
Abstract
We sought to elucidate the role of migratory birds in transmission of H5N1 in an enzoonotic area. Resident, captive, and migratory birds were sampled at five sites in Java, Indonesia. Mist nets were used to trap birds. Birds were identified to species. RNA was extracted from swabs and reverse transcriptase polymerase chain reaction (RT-PCR) conducted for the HA and M genes of H5N1. Antibodies were detected by enzyme-linked immunosorbent assay and hemagglutination inhibition test. Between October 2006 and September 2007, a total of 4,067 captive, resident, and migratory birds comprising 98 species in 23 genera were sampled. The most commonly collected birds were the common sandpiper (6% of total), striated heron (3%), and the domestic chicken (14%). The overall prevalence of H5N1 antibodies was 5.3%. A significantly higher percentage of captive birds (16.1%) showed antibody evidence of H5N1 exposure when compared to migratory or resident birds. The greatest number of seropositive birds in each category were Muschovy duck (captive), striated heron (resident), and the Pacific golden plover (migratory). Seven apparently well captive birds yielded molecular evidence of H5N1 infection. Following amplification, the HA, NA, and M genes were analyzed. Phylogenetic analysis of the HA gene showed that the isolates were 97% similar to EU124153.1 A/chicken/West Java/Garut May 2006, an isolate obtained in a similar region of West Java. While no known markers of neuraminidase inhibitor resistance were found within the NA gene, M segment analysis revealed the V27A mutation known to confer resistance to adamantanes. Our results demonstrate moderate serologic evidence of H5N1 infection in captive birds, sampled in five sites in Java, Indonesia, but only occasional infection in resident and migratory birds. These data imply that in an enzoonotic region of Indonesia the role of migratory birds in transmission of H5N1 is limited.
Introduction
Wild birds harbor 16 hemagglutinin (HA) and nine neuraminidase (NA) subtypes of influenza A and are natural reservoirs of the virus (Munster et al. 2007, Brown et al. 2007). The role of migratory birds in the spread of HPAI remains under considerable debate. Some have attributed transmission of H5N1 to water bird migration (Chen et al. 2005, Normile 2006b), although conclusive evidence is sparse (Fergus et al. 2006). The outbreak of H5N1 in wild migratory birds at Lake Qinghai, China in May 2005 led to the deaths of over 6,000 birds (Lei et al. 2007, Wang et al. 2008). Lake Qinghai in western China is a major breeding site for migratory birds whose flyways extend to Southeast Asia, India, Serbia, Australia and New Zealand. This event resulted in the careful evaluation of the role of wild birds in maintaining and spreading the H5N1 virus. Isolation of H5N1 virus has been reported in over 60 species of wild birds (Ellis et al. 2004, Chen et al. 2005, Kwon et al. 2005, Liu et al. 2005). Still, the extent to which free-flying wild birds contribute to sustaining and spreading the disease remains to be determined.
In Indonesia, where the greatest number of human H5N1 cases and fatalities has been recorded, H5N1 is enzoonotic, with 31 out of 33 provinces having reported infection (personal communication, Ministry of Agriculture). H5N1 virus infection among poultry was first recognized in Indonesia in December 2003. In July of 2005, the first human case of H5N1 was reported (Kandun et al. 2006). Since that time, the World Health Organization (WHO) has confirmed 135 cases of human infection resulting in 110 deaths (case fatality rate = 81%; WHO, 21 June 2008). Transmission of H5N1 to humans has largely resulted from interaction with dead or dying poultry (Sedyaningsih et al. 2007).
The Indonesian archipelago is home to 1,598 bird species representing 17% of the world’s total species (Aamir et al. 2007). Of these species, 372 are endemic to Indonesia and 149 are migratory. In Indonesia, migratory pathways are well elucidated only for shorebirds and migratory ducks and geese. Shorebirds use the East Asian–Australasian Flyway (Melville and Shortridge 2004) and the migratory ducks and geese use the East Asian and Central Asian flyways. In 2005, there was no systematic surveillance in Indonesia to ascertain the potential introduction of H5N1 from migratory birds to domesticated poultry flocks. To elucidate the role of migratory birds in AI transmission in Java, Indonesia, a prospective surveillance study was initiated to screen for either acute or previous exposure of H5N1 in migratory birds. To ascertain possible vectors of infection, data were collected in paired resident and captive birds in the same regions. Here we report the results of a 13-month surveillance conducted in five sites spanning Java Island, Indonesia.
Materials and Methods
Site selection and bird sampling
From October 2006 to September 2007, samples were collected from captive, resident, and migratory birds at five sites located along major migratory paths within Java, Indonesia (Fig. 1). These included: Muara Gembong, Bekasi, West Java (5° 60′ S; 107° 03′ E); Eretan, Indramayu, West Java (6° 32′ S; 108° 30′ E); Sayung, Demak, Central Java (6° 55′ S; 110° 29′ E); Trisik, Kulonprogo, Yogyakarta (7° 19′ S; 112° 48′ E); and Wonorejo, Surabaya, East Java (7° 58′ S; 110° 11′ E). Sampling occurred in natural wetlands (including mangroves, tidal mudflats, and sand dunes) and human settlements with varying levels of modified habitats. Birds were sampled monthly with the exception of one field site (Muara Gembong) where severe flooding occurred during February 2007.
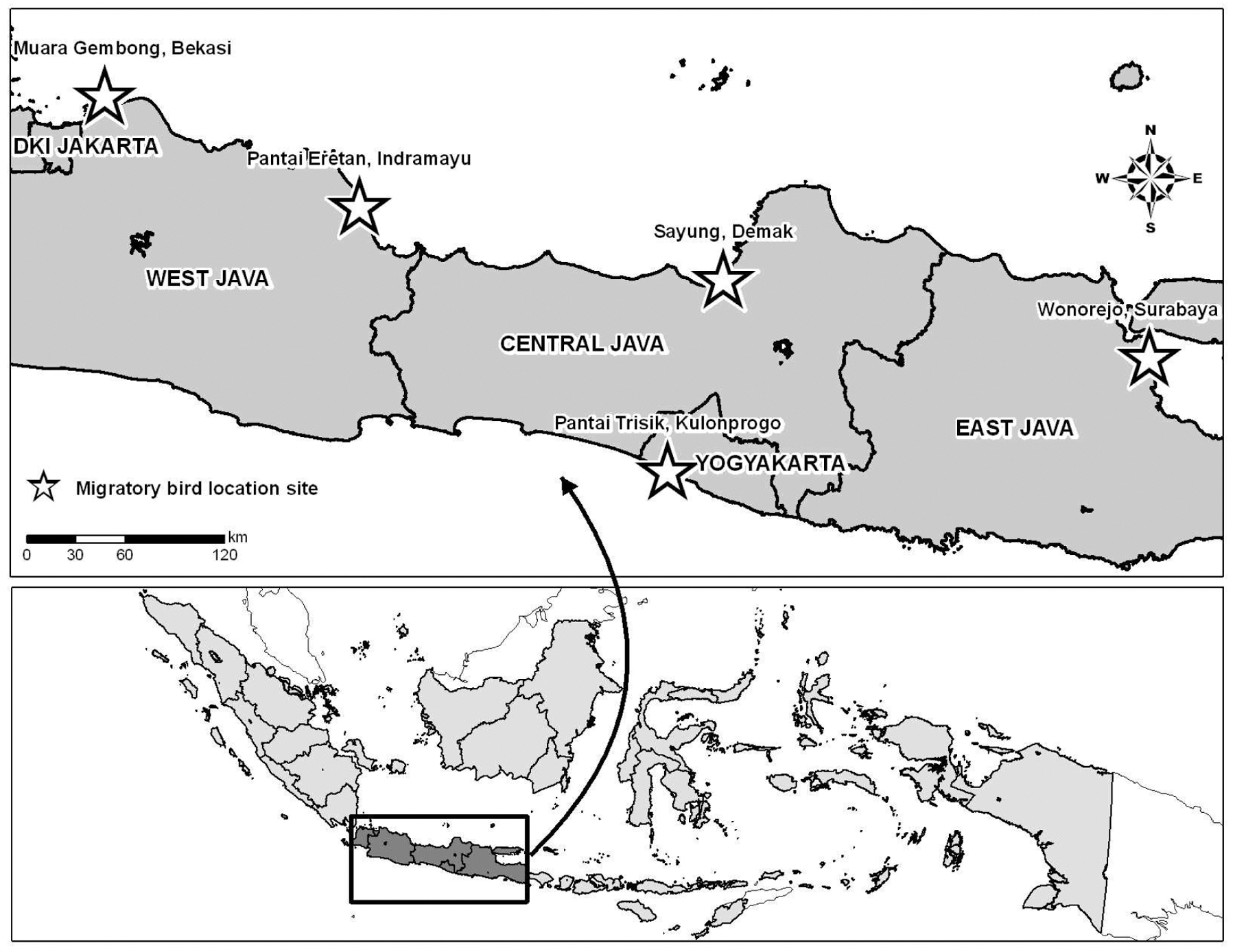
Map of Java Island, Indonesia showing migratory, resident, and captive bird H5N1 surveillance sites, October 2006 to September 2007.
Sampling of migratory, resident, and captive birds was conducted under a protocol approved by the Health Research Ethics Committee of National Institute of Health Research and Development, Ministry of Health of the Republic of Indonesia and the Naval Medical Research Unit #2 Institutional Animal Care and Use Committee. Permits to survey wild birds were issued to Yayasan Kutilang Indonesia under supervision of the Indonesian Ornithologist’s Union by the Ministry of Forestry, Republic of Indonesia. Migratory birds, resident wild water birds, captive birds, domestic waterfowl, and commercial poultry were sampled and screened for using established guidelines (Aavitsland 2005). Birds were collected either by hand, mist, or sweep net and placed into a dark bag to reduce stress. Wild birds were banded and identified to species. Domestic waterfowl and poultry from neighboring farms were collected by hand and processed. Birds recovered and were released following specimen collection.
Specimen collection
Blood, throat, and cloacae swabs were generally collected from each bird, although the small size (<30g) of several species precluded blood sampling. All efforts were made to minimize handling and confinement of birds. Once sampling was complete, birds were returned to the field and released. Serum and swab samples were shipped to U.S. Naval Medical Research Unit #2 laboratory in Jakarta for processing and analysis.
H5N1 serology
Serum samples were tested by Wantai H5-HA (Ab) ELISA (Beijing Wantai Biological Pharmacy Enterprise Ltd., Beijing, China) to detect H5N1 antibody per manufacturer’s instructions. Twenty percent of each species captured per month were screened by hemagglutination inhibition test (HI) (Stephenson et al. 2005).
Polymerase chain reaction amplification of HA and NA
RNA was extracted from cloacae and throat swabs using Imp Viral RNA mini kits (QIAGEN Gumbo, Hilden, Germany) following manufacturer’s instruction and stored at −70°C. Samples were tested for the presence of the following influenza targets using real-time polymerase chain reaction (PCR) assay against several gene targets: Influenza a matrix, H5-HA, H7-HA, and H9-HA genes. One-step reverse-transcriptase PCR (RT-PCR) was performed in a final volume of 25, μl containing 5 all of extracted RNA. Real-time assays were carried out in a Real-Time PCR System 7900HT (ABI, Foster City, CA) with the same cycling conditions for all gene targets: 50°C for 30 min, 95°C for 2 min, and 45 cycles with 95°C for 15 sec, 55°C for 30 sec.
N1-gene detection confirmed H5HA results as previously described (Heine et al. 2007) with modification as follows: 5 μl of Siegen one-step RT-PCR buffer, 400 μm of each dent, 0.6 μm of each primer forward and reverse, 1 μl of Q-enzyme mix (QIAGEN Gumbo, Hilden, Germany) with the addition of 10 units of Nase-inhibitor and nuclease free water to the final volume of 20 μl. Extracted RNA (5 μl) was added as a template. Cycling conditions were as follows: 50°C for 30 min, 95°C for 15 min, 10 cycles of 95°C for 30 sec, annealing at 60°C for 30 sec (with 1°C decreasing in each cycle), 72°C for 1 min followed by 40 cycles of 95°C for 30 sec, 55°C for 30 sec and 72°C for 1 minute with a final extension at 72°C for 10 minutes. PCR amplification was visualized on 2% agar gels with expected product size of 615 bp.
HA, NA, and M gene sequencing
RT-PCR was performed to amplify large fragments of HA, NA, and M genes. Amp icons were purified using QIAquick PCR Purification kit (QIAGEN Gmbh, Germany). DNA sequencing reactions were conducted using the Big Dye Terminator V.3.1 Cycle Sequencing Ready Reaction kit (ABI, Foster City, CA) following priming with an internal primer and precipitated using BigDye X-terminator purification kit (ABI). Amplicons were resolved on an ABI-Prism 3130 XL Genetic Analyzer (PerkinElmer, Norwalk, CT), and sequences were edited and aligned using Sequencher 4.8 (Gene Codes Corporation, Ann Arbor, MI) and Bioedit v.5.0.6 (North Carolina State University, NC) the nucleotide sequences generated were compared on GenBank (
Phylogenetic analysis
Nucleotide sequences were aligned using ClustalW and ends trimmed to equal lengths. Using this alignment, a neighbor-joining tree was constructed using MEGA 4.0.1 with 1,000 bootstrap replicates. Clade 0 was defined as an outgroup to root the tree. Maximum-likelihood trees were generated in PAUP 4.0 beta to confirm clade topology. The nucleotide sequences derived from this study are posted on GenBank as dkIndramayu90461507 and dkIndramayu90461807.
Results
From October 2006 to September 2007, specimens were obtained from 1,110 captive, 1,417 resident, and 1,540 migratory birds captured at five sites (Fig. 1). Sampled birds comprised 98 species in 23 genera. In compliance with the sampling scheme, the number of resident and captive birds did not deviate greatly among sites over time. Greater numbers of birds were sampled in Trisik and Wonorejo between September and April and August to March, although whether this phenomenon was a result of seasonal variation or effectiveness of sampling strategies cannot be ascertained. Seasonality in migratory birds was not evident at Eretan, Maura Gembong, or Sayung (data not shown). The most commonly collected migratory bird was the common sandpiper (Actitis hypoleucos, 17%). Eight percent of resident birds captured were striated heron (Butorides striatus), while the chicken (Gallus gallus bankiva, 55%) was the most common captive bird sampled in the current study (Table 1). Thirty-seven different species of migratory birds, 56 species of resident birds, and five species of captive birds were sampled over the 12-month study.
ELISA, enzyme-linked immunosorbent assay; RT-PCR, reverse-transcriptase polymerase chain reaction.
Serologic testing
Total H5N1 prevalence as determined by enzyme-linked immunosorbent assay (ELISA) was 5% (215 of 4067 birds tested using ELISA) among all birds sampled from all sites (Table 1). Serology-positive birds were evident year-round at all five sites (data not shown). Trisik had the greatest percentage of ELISA-positive results (7%). The majority of ELISA positives (179/1110) were captive birds, with Muschovy ducks (53) and chickens (95) primarily demonstrating serologic evidence of infection (Table 1). Overall, prevalence was 1% (15/1417) among resident and 1% (21/1540) among migratory birds collected at all sites. Of the ELISA-positive resident birds, eight species were represented. A total of five striated herons had detectable antibodies to H5N1. Striated herons made up over 8% (120/1513) of the total resident bird population sampled. While 1% of migratory birds was ELISA positive, apparent infection was seen in all 12 species sampled. HI serology tests were conducted on 20% of all ELISA-positive bird samples to support ELISA results. The possibility of cross-reactive antibodies could not be ruled out. A significant correlation between ELISA and HI testing was found using Pearson’s method (p < 0.001), with an overall agreement of 96.4%. While records were incomplete regarding the general health and survival of the captured birds, field teams indicated that only on rare occasion did captured birds show signs of illness.
Molecular testing
In order to demonstrate acute infection in birds, throat and cloacae swabs were tested by rRT-PCR for evidence of the H5N1 genome. Overall, only 7 of 1,655 (0.4%) birds collected showed PCR evidence of H5N1 infection. These seven birds were also ELISA positive. All seven birds that tested positive by PCR for H5N1 were captive bird species. These birds were collected at the Eretan site (Muschovy (n = 4) and mallard (n = 2) ducks) and Muara Gembong (a single Muschovy duck). Positive samples were obtained from throat swabs from seven birds of the positive birds and from a cloacae swab from one a single Muschovy duck collected from Muara Gembong. Interestingly, the rRT-PCR-positive birds were captured in areas of West Java where numerous human H5N1 infections have been previously documented (Fig. 2).

Map of Java Island, Indonesia showing migratory, resident, and captive bird H5N1 surveillance sites and locations of reported human H5N1-positive cases (as of June 21, 2008).
Sequence analysis of the HA, NA, and M gene segments was conducted on diagnostic samples from two of the birds that were determined to be positive for H5N1 by rRT-PCR. Sequence analysis of the HA gene determined that these viruses contained the multiple basic amino acid motif at the cleavage site, a characteristic feature of highly pathogenic avian influenza viruses. Phylogenetic analysis of the HA gene (Fig. 3) determined that, similar to other H5N1 viruses from Indonesia, these viruses belonged to H5N1 Clade 2.1 viruses. Sequence of the HA, NA, and M gene segments from two captive ducks sampled in West Java were compared to previous sequences deposited in GenBank. The isolates were 97% similar to EU124153.1 A/chicken/West Java/Garut May 2006, an isolate obtained in a similar region of West Java. The A156T substitution that results in glycosylation of asparagine at 154 was not observed in the two samples sequenced. While no known markers of antiviral resistance to neuraminidase inhibitors were found within the NA gene, M segment analysis indicated the presence of the V27A mutation that is known to confer resistance to adamantanes.

Neighbor-joining phylogenetic tree of HA from H5N1 viruses with 1,000 bootstrap replicates. The tree was rooted with Clade 0 as an out-group and bootstrap values for clade defining nodes are indicated. Boxed sequences indicate the sequences obtained in this study.
Discussion
During a time of sustained poultry and human H5N1 infections in Java, we found no PCR evidence of H5N1 infection and only limited serologic evidence of exposure among migratory birds captured and sampled over 13 months at five sites in Java, Indonesia. Our data demonstrated serologic evidence of H5 infection among 16% of captive birds sampled in the same region. H5N1 Virus was also detected by rRT-PCR and characterized in two species of domestic ducks from West Java.
Only Muschovy and Mallard ducks were found to be acutely infected with H5N1 in our survey. More AI isolations have been reported from mallard ducks than any other species (Stallknecht and Shane 1988, Perkins and Swayne 2002, Sturm-Ramirez et al. 2004, Sturm-Ramirez et al. 2005). Analysis revealed that genomic sequences from viruses obtained did not have the A156T substitution that results in glycosylation of asparagine 154 and is predicted to reduce its affinity for sialosides. This change is frequently associated with viral adaptation to terrestrial poultry and increased virulence for these birds (Baigent and McCauley 2001, Mitnaul et al. 2000, Perdue and Suarez 2000). Overall, the sequences obtained from amplicons generated off the diagnostic samples were homologous to previous samples from birds in West Java.
These data support a number of studies that recommended caution in presuming the role of migratory birds in the maintenance and transmission of highly pathogenic H5N1 (HPAI) (Kilpatrick et al. 2006, Normile 2006a, Weber 2006, Weber and Stilianakis 2007). Influenza A viruses normally survive harmoniously in Waterfowl (Anseriformes) and shorebirds (Charadriiformes), although virus replicates and is excreted in their feces (Garcia et al. 1997). On occasion, HPAI viruses emerge in wild birds resulting in high rates of mortality (Gaidet et al. 2008). Such was the case in Hong Kong in 2002 (Ellis et al. 2004) and Lake Quinghai in 2006 (Lei et al. 2007). The ability of infected migratory birds to serve as effective spreaders of HPAI is certainly limited by their inability to undertake prolonged, intense migration over long distances (Weber and Stilianakis 2007, Rappole and Hubalek 2006).
It is possible that mildly ill or asymptomatic migratory birds facilitated efficient carriage of HPAI to susceptible hosts in Java. Small terrestrial birds have shown considerable susceptibility to H5N1 infection (Boon et al. 2007). Previous work has suggested that 9 of 21 introductions of H5N1 in Asian nations resulted from domestic poultry (Kilpatrick et al. 2006). Thus HPAI is likely perpetuated through domestic waterfowl (Krauss et al. 2007). It is possible that migratory birds captured in our study become infected from poultry in a highly enzoonotic area.
Certainly, continued surveillance of migratory birds, including juvenile birds, for H5N1 is warranted to track the introduction and emergence of avian influenza viruses. Furthermore, there is a continued need to monitor the wild bird and poultry interface during massive die-offs of poultry. Through effective collaboration with the Indonesian Government, national field ornithologists and volunteers can make a worthwhile contribution to Avian Influenza (AI) surveillance as well as the protection of native and migratory bird species.
Acknowledgment
The authors are grateful for the assistance of the field staff at the Indonesian Ornithologists’ Union, Yayasan Kutilang Indonesia, PILI-NGO Movement and the Animal Rescue Centers’ Network. We thank the laboratory staff at the National Institute of Health Research and Development and the Naval Medical Research Unit #2 for processing and analyzing samples and Christian Hansen for statistical analysis. This work was supported by grants from the U.S. Department of Defenses’ Global Emerging Infection Systems (DoD-GEIS), the United States Agency for International Development (USAID), and the Centers for Disease Control and Prevention. The views expressed in this manuscript are those of the authors and do not represent official policy of the U.S. Department of Defense or Department of the Navy.
Disclosure Statement
No competing financial interests exist.