
Abstract
A total of 196 small mammals were collected in Portugal and tested for Borrelia burgdorferi sensu lato. Tissue samples were taken from each animal and cultured in Barbour-Stoenner-Kelly (BSK)-II medium. The single strain of spirochete isolated was confirmed as Borrelia lusitaniae by genetic analyses. This is the first report of B. lusitaniae isolated from Apodemus sylvaticus.
Introduction
Different Borrelia genospecies are associated with distinct ecologies and enzootic cycles, specific pathogenicity, and clinical symptomatology in patients. The list of reservoirs for B. burgdorferi s.l. in endemic areas of Europe is extensive. Some associations, however, between Borrelia genospecies and vertebrate hosts have been identified, such as B. garinii and B. valaisiana in birds, B. afzelii within small rodents, and B. burgdorferi s.s. in red squirrels and hedgehogs (Humair and Gern 1998); lizards may be the principal reservoir hosts of B. lusitaniae (Richter and Matuschka 2006, Amore et al. 2007). Although B. lusitaniae circulates between a range of ticks and host vertebrate species such as lizards, birds (Poupon et al. 2006), and small mammals, the vertebrate reservoir of B. lusitaniae had yet to be identified. In a study performed in the Grandola region, B. lusitaniae DNA was detected by polymerase chain reaction in all small mammal species captured (Mus spretus, Apodemus sylvaticus, Rattus norvegicus, and Crocidura russula), but it was not possible to isolate spirochetes. These observations indicate that there may exist a sylvatic cycle specific to this Borrelia genospecies, maintained by a variety of hosts and reservoirs (Baptista 2006). In Portugal, to address this question we initiated a study to determine whether small mammals can contribute to the maintenance of B. lusitaniae in natural foci in Portugal.
Materials and Methods
One hundred ninety-six small mammals (22 A. sylvaticus, 160 M. spretus, and 14 Rattus rattus) were captured using Sherman traps, between July 2002 and October 2004 in three different National Parks of Portugal: Vale Guadiana (W 8°7′19′′, N 41°47′19′′), Peneda Gerês (W 9°2′13′′, N 38°27′28′′), and Arrábida (W 7°39′50′′, N 37°41′21′′).
The rodents were brought to the laboratory and identified by external morphology and skull features. Animals were then anesthetized with ketamine hydrochloride (Imalgene; Merial, Lyon, France) at a dose of 10 mg/kg, bled by cardiac puncture for serodiagnostics, and euthanized by exposure to CO2 gas. Serum samples were then tested for the presence of immunoglobulin G antibodies against Borrelia by indirect immunofluorescence assay (Doby et al. 1991), using a strain of B. garinii as antigen. Positive serology indicated by a titer of 32.
Target organs (skin, bladder, and heart) were aseptically harvested for culture.
The organs were first disinfected by successive immersion in iodine, 70% ethanol, and phosphate-buffered saline; tissues were then minced and placed directly into 8 mL of Barbour-Stoenner-Kelly (BSK)-II medium. All cultures were maintained at 34°C for 3 months and examined weekly by dark-field microscopy to monitor the presence of live spirochetes. Heart tissue from a single A. sylvaticus mouse captured in Vale Guadiana demonstrated viable spirochetes 8 weeks after culture in BSK-II medium; 1 mL of culture was centrifuged for 5 min at 7000 g, and the sediment washed twice with sterile phosphate-buffered saline and frozen at −20°C until DNA extraction. Total DNA was extracted using the QIAamp tissue kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Three genes, flaB (9), rrf (5S)-rrl and (23S) intergenic spacer region of B. burgdorferi s.l. (Johnson et al. 1992, Rijpkema et al. 1995), and the outer surface protein A (OspA) gene, were used to characterize this isolate (Lopes de Carvalho et al. 2008a). The sequences were assembled by combining the sequences generated by each primer, using the BioEdit software. For phylogenetic inference the alignments were made using amino acid sequences and converted to DNA sequences using BioEdit software. All alignments were made using ClustalX program (Thompson et al. 1997) and manually inspected for misalignments. Primer sequences except for the last two (5′) to six (3′) nucleotides were removed from the alignment before phylogenetic analyses, regarding the polymorphisms observed in the remaining sequences. Neighbor-joining tree of DNA sequence alignment was conducted in PAUP* 4.0b10 software. Distance matrices were calculated using the Kimura two-parameter model to correct for multiple substitutions. Bootstrap analysis was obtained with 1000 replicates.
Results and Discussion
Of the 169 small mammals captured, 17 (10.6%) M. spretus and 1 (5.6%) R. rattus showed antibodies to B. lusitaniae; however, no positive cultures were obtained from these host species. Despite isolation of spirochetes from one A. sylvaticus captured in the Vale Guadiana National Park, no seropositive samples were obtained from this species. The flaB gene, the rrf (5S)–rrl (23S) intergenic spacer region of B. burgdorferi s.l., and the OspA gene were amplified from DNA extracted from the A. sylvaticus isolate, and further analysis of this isolate identified it as B. lusitaniae. Phylogenetic analyses based on the OspA gene grouped this new isolate near other B. lusitaniae isolated in Portugal and most closely aligned to the North African clade (Grego et al. 2007) (Fig. 1). OspA clustering via the Neighbor-joining algorithm was confirmed by Maximum Likelihood (ML) and MrBayes analysis (data not shown). Intergenic Spacer (IGS) sequences also confirmed the PoAnB1 strain as B. lusitaniae as the mean intra–B. lusitaniae group sequence identities vary between 95.8% and 91.6% and are below 90% with other Borrelia genospecies (89.8% with B. garinii, 86.9% with B. afzelii, 86.5% with B. valaisiana, and 85.1% with B. burgdorferi s.s.). The eco-epidemiological studies performed to date demonstrate that B. lusitaniae presents different characteristics to existing B. burgdorferi genospecies in other European countries (Zeidner et al. 2002, Grego et al. 2007). Previous serology results (Núncio 2002) as well as the DNA detection (Baptista 2006) and the isolation and molecular identification of B. lusitaniae indicate that small mammals, particularly, A. sylvaticus, may play a role in the maintenance of B. lusitaniae in Portugal. To our knowledge, this is the first report of live B. lusitaniae spirochetes isolated from A. sylvaticus, an indication that this mammal is a competent reservoir of B. lusitaniae.
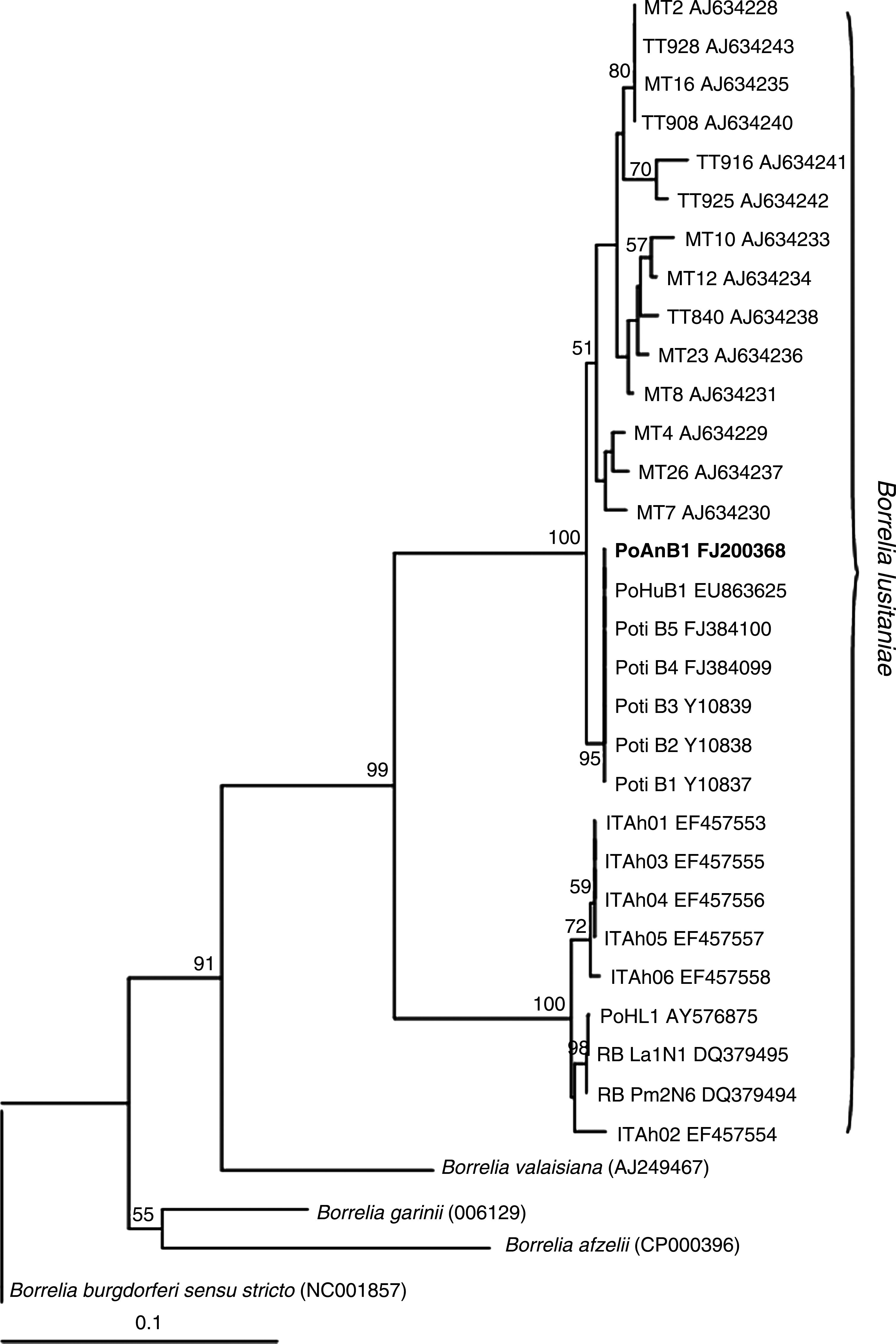
Neighbor-joining tree inferred from partial OspA gene sequences from the isolate obtained in this study (bold) compared to GenBank sequence data. Distance matrices were calculated using the Kimura 2-parameter model to correct for multiple substitutions. Bootstrap values were obtained from 1000 replicate trees and are indicated at the nodes (>50%). Po, Portuguese strains; ITAh, Italian strains; MT, Morocco strains; TT, Tunisia strains; RB, German strains.
Previous studies have demonstrated that B. lusitaniae is not as immunogenic as B. burgdorferi s.s. (Zeidner et al. 2002). Even in human cases, isolates derived by culture were achieved in a seronegative and in a borderline seropositive patient (Collares-Pereira et al. 2004, Lopes de Carvalho et al. 2008a). Thus, the low titer (Ig = 16) antibody detected in the A. sylvaticus mouse from which the borrelia was isolated seems to confirm a pattern regarding B. lusitaniae. Future studies, using an autochthonous borrelia strain as antigen, should improve the sensitivity of our serological testing.
Despite evidence in several other studies indicating lizards as main reservoir host of B. lusitanie in some geographical areas (Richter and Matuschka 2006, Amore et al. 2007), our findings demonstrate that more attention be given to small mammals as potential reservoirs. In addition, parallel studies in birds and lizards are currently ongoing to gain a more detailed assessment of both the ecology and pathogenicity of borrelia strains circulating in Portugal.
Nucleotide sequence accession number: The rrf (5S)-rrl (23S) intergenic spacer region of B. burgdorferi s.l., flaB, and OspA gene from PoAnB1 have been deposited in GenBank with accession numbers EU647595, EU122385, and FJ200368, respectively.
Footnotes
Acknowledgments
The authors wish to thank Mary Crabtree, Division of Vector-Borne Infectious Diseases, Centers for Disease Control and Prevention, Fort Collins, CO, for the help with the phylogenetic analysis. Teresa Luz, Centro de Estudos de Vectores e Doenças Infecciosas, is also acknowledged for her expert technical assistance. This study was partially supported by FLAD with a fellowship and FCT project (POCTI/ESP/39549/2001).
Disclosure Statement
No competing financial interests exist.