) using the same primers as those used for PCR in both forward and reverse directions.
Alignment between homologous sequences of specimens previously collected in the Tuscany Region and available in GenBank (AY232827, AY238424, AY238425, AY238408, and Z50103) and those generated in this study was performed by DS Gene v1.5 (Accelrys Inc. 2003).
Seasonal dynamics were elaborated considering the whole An. maculipennis s.l. sample from resting sites and reported as a monthly average number of females. Parity rate was assessed by ovary dissection of unfed-females (Detinova 1962) collected on human bait and CDC traps in site 2 only.
Feeding preferences were investigated on freshly fed females from indoor resting places (both animal shelters and human dwellings) by a direct ELISA on nitrocellulose membrane (Bongiorno et al. 2003). Blood meal sources were assessed using peroxidase-labeled anti-animal sera (Sigma, St. Louis, MO)—namely, anti-bovine (A-5295), anti-chicken (A-9046), anti-dog (A-6792), anti-horse (A-9292), anti-human (A-8794), anti-pig (A-5670), anti-rabbit (A-6154), and anti-sheep (A-3415).
All anopheline specimens used for parity analysis, trophic activity, and blood meal analysis were identified by molecular method.
Statistical analysis
The differences among all sites in 2005 and 2006 were evaluated by Kruskal–Wallis test, and Mann–Whitney test was used to calculate the difference in each site between the 2 years. Parity rates and human bait collections were analyzed via χ 2 analysis and χ 2 test for trend, respectively. All statistical tests were considered significant at the α = 0.05 probability level. Statistical analyses were processed by EpiCalc 2000 v.1.02 and GraphPad PRISM v.3.02 software.
Results
During the 2005 and 2006 surveys, a total of 8274 mosquitoes were collected in the selected sites, 7691 (93%) were anophelines, of which 7518 (97.7%) belonging to the An. maculipennis complex. Fourteen species belonging to five genera were identified in the total sample: An. labranchiae; An. maculipennis s.s.; Anopheles melanoon Hackett, 1934; An. atroparvus; Anopheles claviger (Meigen, 1804); Anopheles plumbeus Stephens; Ochlerotatus caspius (Pallas, 1771); Ochlerotatus detritus Haliday, 1833; Aedes albopictus (Skuse, 1897); Culex pipiens Linné, 1758; Culex impudicus Ficalbi, 1890; Culex hortensis Ficalbi, 1899; Culiseta longiareolata (Maquart, 1838); and Culiseta annulata (Schrank, 1776).
As our study focused on An. maculipennis s.l., results of collections by site and by capture method are referred to this complex only (Table 1).
ND, not done.
About 15% (n = 1111) of the whole An. maculipennis s.l. sample was identified by the egg morphology or by multiplex PCR; besides, 25 females were simultaneously identified using both methods and no diagnostic discrepancy was observed. Nineteen percent (n = 116) of the sample already identified by multiplex PCR generated sequences of An. labranchiae, An. maculipennis s.s., An. Melanoon, and An. atroparvus, sharing 100% identity with the homologous sequences of specimens from the Tuscany Region available in GenBank. No intraspecific variations were detected in the samples examined.
Dynamics and species composition of the An. maculipennis complex
Seasonal dynamics of the An. maculipennis s.l. populations from all sites were calculated using resting mosquitoes only and are reported in Figure 2. Species composition and relative frequencies (%) by site of members belonging to An. maculipennis complex are showed in Table 2. Significant differences in the relative abundance of An. maculipennis s.l. were found among sites in both years (p < 0.05).
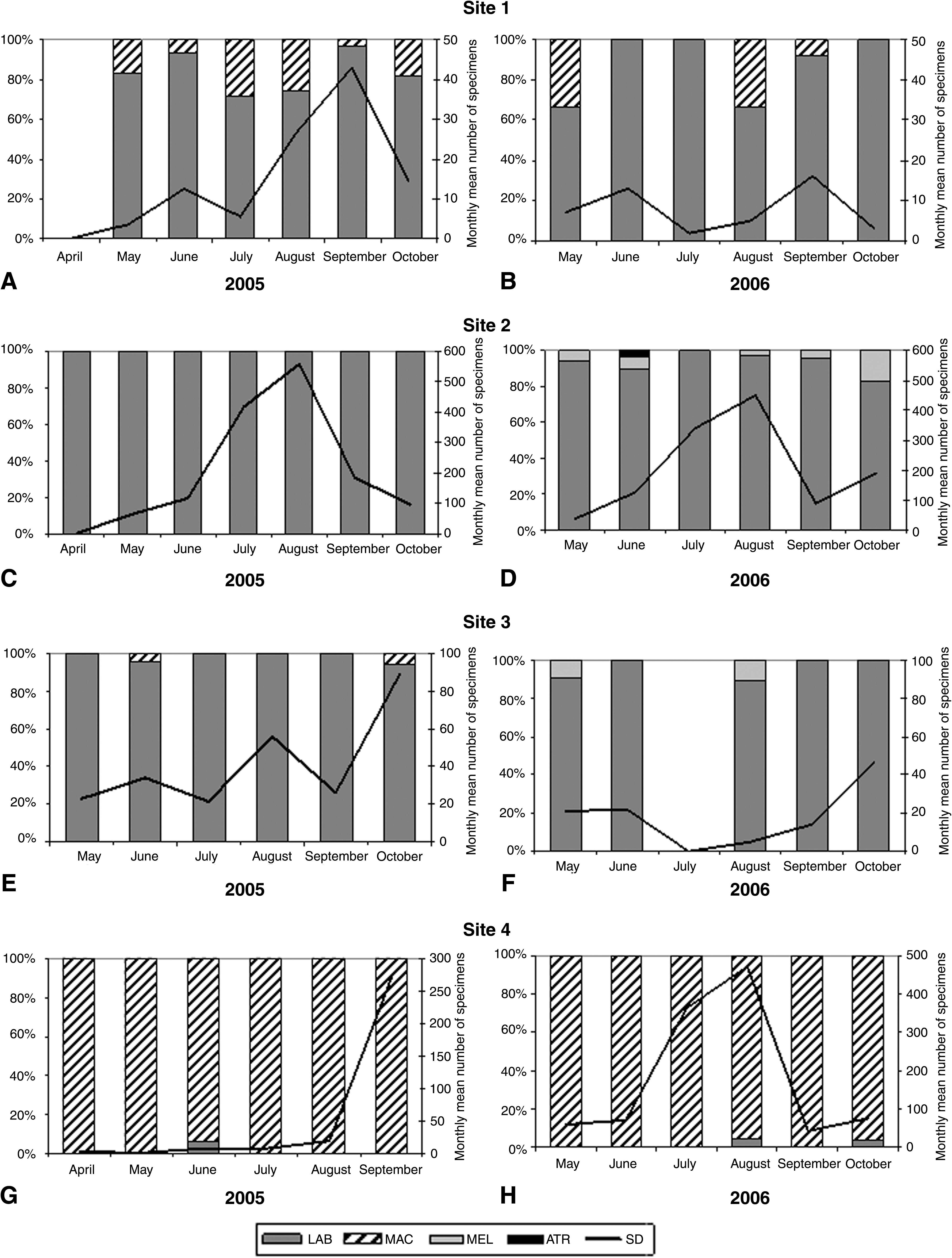
Seasonal dynamics (lines) and relative composition in species (histograms) of Anopheles maculipennis complex in the four sites. Histograms represent the percentage of species morphologically and/or molecularly identified, as reported in Table 2. Site 1: (
LAB, An. labranchiae; MAC, An. maculipennis s.s.; MEL, An. melanoon; ATR, An. atroparvus.
Site 1—About 51% (n = 142) of the specimens was morphologically and molecularly identified. An. labranchiae resulted as the predominant species in both years (>85%), with respect to An. maculipennis s.s. (Table 2). In site 1, no significant difference in abundance was found in both years (p > 0.05). Seasonal dynamics of the local An. maculipennis s.l. population followed a similar trend in 2005 and 2006. In both years a coincident first peak occurred in the middle of June (12.5 and 13 specimens/month in 2005 and 2006, respectively). A second peak occurred in September, with higher abundance (43 and 16, respectively) (Fig. 2A, B).
Site 2—About 12% (n = 566) of the An. maculipennis s.l. sample was identified. In 2005 An. labranchiae was the only species recorded, while in 2006 two other species of the complex, An. melanoon and An. atroparvus, were found, representing about 4.9% and 0.7% of the whole sample, respectively (Table 2). In site 2, no significant difference in abundance was found in both years (p > 0.05). Seasonal dynamics of An. maculipennis s.l. showed a similar trend during the study period, beginning with a gradual growth during spring that became more drastic in June, peaking in August (558 and 449 specimens in 2005 and 2006, respectively) and decreasing in September (186 and 90 specimens in 2005 and 2006, respectively). In October 2005 the abundance of mosquitoes continued to fall (97 mosquitoes/month), whereas in 2006 an inversion in the trend (189 mosquitoes/month) was recorded (Fig. 2C, D).
Site 3—Over 34% (n = 214) of An. maculipennis s.l. specimens was identified. An. labranchiae resulted the most abundant species (>96%, Table 2), being in sympatry with other two species, An. maculipennis s.s. in 2005 and An. melanoon in 2006, respectively (Table 2). The season 2005 was characterized by a higher mosquito density with respect to 2006 (p < 0.05). A similar first peak occurred in June (34 specimens in 2005 vs. 22 in 2006), but while in 2005 a second peak occurred in August (56 mosquitoes/month) followed by a third one in October (89), in 2006 after the first peak, a dramatic decrease in abundance was recorded. The complete absence of mosquitoes in July was due to an unexpected insecticide treatment of animal shelters. However, a new, slow but constant increase in abundance was recorded from August to October, with 46 specimens/month at the end of the season (Fig. 2E, F).
Site 4—About 11% (n = 214) of the An. maculipennis s.l. sample was identified. An. labranchiae was found in low density (1% in 2005 and 3% 2006), resulting An. maculipennis s.s. the predominant species (>97%) in both years (Table 2). In 2005 and 2006, besides differences in abundances (p < 0.05), the seasonal dynamics showed a noteworthy different trend (Table 1), probably due to the absence or presence of livestock rearing in the farm (Fig. 2G, H). In 2005 the dynamic was characterized by a complete absence of any peak up to September, with a sudden increase at the end of the season, when a cycle of swine rearing started. In 2006, when swine rearing lasted the whole year round, a gradual increase of the mosquito population was recorded, which reached a peak in August (about 450 specimens) and decreased dramatically in September.
Breeding sites
During the whole study period, the potential anopheline breeding sites, previously identified around the four sites selected, were regularly visited during 2005 and 2006 surveys. All irrigation and drainage canals were positive for anophelines, showing a density of larvae/dip, ranging from 0.1 to 1.9. In the stream of site 3, the larval density ranged from 0.7 to 2.3. In the ricefields of site 2, larval density varied between 0.14 and 1.5, and in site 4 between 0.05 and 0.7. The molecular identification of larval specimens from each site always confirmed the same species composition obtained from processed adults (data not shown).
Parity rate, trophic activity, and feeding preferences
The molecular analysis of specimens processed for these purposes identified An. labranchiae as the only species present in sites 1, 2, and 3 and only An. maculipennis s.s. in samples from site 4.
In both 2005 and 2006 from June to August, parity rate was assessed for site 2 only (Fig. 3). This choice was suggested by our previous studies (Romi 1999), showing An. labranchiae as the predominant species in this site, where it seasonally reached the highest abundance of the whole study area. Parity rates showed a similar trend in 2005 as well as in 2006. Although the value in June 2005 appeared considerably lower (10%) than that obtained in 2006 (30%), as well as in July (p < 0.05), in August the increases of age populations reached similar values around 70–75% in both years (p > 0.05).
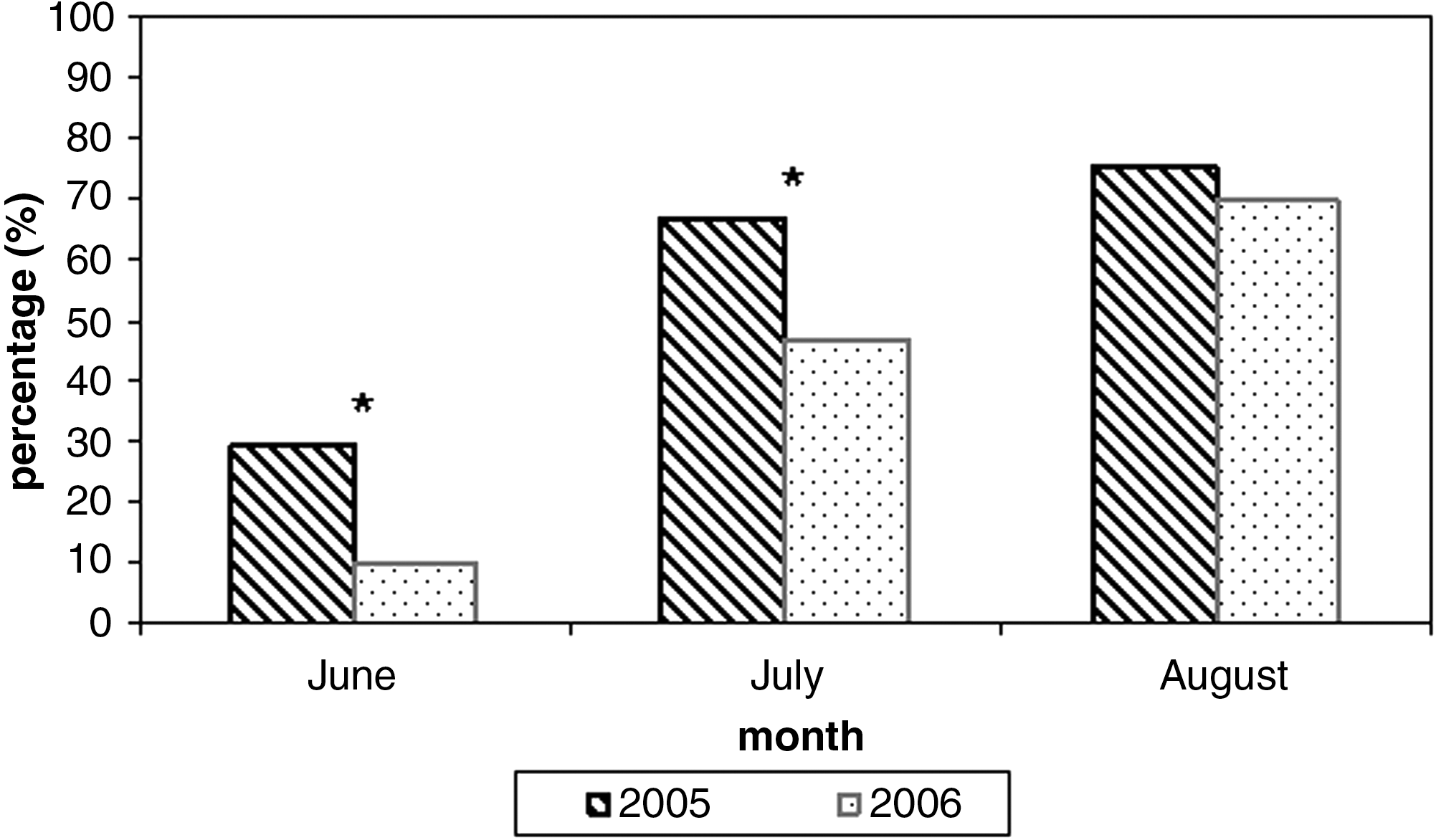
Parity rates of An. labranchiae specimens collected in site 2, from June to August in 2005 and 2006 (*p < 0.05).
The results of night catches carried out in 2005 and 2006 are reported in Table 1. Based on the results of 2005 and according to our previous experiences (Romi et al. 1997a), in the 2006 catches were carried out in sites 2 and 4 only, where the abundance of An. labranchiae and An. maculipennis s.s., respectively, allowed us to further our study of both species. In site 2, most of An. labranchiae was collected during the first hour of exposure (h 20:00–21:00), even if the human biting activity remained relatively high in the following hours until h 24:00 (Fig. 4A); a statistical significance between the 2 years was observed (p < 0.05). In site 4, no significant differences were found (p > 0.05), and trophic activity of An. maculipennis s.s. appeared rather high during the first 2 h, dramatically decreasing at h 22:00, ending at h 23:00 (Fig. 4B).
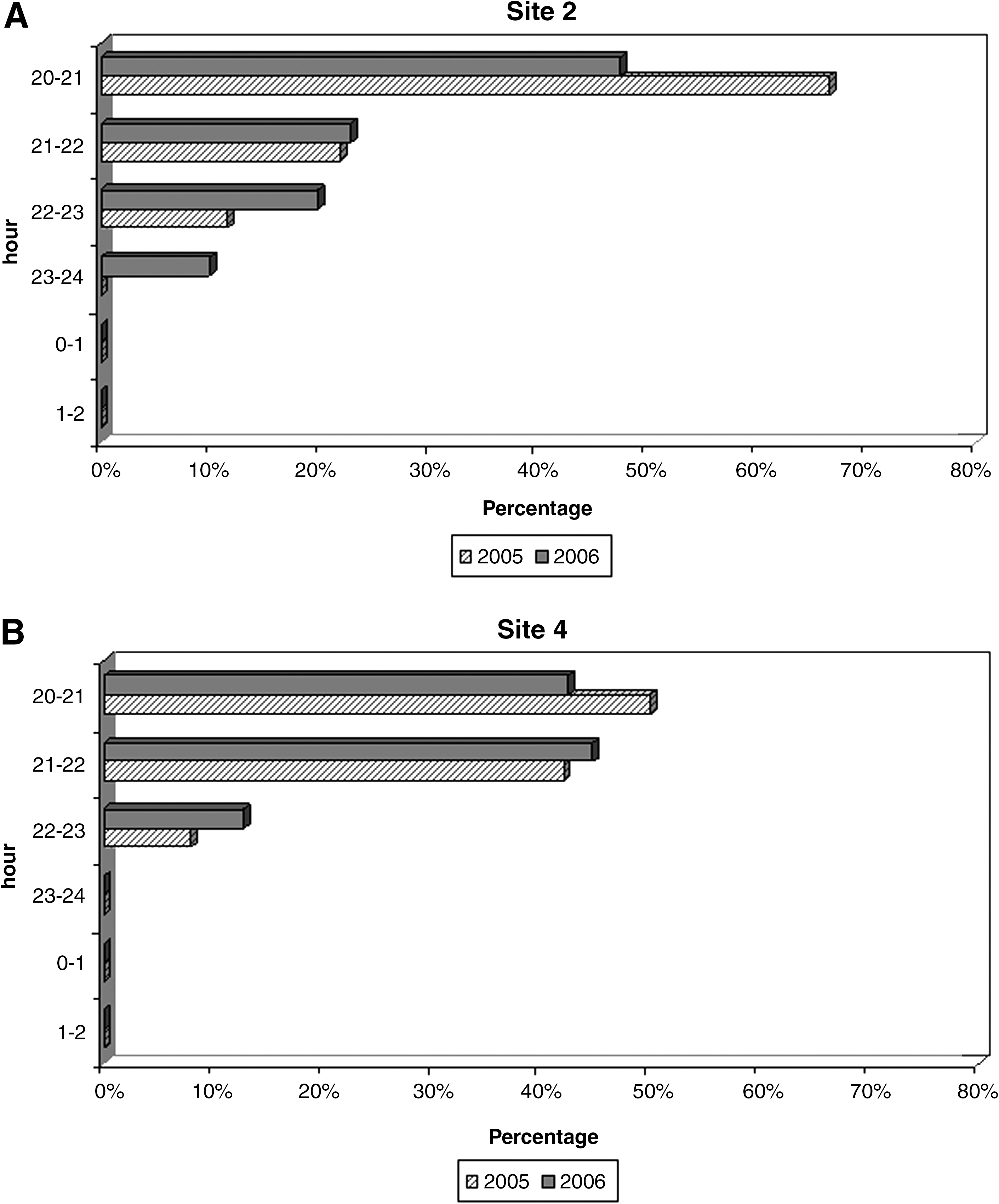
Hourly human bait collection in site 2 (
A total of 258 freshly fed females collected in both years were analyzed to establish the source of blood (Table 3). About 95% of processed specimens came from animal shelters, while the remaining 5% from human dwellings, reflecting the same ratio of the whole sample from resting sites (data not shown). The blood meal sources were unambiguously identified as coming from eight host animals, and no mosquito contained a double feed. An. labranchiae (n = 237) exhibited a wide feeding preference, depending on the host available, from big mammals (bovine, sheep, pigs, and horses) to poultry. Only three specimens, collected in a garage from site 2, resulted positive to anti-human serum. An. maculipennis s.s. blood meals (n = 21) were taken on pig and on poultry.
LAB, An. labranchiae; MAC, An. maculipennis s.s.
Discussion
According to our findings, foci of anophelines belonging to the An. maculipennis complex were found all over the study area. However, species composition, seasonal abundance, and population dynamics showed differences related to the variety of biotopes and human activities that characterized the four study sites. An. labranchiae is the predominant species in the central part of the area (sites 2 and 3), in particular where ricefields (site 2) allow the species to reproduce massively during the warm season. The species also results prevalent in the southern part of the study area (site 1) where, however, it does not reach high levels of abundance, being mainly associated with temporary breeding sites. In site 4, the different environmental conditions of the area (300–400 m a.s.l.) favor a huge development of An. maculipennis s.s., An. labranchiae representing only 1–3% of the whole sample. Nevertheless, this small fraction of An. labranchiae represents the first record of the species in this internal part of the study area.
The seasonal dynamics of An. labranchiae populations reported in the central part of the study area follow a typical thermophilic trend where ricefields ensure a constant presence of water (site 2), while they show an irregular trend where permanent breeding site depends mainly on rainfall (site 3). An. labranchiae parity rate seems consistent with that trend, as previously assessed by studies carried out in the same area in the 1990s (Romi 1999, Romi et al. 2001). The lowest values of June reflected the young age of populations; in July, the rising values were consistent with the population growth; the highest values were recorded in late August, when the abundance of the population peaked.
Trophic activity of An. labranchiae and An. maculipennis s.s. lasts from dusk to the beginning of the night, being mainly concentrated in the first hours after sunset. An. labranchiae, as expected, results as being very aggressive during human bait catches, confirming the high anthropophilic degree of this species (Missiroli et al. 1933, Romi et al. 1997a). It is interesting to note that An. maculipennis s.s., although considered mainly zoophilic (Missiroli et al. 1933, Zahar 1990), also results as being very aggressive on humans during the night catches, both in presence or absence of animals.
The results of blood meals analysis showed that An. labranchiae fed on a wide range of hosts, from big mammals to poultry, and also on humans when they are available. It is worthy to note that, in spite of the small number of human blood meals (only three) found, the discrepancy between our findings of ELISA analysis and human bait catches remains only apparent, because of the real difficulty for mosquitoes in accessing humans.
Although the results of our study highlight in site 2 a particularly high receptivity (vector predominance and abundance, anthropophily, aggressiveness on humans, and high parity rate during the warmer season), the whole study area (sites 1, 3, and 4) may be still considered at risk for malaria reintroduction, as well as other regions of southern European countries recently investigated (Merdić and Boca 2004, Lopes et al. 2005, Patsoula et al. 2007, Ponçon et al. 2007). Within this context, the Maremma—in general terms, Italy—represents a suitable model of study, because of the presence of An. labranchiae (Romi et al. 1997a, Romi 1999), the most efficient vector when malaria was endemic in the countries of Western Europe. Moreover, in Central Italy, An. labranchiae is probably extending its range northward and eastward, across to the internal hills of the Tuscany, Umbria, and Latium regions (Boccolini et al. 2004, and our unpublished data), where in the past, low levels of malaria transmission were supposed to be maintained by An. atroparvus and/or Anopheles messeae Falleroni, 1926 only (Missiroli et al. 1933, Bettini 2002). At present it is very difficult to establish if our findings could be the result of a new, slow but real expansion of An. labranchiae in these internal areas (after the malaria control campaign), or if, more simply, its presence was not detected in the past because of the ambiguous characters of the egg morphology that did not permit us to distinguish this powerful vector from the other two species, when in sympatry.
In conclusion, 60 years after malaria eradication in Italy, as well as in the other European developed countries, we may reasonably assume that the reintroduction of endemic malaria is an unlikely but not an impossible event (Jetten et al. 1996, Rogers and Randolph 2000, Kuhn et al. 2002, WHO 2007). We cannot exclude that sporadic, isolated P. vivax cases (as occurred in 1997) or even small, limited outbreaks may occur in Central Italy, when climatic conditions and vector densities are favorable to this event, together with the possible presence of gametocyte carriers, both those officially reported (Romi et al. 2006) and above all those not recorded (unknown number).
Footnotes
Acknowledgments
We are grateful to Angelo Tamburro, Maurizio Cocchi, Giorgio Pontuale, and Irene Raffaelli of Environmental Zoology Unit, ASL 9, Grosseto, for logistic support and their precious information concerning the Maremma. We also thank the Angiolini, Berni, Borracelli, Fini, Mattei, Raffi, and Stefanelli families for all their kind help. A special thanks to Antonino Bella for statistical consultancy.
This research was supported by ISS and funded under the EU 6th Framework Program (GOCE-CT-2003-010284 EDEN). It is officially catalogued by the EDEN Steering Committee as EDEN0118 (EDEN). The contents of this publication are the sole responsibility of the authors and do not necessarily reflect the views of the European Commission.
Disclosure Statement
No competing financial interests exist.