
Abstract
Chagas disease is prevalent in Peru. The province of Nazca, in the southwestern region of the country, shows a high intradomiciliary infestation rate of Triatoma infestans bugs. Although the vector is present, the number of Chagas disease cases appears to be much lower than those reported in the neighboring region of Arequipa. We examined 624 T. infestans from Nazca to determine the current Trypanosoma cruzi infection rates, and found that no bugs were infected with this parasite. These results contrast with those found in Arequipa, where 19–30% triatomines have been reported infected. To compare their vectorial capacity, we infected 30 T. infestans specimens, selected both from Nazca and Arequipa, by feeding bugs on T. cruzi–infected mice. The parasites developed all stages expected in the vector; furthermore, the infective stage, metacyclic trypomastigote, was found in both insect populations from the second week after infection. In addition, those insects that accepted to be fed with mice blood defecated immediately after finishing blood meal, indicating that they might be efficient vectors. We maintain that differences observed in infection rates between vectors from Nazca and Arequipa may be explained by differences in host availability. In Arequipa hosts are mainly small animals, whereas in Nazca the main blood source comes from birds, which are refractory to the infection.
Introduction
Chagas disease is transmitted to humans when insect feces containing T. cruzi trypomastigotes enter the host open skin or mucous membranes. The vectors, some species of the family Reduviidae, become infected by feeding blood from infected hosts containing T. cruzi trypomastigotes (Zeledon and Rabinovich 1981). In the intestinal tract of the bugs the parasites undergo several transformations. It is possible to identify spheromastigotes, epimastigotes, and trypomastigotes as the most representative stages, in addition to the intermediate forms described by Kollien and Schaub (1998a). In Peru, the main reservoirs of the disease in domestic and peridomestic environments are humans, cats, dogs, rodents, and guinea pigs (Mendoza Ticona et al. 2005).
In the southwestern region, Chagas disease has also been reported as endemic in the departments of Ica, Moquegua, and Tacna (Cornejo del Carpio 2003); however, information for these departments is scant. Triatomine infestation has been reported since early studies in Nazca (Ayulo-Robles 1946), but scarce information about the triatomines infection rates appear to be available for this region. Solis Acosta et al. (1997) reported that 20.7% of the population evaluated from Nazca, the southernmost province of the Ica department, was seropositive to Chagas disease. This study had two main objectives: the first is to investigate the natural rate of infection (tripano-triatomine index) in Nazca, and the second is to determine the vector efficiency of the T. infestans populations that exist there. The presence of trypomastigotes in the insect feces and the defecation patterns of the vector were used to evaluate the vector efficiency (Zeledon et al. 1977, Kirk and Schofield 1987, Trumper and Gorla 1991, Crocco and Catala 1996, Rodriguez et al. 2008). We are hoping to contribute to the knowledge of the transmission of the Chagas disease in the province of Nazca, to date largely unknown.
Materials and Methods
Collections
The province of Nazca, or Nasca, is one of the five provinces of the department of Ica. It is located in the southwestern region of Peru, bordering with the Arequipa department in the south, Ica and Palpa in the north, Ayacucho in the east, and the Pacific Ocean in the west (Fig. 1). The area is arid with scarce precipitations. Mining, agriculture, animal farming, and tourism are among the most economically important activities of the Nazca inhabitants (Solis Acosta et al. 1997). Entomological surveys took place on August 2003 and November 2005, in the districts Vista Alegre and Nazca, in the province of Nazca (Fig. 1). We only visited triatomine-infested houses, selected using information from prior surveys carried out by staff from the Ministry of Health in Nazca.
Map of Peru showing collection sites in the province of Nazca, obtained from the Institute of Statistics and Informatics of Peru (
Domestic and peridomestic environments were explored. We followed Walter et al. (2007) definition for a peridomestic environment: the full area around the house supporting permanent or temporary structures built and used by humans or by their domestic animals. Bugs were collected from chicken coops, corrals of domestic mammals, charcoal piles, pigeon houses, palms, and dog houses. Within the houses, bugs were collected from holes in walls and ceiling. When insects were found in wall crashes and holes, they were pushed with metal sticks and forceps. Specimens collected were placed into metal cages covered with nylon tulle and then transported to the University of San Marcos (UNMSM) laboratories in Lima.
Rate of natural infection
One fecal drop was obtained by abdomen compression of bugs, placed in saline (NaCl 0.9%), and examined in the optical microscope, at 400 magnification. Flagellates were searched by examining 50 fields per specimen. Only fourth and fifth instars nymphs and adults were used, whereas the remaining stages of the insects were kept in the laboratory until they turn to fourth-stage nymphs.
T. cruzi–free insects were kept in the insectary at the UNMSM, at an 80% of relative humidity and a temperature of 28°C, and fed on chickens. Eggs obtained were raised until hatching. Only insects from first generation born in captivity were used for further experiments.
Infective capacity
Two sets of insects were used in the experiments, each comprising 30 fourth instar-nymphs of T. infestans. Set 1 comprised specimens from a colony maintained for several years at the insectary of the Institute of Tropical Medicine (IMT) from the UNMSM, collected from Arequipa department. Set 2 were fourth instar progeny broods of the specimens collected in Nazca in this study. The T. cruzi strain used in the assays was obtained from the province of Sihuas in Arequipa, an Chagasic area, and maintained by successive passages in albino mice (Swiss strain), at IMT-UNMSM.
Insects were fed from T. cruzi–infected mice on the day of the highest parasitemia, which had been previously estimated in the IMT-UNMSM (Hilda Solis, personal communication). Triatomine fecal samples were examined after 1 week, 2 weeks, 4 weeks, 2 months, and finally after 3 months. Each time, we estimated the number of flagellates observed in 50 fields and the percentage of each stage of development. The stages of the parasites observed were defined according to Kollien and Schaub (2000) as follows: spheromastigotes, epimastigotes, metacyclic trypomastigotes, “drop-like” intermediate stage, and “slender-shape” intermediate stage. Insects were fed on chicken blood every 15 days.
To estimate the defecation pattern of the insects, specimens containing metacyclic trypomastigotes were fed on noninfected mice. The periods from beginning to feed, bite interruption, total feeding time, and defecation were estimated. Triatomine bugs were infected individually.
Results
Natural rate of infection
A total of 624 T. infestans specimens were collected in two collection dates: 173 specimens on August 2003 and 451 specimens on November 2005. Insects were collected from 10 houses from Vista Alegre district and 45 from Nazca. Domestic animals were found in all 55 houses, 70.9% of the houses had birds and mammals, 18.2% birds only, and 10.9% mammals only. The microscopic examination showed that none of the insects were infected with any of the stages of T. cruzi.
Infective capacity of Trypanosoma cruzi individuals
All triatomines fed on infected mice contained parasites in their feces. The total number of parasites quantified in the five observations was somewhat similar in the two populations evaluated: 1455 parasites were counted in the 30 specimens from the Arequipa colony and 1368 in the 30 individuals from Ica. Results are given in terms of the percentage of parasites observed, since the total numbers depended on the amount of fecal sample or saline examined by the examiner and, therefore, subject to errors.
Seven days after infection (dai), the drop-like intermediate stages (between spheromastigotes and epimastigotes or trypomastigotes) dominated in both populations. These were 70% and 61.9% of the parasites observed in Ica and Arequipa, respectively. Their number decreased 2 weeks after infection (Fig. 2A). The other stages present 7 dai were epimastigotes and parasites with “slender shape.” No trypomastigotes were observed at this point. The spheromastigotes (9.09% and 11.45% of Ica and Arequipa specimens) appeared 14 dai, except a single flagellate observed in the Nazca specimens 21 dai (Fig. 2B). The metacyclic trypomastigotes were observed 14 dai in both set of insects, 7.95% in Ica and 4.11% in Arequipa specimens. Their number increased each time they were observed (Fig. 2C). On the last examination, 90 dai, trypomastigotes become the most abundant stage observed in Ica triatomines (34.77%). In contrast, in the Arequipa specimens, trypomastigotes (29.32%) were outnumbered by the drop-like parasites (33.83%). Epimastigotes were observed from the first to the last observation in both sets of specimens (Fig. 2B). We calculated the linear regression coefficient of the percentage of T. cruzi stages as a function of time (Fig. 2). Trypomastigotes were the only stage that showed a linear trend during the five observations. Regression coefficients were close to unity (0.98 and 0.94 for Ica and Arequipa specimens, respectively). We used the slope of the linear regression to define the growth rate of trypomastigotes. The differences between Ica and Arequipa trypomastigotes growth rates (Ica, 0.37 ± 0.05; Arequipa, 0.33 ± 0.03) were negligible (Fig. 2).
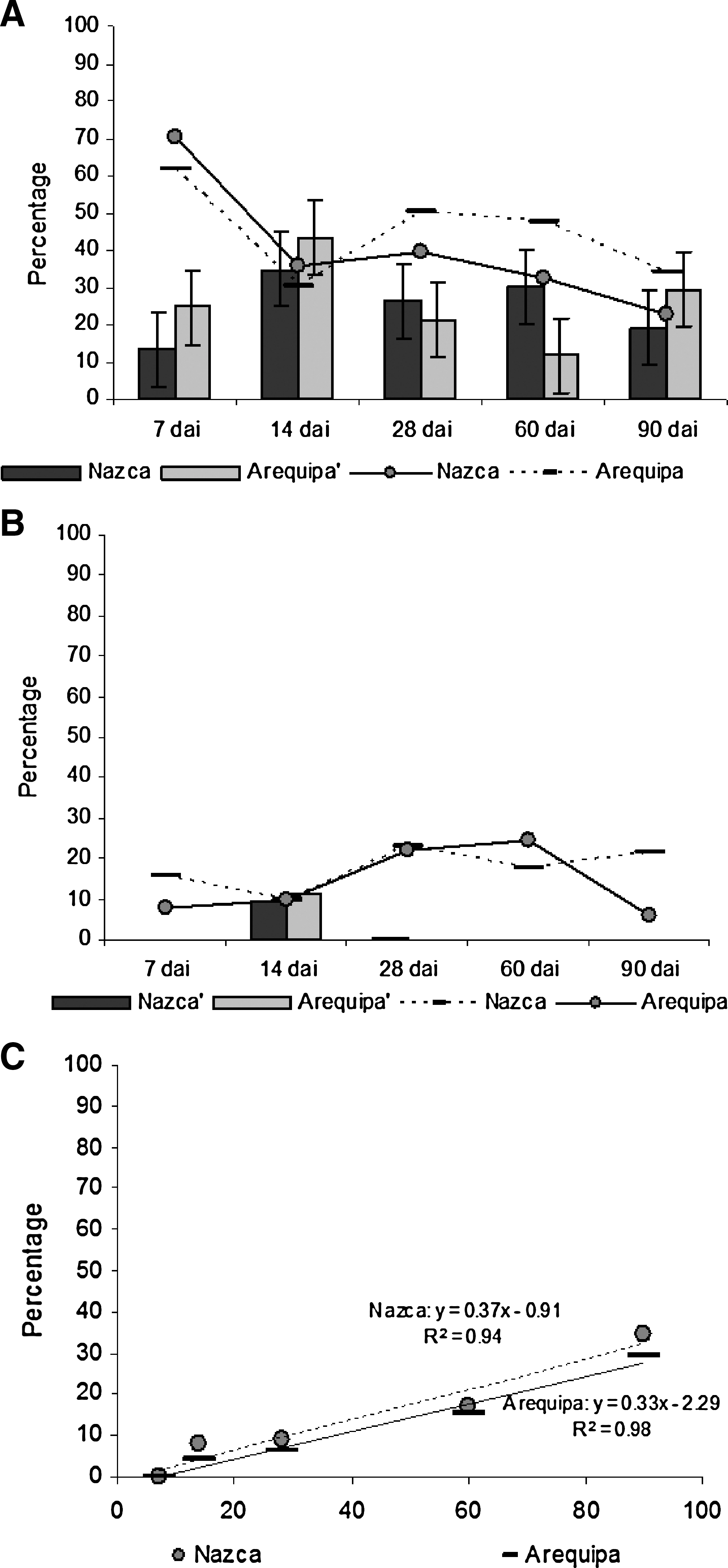
Percentage of Trypanosoma cruzi stages observed in Triatoma infestans from Ica and Arequipa. (
Defecation after feeding
It was not possible to feed all triatomine bugs on mice, since only 30% and 50% specimens from Ica and Arequipa, respectively, completed their blood intake without interruptions. These eight bugs defecated immediately after completing their meal.
From the 10 specimens from Ica used in the experiments, only 20% of the insects bite immediately; the remaining ones started between 1 and 3 min after mice blood was offered. Similarly, in Arequipa, only one single specimen (10%) started to feed immediately, whereas the remaining ones started between 1 and 8 min later. Feeding times ranged 18–67 min in specimens from Arequipa and 6–44 min in specimens from Ica (Table 1).
n, number of triatomines; M, mean; SD, standard deviation.
Discussion
For decades, the Peruvian department of Ica was considered to be having a high risk of becoming a Chagas disease–endemic area (Ayulo-Robles 1946). However, there is a lack of confirmed Chagasic patients and external evidence of the disease (such as the Romaña's sign) in the last years, except for introduced cases from Arequipa (Cabrera et al. 2002). Further, scant information on T. cruzi infection rates in the triatomines vector is also difficult to obtain from this province (Cornejo del Carpio 2003).
We found no T. infestans bugs infected by T. cruzi in the 624 specimens examined from Nazca, even though houses in this area show high infestation by triatomines. Low T. cruzi infection rates in triatomines were previously reported by Solis et al. (2003), where a single triatomine infected with T. cruzi was identified out of specimens in 494 houses surveyed in Nazca during 1997–1998. In contrast, in the neighboring department of Arequipa, Levy et al. (2006) reported that 19.3% of the 397 houses surveyed in this region harbored infected triatomines. Additionally, infection rates as high as 30% in Arequipa have been reported (Cornejo del Carpio 2003).
Our results are consistent with serological surveys carried out by the local authorities (Direccion de Salud de Ica, Health Direction of Ica, unpublished), and with the Panamerican Health Organization, which report 0% of seropositivity to Chagas in Ica inhabitants in 2000 (PAHO 2001). Nevertheless, these conflict with those of the 20.6% of seropositivity found by Solis Acosta et al. (1997) in Nazca. It is important to note that the main limitation of Solis Acosta et al. study was that they used the indirect fluorescent antibody test only, whereas current diagnosis of Chagas disease requires positive results on two or more serological tests (WHO 2000). Since serological tests are based on the detection of antibodies, they could not easily discriminate between patients with an acute or chronic phase of the disease. The reconstruction of Nazca houses with masonry material after the earthquake of 1996 may have limited the transmission of Chagas in some areas and positive results probably detected mainly chronic infections. Large epidemiological studies are required, including serological and entomological surveys, to evaluate whether reconstructed areas have lower incidence rates in Nazca. It would also be important to investigate if there are any transmission hotspots in Nazca, as reported in Arequipa (Levy et al. 2007).
T. infestans appears to be an efficient vector in all regions where it is found, and no conclusive evidence demonstrating vector refractoriness to parasite infection has been found (Zeledon and Rabinovich 1981). However, future research on the molecular interactions between strains of within the vector is required (Araujo et al. 2007). There is some evidence of co-adaptation between this parasite and its vectors, being triatomines exhibiting a higher susceptibility to their local parasites (Vallejo et al. 2003, Araujo et al. 2008). Some triatomines species have an hemolytic factor that can affect the survival of certain T. cruzi serotypes (Kollien and Schaub 2000). Taking these lines of evidence into consideration, an experimental infection of T. infestans specimens from Nazca (Ica) and Arequipa was undertaken to investigate the vectorial capacity of this species. Unexpectedly, both insect sets were capable of producing metacyclic trypomastigotes. Similarly, the Arequipa specimens showed less epimastigotes. It is likely that specimens from Arequipa may have diminished their vectorial capacity for being kept in a colony for several generations. Regardless of these differences, both insect groups were capable of completing T. cruzi cycle.
In both insect groups, a peak in total parasite numbers was observed 2 weeks after infection. These results were consistent with those obtained by De Stephani Marquez et al. (2006), who found a higher number of T. rangeli parasites 10 dai. Trypomastigotes were detected from the second observation date, 14 dai. Their number increased linearly (regression coefficients close to unity) through the days until the last examination, 2 months after infection. Trypomastigotes from both populations grew at the same rate (slope values 0.37 ± 0.05 and 0.33 ± 0.03 for Nazca and Arequipa, respectively), indicating that they had a similar grow rate (Fig. 2C). The rest of the stages showed low regression coefficients, indicating a lack of trend. These results differ from the natural infection in T. brasiliensis, which showed trypomastigotes 3 dai (Araujo et al. 2008), but are more similar to those found by Piesman and Sherlock (1985), who detected trypomastigotes of T. cruzi between 2 and 4 weeks after infection in Panstrongylus megistus. The development of spheromastigotes within the intestine of the vector has been described as an alternative route to metacyclogenesis and has been related to stress conditions, such as starvation (Kollien and Schaub 2000). However, spheromastigotes were found 2 weeks after infection in both groups of bugs, which had not been subject to starvation. Similarly, prolonged starvation has been linked to the increase in the number of drop-like intermediate stages (Kollien and Schaub 1998b, 2000), but we found them as the dominant stage in both groups during the insect fecal examinations. It is then likely that the specimens were undergoing certain form of stress as a result of manipulation during experiments. It is also possible that these stages are observed under normal circumstances. The limited information in this respect did not allow us to arrive to further conclusions.
Zeledon et al. (1977) maintained that triatomines that defecate before 10 min postfeeding are potentially effective transmitters of T. cruzi. In this study we showed that specimens from Nazca defecate immediately after feeding when completing their blood intake, demonstrating that they are efficient vectors. However, a proportion of the insects did not accepted the blood meal on mice immediately. This may be explained by the stress caused on insects during experiments. We considered the possibility that insects developed preference for bird blood as they had been previously fed on chickens in the insectary. However, this seems unlikely according to recent studies (Gurtler et al. 2009).
Feeding preferences may have affected the role of T. infestans populations as vectors of Chagas disease in Nazca. Solis et al. (2003) showed that triatomines in this province had strong preference for avian blood. Using the precipitin test, authors observed that 62.8% of bugs collected from Nazca (reactive to any type of blood) fed on birds only. Houses surveyed in Nazca had indoor environments reconstructed with masonry material after the earthquake in 1996. However, peridomestic environments where chicken dwellings are located are still constructed with adobe materials, favoring triatomines colonization. Since host proximity may be the main reason for the eclectic host-feeding patterns in T. infestans (Minter 1976), birds accessibility in Nazca may explain the results of Solis et al. (2003). Taking into account that birds are refractory to T. cruzi infection (Teixeira et al. 2006), this may also explain the low infection rates found in this area. Conversely, Arequipa guinea pigs, which are important reservoir of T. cruzi, are consumed as typical food in the Andes area. Levy et al. (2006) showed that enclosures with guinea pigs had an average of 33.9 triatomines, which is 1.69 times more likely to keep triatomines than other animal dwellings. We suggest that host availability is the most plausible explanation for the differences observed in T. infestans infection rates in the Peruvian regions of Nazca and Arequipa.
Particular care must be taken on winter seasons, as triatomines may prefer to colonize warmer indoor environments. As vector control for Chagas disease requires effective measures that reduce expenses and increase efficiency, budgetary limitations currently restrict insect control and monitoring programs, and thus disease control is considered insufficient in Peru (Cornejo del Carpio 2003). This can only be remedied by encouraging research that improves our understanding of vector biology, including feeding preferences, dwelling preferences, and seasonal variability. We recommend that insecticide spraying to control T. infestans in Nazca should be carried out before the winter season, when the infected triatomines have a higher risk of transmitting the disease to humans, and while insects may prefer to stay in the peridomestic environments instead of seeking shelter and warmth in human dwellings.
Footnotes
Acknowledgments
We would like to thank Mirtha Huaripaucar and Mónica Guevara from the Dirección de Salud de Ica for helping us with the collection of triatomines. We also thank Ling Chipana, Mónica Saavedra, and Miguel Penagos (UNMSM) for their help, and Prof. Ines Gárate and Prof. Azucena Naupay (UNMSM) for providing us with laboratory facilities. Finally, we are very grateful to Lee Haines, Dee Walshe, and James Nash for their valuable comments on the manuscript, and to Luis Laglera for his help with the regression analysis.
Disclosure Statement
No competing financial interests exist.