
Abstract
Emerging infectious diseases are recognized as increasing threats to public and animal health, global economy, and social and political stability. Climate change, environmental changes, changes in human demographics and behaviors, and the rise of global trade and travel are most-often-cited drivers for the emergence of infectious diseases in human and animal populations (Schrag and Wiener 1995, Daszak et al. 2001, Dobson and Foufopoulos 2001, May et al. 2001, Taylor et al. 2001, Antia et al. 2003, IOM 2003, Kuiken et al. 2003, Weiss and McMichael 2004, Wolfe et al. 2005, Woolhouse and Gowtage-Sequeria 2005, Chomel et al. 2007, Woolhouse and Gaunt 2007, Jones et al. 2008). Emerging pathogens are more likely to be zoonotic or vector-borne with a broad host range (Daszak et al. 2000, Taylor et al. 2001, Woolhouse and Gowtage-Sequeria 2005, Jones et al. 2008). Emerging pathogens are also more frequently RNA viruses (Woolhouse and Gowtage-Sequeria 2005), which may better adapt to and establish in novel host species (Antia et al. 2003, Andre and Day 2005, Woolhouse et al. 2005). Determining the factors driving disease emergence eventually aims at assisting the prediction of the future emergence of infectious diseases. However, because of the multifactorial nature of the drivers involved (IOM 2003), our ability to predict which pathogens may arise in human or animal populations in the future remains limited (WHO/FAO/OIE 2004). In particular, the ability to identify the animal reservoirs where the greatest risks to human health will originate was deemed improbable (WHO/FAO/OIE 2004). Applying the theory of island biogeography (MacArthur and Wilson 1967) to emerging pathogens identifies (i) interactions between recipient host species and species sources of pathogens (including vector species), (ii) interactions within species sources of pathogens, and (iii) interactions within recipient host species, as mechanisms directly driving disease emergence. Most identified drivers for disease emergence correspond to changes in one or more of these three sets of interactions. Importantly, applying the theory of island biogeography to emerging pathogens assists in predicting from which animal species future zoonotic and vector-borne pathogens will most probably emerge. Identifying known and unknown pathogens such species harbor may bring us one step forward toward predicting which pathogen will next emerge in human populations.
Hosts can be considered as islands for pathogens (Kuris et al. 1980)—islands that can successfully be colonized provided that introduced pathogens are able to establish (cause infection) and avoid extinction (transmit between hosts) (May et al. 2001). The analogy of hosts as islands for pathogens was proposed after MacArthur and Wilson's theory of island biogeography, a landmark contribution to ecology (MacArthur and Wilson 1967). Biogeography is the study of the distribution of biodiversity, which can be measured by species richness (number of species), over space and time. MacArthur and Wilson (1967) proposed that a biotic equilibrium of the number of species on a given island is reached when the immigration and extinction rates (species/unit time) equal each other (Fig. 1). Immigration rates fall and extinction rates rise with the number of species present on the island. Immigration rates also fall when the degree of isolation from the source regions that supply the species (source-island distance) is increased; extinction rates also rise when the area of the island is decreased (Fig. 1). Likewise, it was proposed that parasite species richness results from the balance of rates of parasite colonization and extinction (Poulin 2004) (Fig. 1). Source-island distance–like factors may influence the rate of parasite colonization, and relate to interactions between sources of parasites and recipient hosts, mostly through spatial or behavioral overlap (e.g., host habitat and geographical ranges, and diet). Island area–like factors may influence the rate of parasite extinction, and include species–area relationships (through host body size and lifespan) but also relate to intraspecific interactions of the newly colonized hosts (through population density and social behavior) (Kuris et al. 1980) (for a review, see Poulin, 2004).

Equilibrium models of species richness on islands (black and bold text) and of parasite species richness within host species (gray and nonbold text). S, number of species at equilibrium; X, species turnover rate at equilibrium. Adapted from MacArthur and Wilson (1967).
Changes in source-island distance–like factors and island area–like factors (i.e., changes in the degree of interactions between recipient host species and species source of pathogens, and changes in the degree of interactions within recipient host species) resulting in changes in rates of pathogen colonization and/or extinction may explain the increase in the number of novel pathogens in human (or animal) populations (Fig. 2). Indeed, most identified drivers for disease emergence in human populations correspond to changes in one or both sets of interactions (Table 1). Thus, economic development and the rise of bushmeat hunting in Africa and Asia, or changes in land use and the conversion of natural habitats into agricultural lands in South America, have reduced source-island distances between animal species sources of pathogens and humans, and contributed to the introduction of emerging pathogens into human populations, for example, human immunodeficiency viruses (HIV), filoviruses, monkeypox virus, Severe Acute Respitory Syndrome (SARS) coronavirus, or New World arenaviruses (Charrel and Lamballerie 2003, Wolfe et al. 2005, Chomel et al. 2007). Urbanization, global travel, and changes in human demographics and behaviors have facilitated contact between humans, and increased island area, contributing to the establishment and spread of emerging pathogens, for example, urban strains of dengue and yellow fever viruses, HIV, or SARS coronavirus (Yuill 1986, May et al. 2001, Anderson et al. 2004).
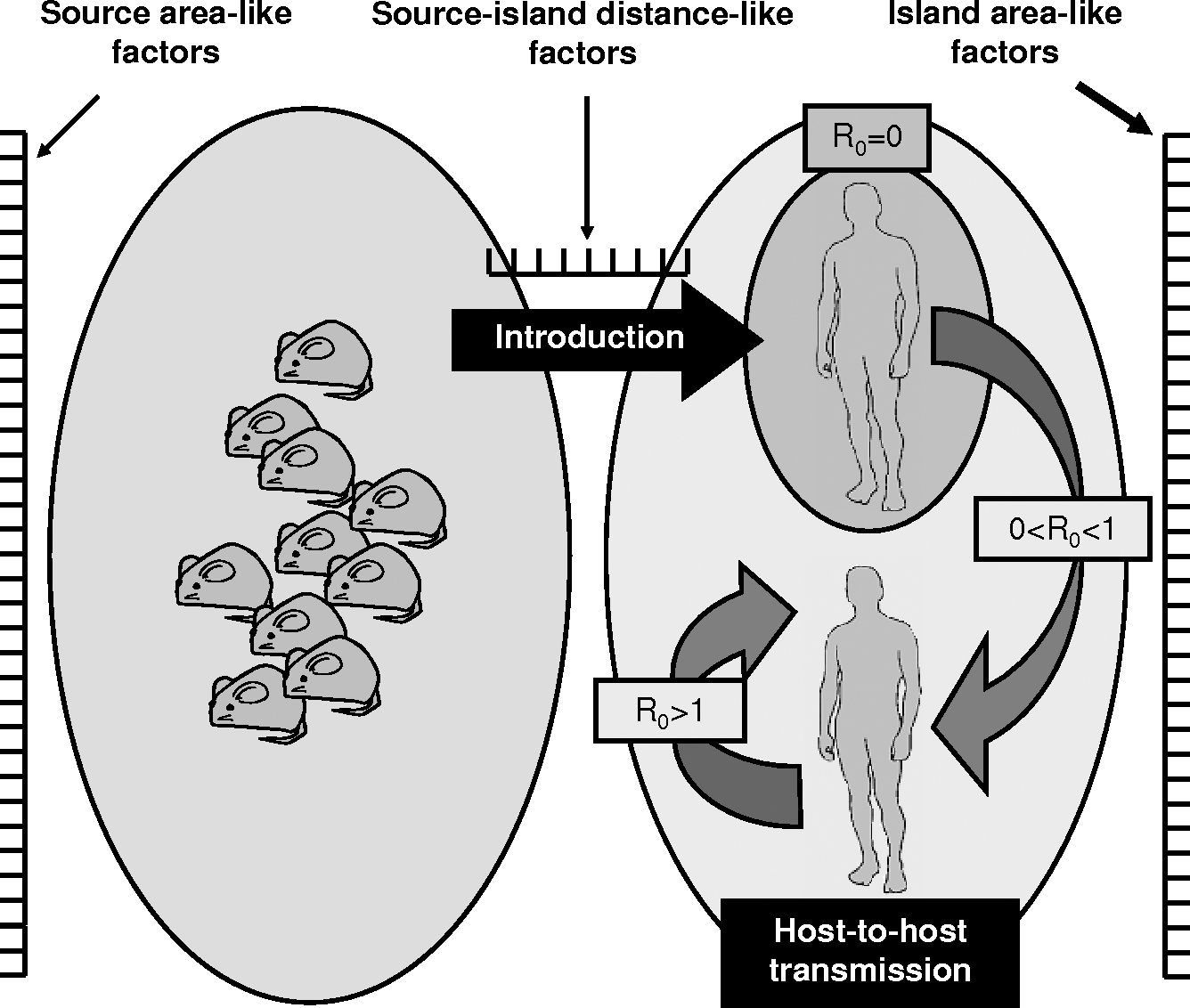
Illustration of the process of disease emergence based on the theory of island biogeography applied to the transmission of pathogens to novel host species.
Importantly, MacArthur and Wilson (1967) demonstrated that source area and distance to the recipient island were also major source factors influencing the rate of propagules leaving the source regions and the probability for these propagules to reach the recipient island, respectively. Changes in source-area–like factors and source-island distance–like factors (i.e., changes in the degree of interactions within species sources of pathogens, and changes in the degree of interactions between species sources of pathogens and recipient host species) are thus likely to influence the rate and probability of successful cross-species transmission of pathogens (Fig. 2). Accordingly, the rise of agriculture and domestication, around 10,000 years ago, resulted in dense animal populations (large source area) in close contact with humans (short distance to recipient island) that contributed to the emergence of most pathogens now long restricted to the human species (Wolfe et al. 2007). Likewise, destruction of natural habitats in Australasia has led fruit-eating bats to colonize anthropogenic environments, reducing source-island distances between bats and humans, and favoring the emergence of Australian bat lyssavirus, menangle virus, and henipaviruses (Field et al. 2001). In addition, animal species that exploit anthropogenic environments (reducing source-island distances) typically reach higher densities than in natural environments (McKinney 2002), resulting in an increase in source area. For example, colonization of residential areas by white-tailed deer (Odocoileus virginianus) in the eastern United States is accompanied by an increase in their density and the density of tick vectors, and is associated with the emergence of Borrelia burgdorferi (Barbour and Fish 1993). Climate change or global trade also may result in expanding populations and/or increasing densities of source species, contributing to the emergence of pathogens; for example, Sin Nombre virus associated with higher density of its rodent reservoir due to El Nino events (Engelthaler et al. 1999) and European tick-borne encephalitis virus associated with increasing populations of tick vectors partly due to rising temperatures (Randolph 2001), or monkeypox virus in the United States associated with the importation of exotic pets (CDC 2003) and yellow fever virus and other vector-borne pathogens transmitted by Aedes spp. that are disseminated around the world via shipping (IOM 2003) (Table 1).
MacArthur and Wilson (1967) also demonstrated that the presence of one stepping-stone connecting the source region to a recipient island can significantly enhance immigration to the recipient island, provided that the stepping-stone is able to support populations of the immigrating species in the first place. The larger and closer is the stepping-stone to the recipient island, the greater is the increase in the immigration rate. Domestic animals have been implicated as relay species in the transmission of zoonotic pathogens from wildlife species to humans, for example, horses and swine and transmission of menangle virus and henipaviruses originating from fruit bats (Field et al. 2001), or swine and poultry and transmission of avian influenza viruses (Webster and Hulse 2004). Because of close contact with both humans and wildlife species, and increasing populations worldwide, domestic animals are likely to be major stepping-stone species favoring the emergence of infectious diseases in humans. Wildlife species also have played the role of relay in the cross-species transmission of pathogens, for example, primates and transmission of filoviruses (Wolfe et al. 2005), prairie dogs (Cynomys spp.) and transmission of monkeypox virus (CDC 2003), or palm civets (Paguma spp.) and transmission of SARS coronavirus (Chomel et al. 2007).
Surveillance of animal populations and identification of known and unknown pathogens they harbor are urged in our efforts to mitigate the effects of infectious diseases that will emerge in the future (Daszak et al. 2000, 2001, Dobson 2005, Kuiken et al. 2005, Jones et al. 2008). However, animal populations to be targeted by such surveillance efforts are rarely clearly specified. Applying the theory of island biogeography (MacArthur and Wilson 1967) to emerging pathogens identifies (i) interactions between recipient host species and species sources of pathogens (including vector species), (ii) interactions within species sources of pathogens, and (iii) interactions within recipient host species as mechanisms directly driving disease emergence. Most identified drivers for disease emergence correspond to changes in one or more of these three sets of interactions. This highlights the role not only changes in the ways recipient host species, interact with their environment and with each other, but also changes in the ways species sources of pathogens interact with their environment and with each other, play in the emergence of infectious diseases (Fig. 2). Therefore, animal species, including vector species, associated with humans and/or with expanding or growing populations at the interface of natural and human habitats (short source-island distances and large source area) should be targeted by active surveillance efforts. These species include mostly (i) domestic and laboratory animals, (ii) animals from pet trade and meat markets, (iii) the so-called pest species and managed game species, which dwell in cultures and recreational areas, and (iv) the so-called urban adapters and exploiters (McKinney 2002), which dwell in urbanized environments. A scaling system designed to assess the extent of the three sets of interactions within and between candidate source species (including vector species) and humans may help identify species with highest risk for cross-species transmission of pathogens. These species are most likely to be the sources of future emerging vector-borne and zoonotic diseases.
Footnotes
Acknowledgments
I warmly thank Andy Dobson, Penelope Koraka, Thijs Kuiken, Byron Martina, and Ab Osterhaus for fruitful discussions and useful input.
Disclosure Statement
No competing financial interests exist.