
Abstract
One of the unique characteristics of West Nile virus (WNV) in North America is the large number of bird species for which the virus can be fatal. WNV mortality has been documented through experimental infections of captive birds and necropsies of free-ranging birds. Investigations of WNV-related mortality in wild birds often focus on species with dramatic population declines (e.g., American Crow, Corvus brachyrhynchos); however, few studies have addressed WNV-related mortality in species not exhibiting marked population declines since the arrival of WNV. We conducted a mark-recapture study of 204 Northern Cardinals (Cardinalis cardinalis) in an area with endemic WNV activity to estimate WNV-related mortality. Previous research has shown that once a bird is infected and recovers from WNV it develops antibodies making it resistant to future infection. Assuming that mortality risks from non-WNV causes were the same for individuals with (had been exposed to WNV) and without antibodies (had not been exposed to WNV), we compared the survival rates of birds with and without WNV antibodies to estimate the impact of WNV on wild birds. An information theoretic approach was used, and the apparent survival was found to be 34.6% lower for individuals without antibodies during the period when WNV was most active (July–September). However, the apparent survival rate was 9.0% higher for individuals without antibodies over the rest of the year. These differences in apparent survival suggest that WNV increases mortality during the WNV season and that chronic effects of WNV infection may also be contributing to mortality. Although WNV appears to have increased mortality rates within the population, population trend data do not indicate declines, suggesting that some cardinal populations can compensate for WNV-related mortality.
Introduction
Individuals that survive WNV infection develop antibodies, and are likely immune to future WNV infection (Fang and Reisen 2006, Komar et al. 2001, 2005, Nemeth et al. 2007). We assumed that survival of seropositive individuals reflects natural, population-wide survival rates in the absence of WNV. By contrast, the survival rate of birds with no previous WNV exposure (no antibodies) should be equal to natural baseline survival plus additional mortality due to WNV. Thus, by comparing survival of immune individuals (antibody positive) with vulnerable individuals (antibody negative), we can estimate mortality associated with WNV infection. However, an additional confounding factor may be that chronic effects lower the survival of seropositive individuals (Kilpatrick et al. 2006). We refer to chronic effects as an adverse effect stemming from WNV exposure that, while not directly responsible for the individual's death, contributes to its death. Some examples may be slower reflexes and therefore a higher chance of predation, or decreased foraging success and therefore a greater chance of starvation. Further, if chronic effects influence survival, individuals with WNV antibodies would have a lower survival rate than individuals without antibodies during the non-WNV season.
Materials and Methods
We captured, banded, and recorded recaptures of male and female Northern Cardinals every 2 weeks from August 2005 until June 2007 in three woodlots (8, 12, and 25 ha in size) on the South Farms of the University of Illinois, Urbana-Champaign. Cardinals were captured in seven walk-in potter traps and five mist nets positioned in the same locations throughout the woodlots, and only adult birds (i.e., after hatch year individuals) (Halkin and Linville 1999) were used in the analysis. In this location WNV has been detected since 2002, and the overall infection rate in Culex mosquitoes (the species of mosquitoes responsible for WNV transmission in the area) from the beginning of July to the end of September (the peak of WNV transmission) in the Champaign-Urbana area (which includes the study area) was 7.5 per 1000 mosquitoes in 2005 and 21.9 per 1000 mosquitoes in 2006 (Lampman et al. unpublished data).
We had 49 sampling periods with approximately equal sampling effort (3–4 days, ∼4 h in the morning per day, each 2-week period) in which Northern Cardinals were captured. Each time a bird was captured, we collected 0.1–0.2 mL of blood from the jugular vein using a 27-gauge and ½-inch needle, or from the brachial vein by making a small prick in the vein and collecting the blood via capillary tubes. An epitope-blocking enzyme-linked immunosorbent assay was performed, as previously described in Beveroth et al. (2006), to detect the presence of WNV antibodies. We used three monoclonal antibodies, 3.1112G, 2B2, and 6B6C-1. Monoclonal antibody 3.1112G is specific to WNV, whereas 2B2 reacts with both WNV and Koutango Virus and 6B6C-1 can react with all flaviviruses, including WNV.
Survival analysis
The program MARK (version 5.1) was used to generate maximum-likelihood estimates of apparent survival (ϕ), 95% confidence intervals (CI), and recapture probabilities (p). Rather than simply using recapture rates, MARK accounts for individuals that were known to be alive but were not recaptured at a specific time (i.e., individuals that were caught at a later date; p, recapture probability). We used Akaike's Information Criterion (AICc, adjusted for sample size) to evaluate a suite of candidate models (White and Burnham 1999, Burham and Anderson 2002). We assessed the fit of the best model using a bootstrapped goodness of fit test. To determine if apparent survival differed between Northern Cardinals with and without WNV antibodies, we used antibody status as a grouping variable. We also used sex as a covariate in a model to investigate if there were differences in survival between males and females. The candidate model set included constant model (“.” no change) for apparent survival and recapture probability, as well as models incorporating differences in survival or recapture probabilities. Between groups (e.g., antibody status) over time (t) and interaction between group and time were all included. We had an a priori hypothesis that individuals susceptible to WNV would have lower survival during the WNV season in central Illinois, which is from July through September. We developed a model [ϕ(season) p(t)] to test whether survival was different between the seasons (WNV season and the non-WNV season) regardless of antibody status; a model [ϕ(WNV season*anti) p(t)] to investigate whether there was a difference in survival between individuals with and without antibodies only during the WNV season; and finally a model [ϕ(season*anti) p(t)] to investigate if there were differences both between individuals with and without antibodies and during the WNV-season and the non-WNV season. In total there were 20 models; Table 1 contains the complete set of candidate models. The estimated apparent survival estimates were retrieved from the most supported model (Burham and Anderson 2002). Cardinals that acquired antibodies during the study were not included in the analysis. Because these estimates were for a 2-week period, we used the delta methods to estimate apparent survival and 95% CI over the course of a year (Powell 2007). We used a binomial test to compare the WNV-related mortality found in this study to the experimental infection conducted by Komar et al. (2005).
Models were ranked by Akaike's Information Criterion adjusted for small sample size (AICc). K is the number of parameters in the model, AICc is the difference in AICc values from the most supported model, and wi is the model weight. In these models, ϕ is apparent survival probability, p is recapture probability, (anti) is the grouping variable (individuals with and without WNV antibodies), (t) is time, (season) is grouping variable (period when WNV is active and the remainder of the year), (WNV season*anti) refers to investigated differences in survival of individuals with and without WNV antibodies only during the WNV season, (.) refers to a constant model, and (*) is an interaction effect.
WNV, West Nile virus.
To investigate whether the arrival of WNV, and the potentially increased mortality associated with WNV, affected Northern Cardinal populations, we used the Christmas Bird Count (CBC) and Spring Bird Count (SBC) data from the areas that include our study area. The CBC is conducted by volunteers who census birds within a 15-mile-diameter circle in late December and early January. For this study we used data collected from the winter of 1996–1997 through 2008–2009. The SBC is a statewide census that has been conducted since 1972 in Illinois, and is conducted by volunteers in all 102 counties on the Saturday that falls between May 4 and May 10. These volunteers record all birds, and the number of hours spent by each party in the field. Again data from 1996 through 2008 were used for this analysis. The area where this research was conducted is regularly sampled, usually by the same participants, by both the SBC and CBC. The CBC data are present at
Results
Of the 204 Northern Cardinals captured, 21 went from being WNV antibody negative to antibody positive (these were not included in the analysis), 24 remained antibody positive throughout, and 159 never tested positive for antibodies. There was an adequate fit of the model to the data (p = 0.23). The model with the most support suggests differences in the biweekly survival rate of Northern Cardinals between individuals with and without WNV, in both the WNV season and the rest of the year, and that recapture probability of the cardinals varied over time (Table 1). In our study population, there was an estimated 34.6% (i.e., 94.9% [95% CI 78.5–100.0%] antibody positive and 60.3% [95% CI 40.8–82.7%] antibody negative) reduction in survival between individuals with and without WNV antibodies during the WNV season (Fig. 1). Conversely, the trend in the non-WNV season (all months except July, August, and September) suggests that individuals without WNV antibodies had a slightly greater (9.0%) survival rate (41.1% [95% CI 24.8–58.6%] antibody positive and 50.1% [95% CI 37.7–62.5%] antibody negative). There was no evidence for sex-specific differences in Northern Cardinal survival. The apparent annual survival rate of antibody-positive individuals was 45.5% (95% CI 59.6–34.4%).
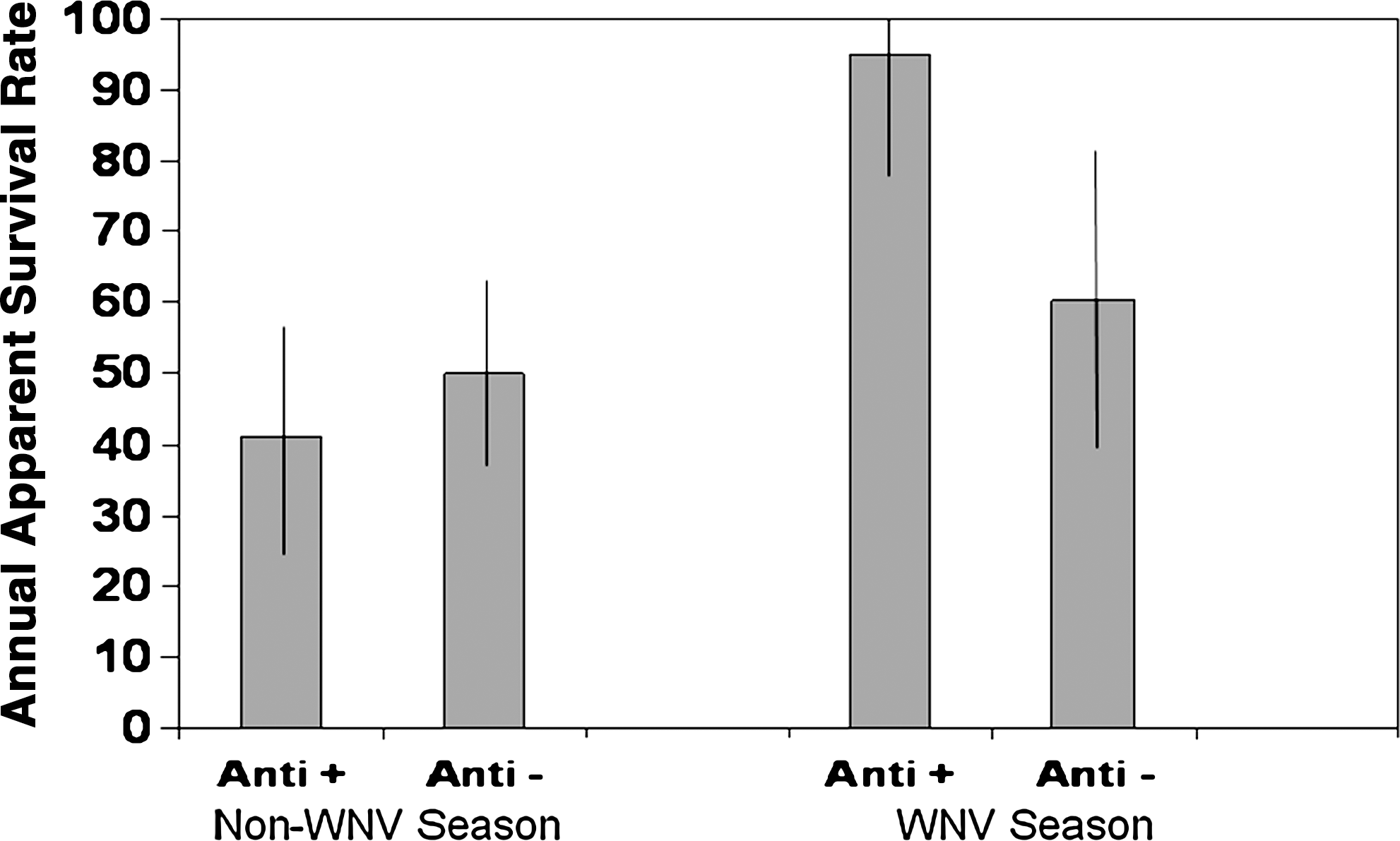
Apparent biweekly survival rates of Northern Cardinals with (Anti+) and without (Anti−) West Nile virus (WNV) antibodies during the WNV season and the non-WNV season. These estimates were generated from the most supported model [ϕ(season*anti) p(t)]. 95% Confidence intervals are shown.
Our estimate of mortality attributed to WNV infection (34.6%) was not significantly different than that reported from the experimental infections of this species by Komar et al. (2005) of 22.2% (p = 0.39). We also did not detect any decline in the local Northern Cardinal population with the advent (circa 2002) or continued WNV activity in the area (slope of 0.01 for the CBC and 0.02 for the SBC; Fig. 2).
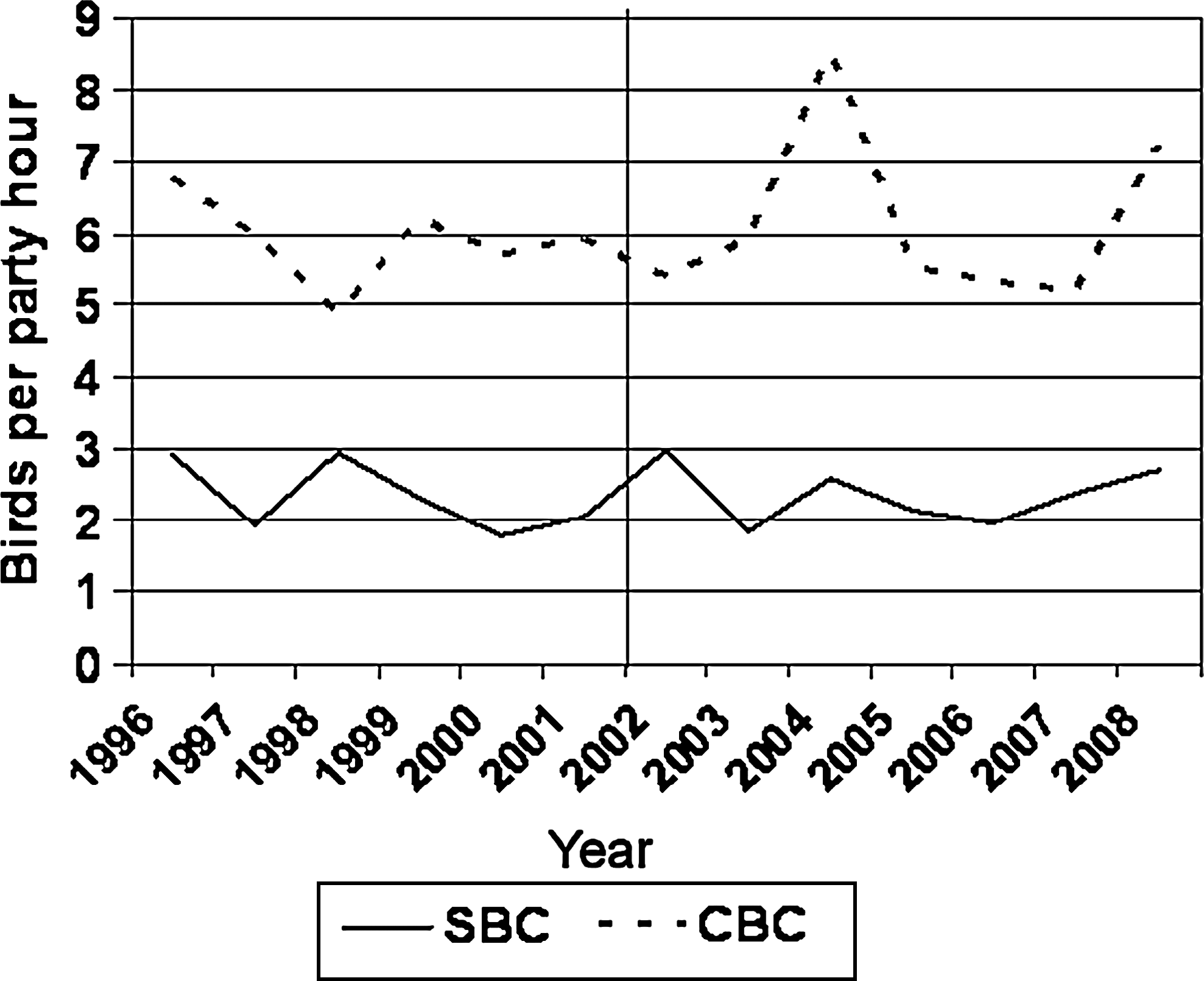
The number of Northern Cardinals per party hour during the Champaign County Spring Bird Count (SBC) and the National Audubon's Christmas Bird Count (CBC) conducted in Champaign County from 1996 to 2008. The line represents the arrival of WNV in the area.
Discussion
Our results suggest that WNV affects the survival of free-ranging Northern Cardinals. Support for the season*anti model indicates differences in survival between birds with and without WNV antibodies in both the WNV season and during the remainder of the year. Although the model suggests differences in both time periods, difference in apparent survival was much greater during the WNV season. Individuals that have been infected with WNV and survive, develop antibodies and have a relatively high rate of survival during the WNV season, whereas susceptible individuals have a relatively low survival rate. Although the survival rate of Northern Cardinals without WNV antibodies was low in the WNV season, it was still not as low (though the 95% CI do overlap) as it was in the winter. This suggests that survival is lowest in the non-WNV season, probably due to winter mortality (Newton 1998). We also did not detect any difference in WNV-related mortality between this study and the experimental inoculations by Komar et al. (2005), suggesting that the experimental inoculations may produce mortality estimates similar to what would be observed in the field.
The difference in apparent survival between the antibody-positive and antibody-negative individuals may reflect the percentage of the population that succumb to WNV. This is assuming there is no difference in dispersal rates and non-WNV-related mortality. The recapture probability varied over time, likely because in winter we had a higher recapture rate as birds were moving between feeding locations, which resulted in the concentration of birds near our nets and traps. During the breeding season the cardinals were dispersed across the area and we generally caught fewer individuals.
A comparison of the apparent survival rate of antibody-positive individuals (45.5%) in this study to other studies suggest antibody-positive individuals may have a slightly lower survival rate when compared with what has been found in other studies. A similar study in Maryland before the arrival of WNV estimated annual survival for cardinals at 60% (SE 6.0%) (Karr et al. 1990), and a study in southern Ontario found that 65% of breeding birds returned between years (Scott 1967). This finding of slightly lower survival than other species before the arrival of WNV, and the result of the survival analysis suggest the possibility of chronic effects from WNV. Although the magnitude of differences in the apparent survival rates for the non-WNV season is much smaller than during the WNV season, the difference is consistent with chronic effects. These results are also consistent with Kilpatrick (2003), who found that individuals that survive infection of malaria had lower survival rates, and Reisen et al. (2006) have found that birds can be chronically infected with WNV. A longer-term study with a larger sample size would be needed to adequately address the chronic effects of WNV in birds. If chronic effects persisted after the WNV season, given that winter mortality may be the bottleneck for resident bird populations (Newton 1998), it is likely that these lower conditioned birds would quickly be removed from the population, so the effect may be most pronounced in early winter.
Although substantial WNV-related mortality is occurring in our study area, WNV does not appear to be impacting the local population. This is in agreement with LaDeau et al. (2007), who estimated population trajectories of Northern Cardinals before and after the arrival of WNV in Illinois using the Breeding Bird Survey data. They found that the Northern Cardinals were exhibiting no decline, and there were no clear impacts from WNV in this population. This was true for Northern Cardinals not only in Illinois, but also in all areas they analyzed. The finding that Northern Cardinal populations appear to be unaffected by WNV, while WNV has an impact on survival rates, suggests that Northern Cardinal populations are able to compensate for the WNV-related morality. Komar et al. (2005) estimated that WNV may have had a small impact (12% decline based on formulas associated with WNV-related mortality, population size, and seroprevalence rate) on a local Northern Cardinal population in Slidell, Louisiana. Although there are likely many reasons why some species are impacted while others are not, this study suggests that, while mortality in Northern Cardinals may increase with the presence of WNV, the population appears to be resilient to this disease.
Footnotes
Acknowledgments
We thank M. Alessi, M. Avara, M. Barron, L. Condon, and J. Horn for assistance in the field, and Drs. Pat Weatherhead, Thomas Benson, and an anonymous reviewer for comments on the article. This research was conducted under Animal Protocol #04010. This work was funded by the Illinois Waste Tire Act Fund and USDA/CREES 2005-34523-15639.
Disclosure Statement
No competing financial interests exist.