
Abstract
Since the 1993 discovery of a highly pathogenic hantavirus associated with the North American deer mouse (Peromyscus maniculatus), intensive ecological studies have led to many advances in our understanding of the natural history of New World hantaviruses as it relates to human disease. Seventeen named hantaviruses have been identified in North America. Field and laboratory studies of Sin Nombre and other hantaviruses have delineated host associations, geographical distributions, mechanisms of transmission, temporal infection dynamics of these viruses in host populations, and environmental factors that influence these dynamics. Using data from these studies, preliminary predictive models of the risk of hantavirus infection to humans have been developed. Improved models using satellite-derived data are under development. Multidisciplinary collaboration, integration of field and laboratory studies, and establishment and maintenance of long-term monitoring studies will be critical to continued advancement in the understanding of hantavirus–host ecology and disease prevention in humans.
Introduction
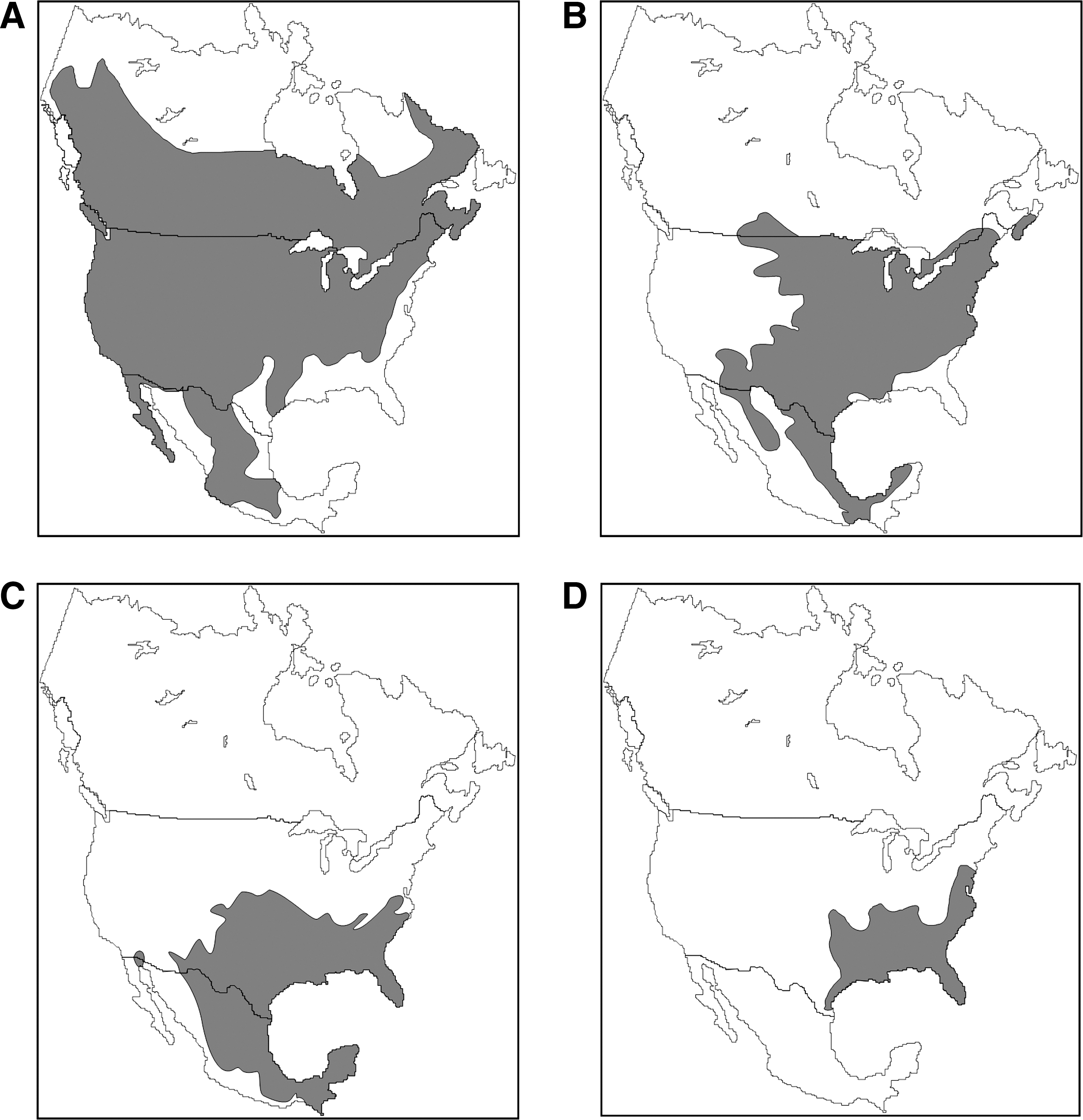
North American geographic ranges of rodents of four species of hantavirus reservoir hosts that carry hantaviruses known to be human pathogens. (
Virus nomenclature follows the 8th report of the International Committee on the Taxonomy of Viruses (Nichol et al. 2005).
Host nomenclature follows Musser and Carleton (2005).
HPS, hantavirus pulmonary syndrome.
Most of the >480 confirmed cases of HPS in 30 U.S. states (Fig. 2) are attributed to SNV, and the majority have occurred in the southwestern United States (Centers for Disease Control and Prevention [CDC], Special Pathogens Branch, unpublished data). Seventy-three cases were reported in Canada as of December 2008 (Webster et al. 2007; H. Artsob, personal communication, 2008); no HPS cases have been reported in Mexico.
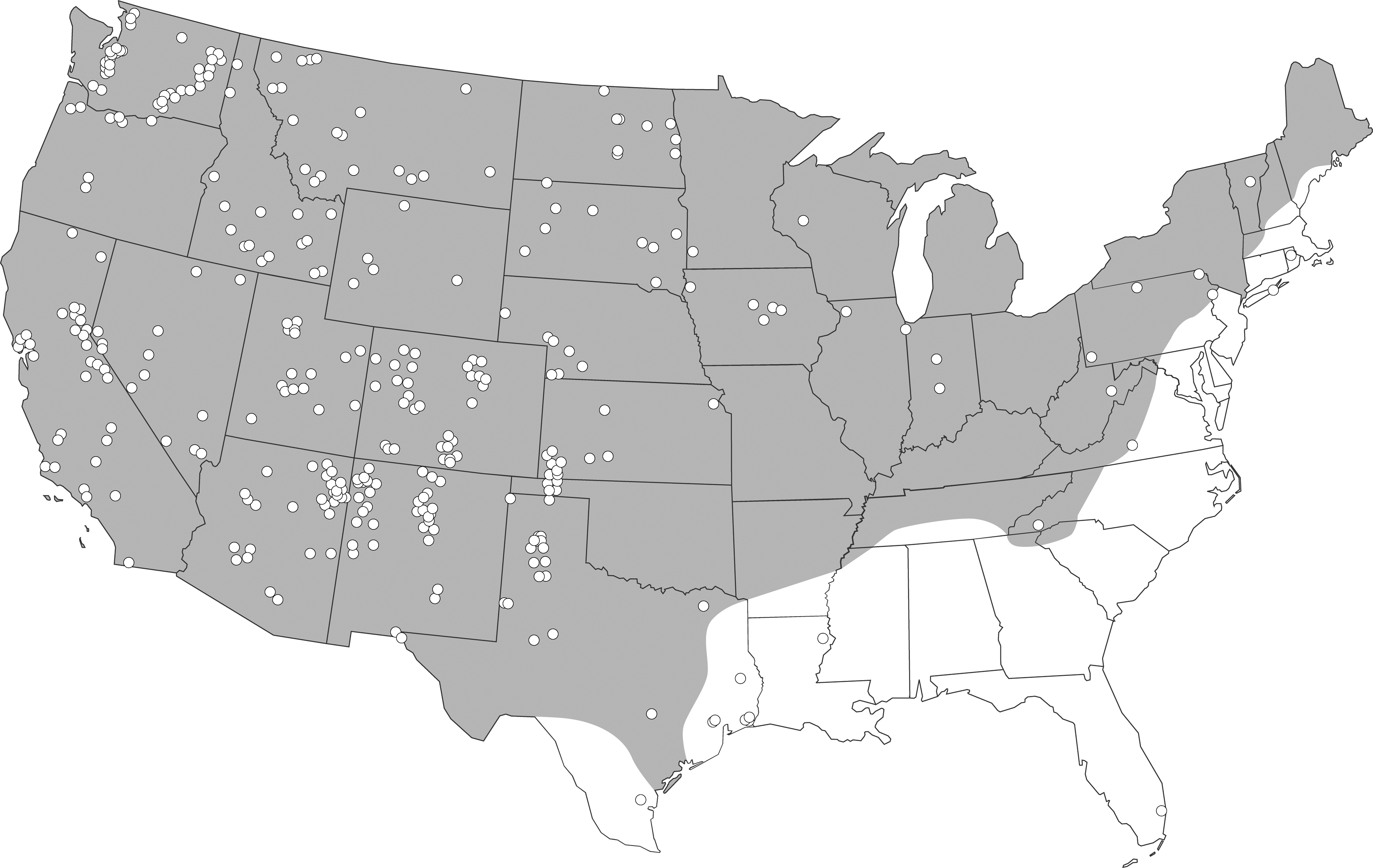
White dots represent approximate sites of exposure for 480 cases of hantavirus pulmonary syndrome (HPS) confirmed in the United States (1993–2008; Centers for Disease Control and Prevention, Special Pathogens Branch, unpublished data). Shaded area represents the range of the deer mouse (P. maniculatus) in the United States. Cases outside the range of the deer mouse were associated with hantaviruses carried by rodents of other species (Fig. 1). For example, the HPS case in southern Florida in 1994 was caused by Black Creek Canal virus, which is principally associated with the hispid cotton rat (S. hispidus).
Hantavirus Studies Before 1993
In 1982, Lee et al. (1982) detected antibody reactive with Hantaan virus in 48 of 161 meadow voles (Microtus pennsylvanicus) from Maryland. The virus they characterized (Prospect Hill virus) has not been associated with human disease. Seoul virus, which causes mild hemorrhagic fever with renal syndrome in Asia, was isolated from Norway rats (Rattus norvegicus) from Pennsylvania, Maryland, and Louisiana, and antibodies to SEOV were found in Norway rats in Texas and California (LeDuc et al. 1986). Although the distribution of SEOV in the United States is poorly studied, it has generally been found wherever it has been sought, suggesting that SEOV virus is associated with the Norway rat throughout its range, which includes most urban areas of the United States (LeDuc et al. 1986). Antibody to SEOV was detected in humans in the United States in the late 1980s (Childs et al. 1988a), but was only recently associated with human disease in North America (Baltimore, MD) (Glass et al. 1994).
Studies of the natural history of Prospect Hill virus and SEOV and the rodents that serve as their natural hosts in Baltimore, Maryland, were some of the earliest ecological studies of hantaviruses in the Americas (Childs et al. 1987a, 1987b, 1988b, Korch et al. 1989). These studies illuminated basic concepts of hantavirus natural history that still guide research. The highly specific relationship between hantaviruses and the rodent species that serve as reservoirs of these viruses in nature was apparent from these studies. Even in areas where Norway rats and meadow voles were syntopic, there was no evidence for cross-infection (Korch et al. 1989). The frequent detection of SEOV by reverse transcriptase (RT)–polymerase chain reaction or virus isolation from tissues of antibody-positive rodents was considered evidence that rodents of that species constitute the natural reservoir (Childs et al. 1987a, Arthur et al. 1992), while the sporadic occurrence of antibodies alone in rodents of syntopic species was interpreted as spillover from rodents of the primary reservoir species.
Data from laboratory and field studies led to the identification of mechanisms of SEOV transmission between Norway rats. Experimentally infected rats were much more susceptible to SEOV infection by intramuscular than by aerosol inoculation (Nuzum et al. 1988). Field studies demonstrated an association between acquisition of scars and acquisition of antibody, implicating aggressive interactions (e.g., biting) as a mode of intraspecific transmission of SEOV (Glass et al. 1988). Finally, seroconversion in the field occurred nearly uniformly throughout the year, rather than seasonally, as would be expected if arthropod transmission was involved (Childs et al. 1987b).
Ecological Studies, 1993 to Present
Although antibodies to hantaviruses were detected as early as the mid-1980s in rodents of several species of native North American (Cricetidae) rodents (Tsai et al. 1985), the implications of this finding were not understood until the association of SNV with HPS was discovered in 1993. In the wake of the 1993 HPS outbreak, field studies were designed to achieve a series of goals concerning hantavirus/host ecology and the risk of hantavirus infection in humans (Mills and Childs 1998). These goals initially included identification of the virus–host associations and extended to surveys (cross-sectional studies) designed to identify the potential HPS endemic areas and high-risk habitats, and to determine mechanisms of viral transmission within rodent host populations. Longitudinal studies were initiated to understand the temporal dynamics of hantavirus infection in rodent host populations and to identify environmental factors associated with these temporal dynamics. Currently, data from the cross-sectional surveys and longitudinal studies are being used to build predictive models with the goal of identifying environmental conditions associated with increased transmission of hantaviruses in host populations before these conditions lead to increased disease risk in human populations.
Cross-Sectional (Short-Term) Studies
The goals of the earliest studies were to extend our knowledge of the geographical distribution and natural host range of SNV and other hantaviruses. Host and virus identification in North America often derived from human case follow-up studies (Childs et al. 1994, Hjelle et al. 1995b, Rollin et al. 1995, Rawlings et al. 1996, Ksiazek et al. 1997). Several hantaviruses not associated with human disease were discovered as a result of investigations of HPS cases associated with other hantaviruses (Rawlings et al. 1996), during the course of long-term ecological studies of rodent populations (Sanchez et al. 2001), or by examination of rodent tissues archived in the freezers of museums in the United States (Song et al. 1995, Arai et al. 2007, 2008). Because most North American small mammals are relatively well studied, published distribution maps were available. These maps identify the maximum potential disease endemic area for human disease under the assumption that the distribution of each virus coincides with the distribution of its rodent host. For example, cross-sectional studies showed that, although prevalence of infection varied in space, the distribution of SNV coincided with the known range of the deer mouse in North America, thus effectively outlining the potential geographical range of HPS caused by SNV (Mills et al. 1998, Drebot et al. 2000). Similarly, Bayou virus was shown to occur across much of the range of its principal host, the rice rat (Oryzomys palustris; Ksiazek et al. 1997). In contrast, although the hispid cotton rat (Sigmodon hispidus) is distributed throughout the southeastern United States, Black Creek Canal virus (BCCV) has been found in a limited area of southern Florida. In addition, results of a preliminary study suggest that Muleshoe virus is associated with hispid cotton rats in west Texas (Rawlings et al. 1996), suggesting that two viruses are hosted by one rodent species. An explanation for this apparent lack of a specific one-to-one relationship between host and virus may be that S. hispidus is a complex of species or subspecies (Musser and Carleton 2005). BCCV is restricted to the range of what has been considered S. h. (Sigmodon hispidus) spadicipygus on the southern tip of Florida, while Muleshoe virus may be associated with one or more forms in the western part of the range.
These distributional studies were augmented by regional studies demonstrating differences in host and virus activity and thus differences in relative risk of infection to humans among habitat types. For example, there were distinct differences in deer mouse population density and prevalence of infection with SNV among habitats and elevations in the southwestern United States (Jay et al. 1997, Mills et al. 1997). The lowest deer mouse population densities and the lowest prevalence of SNV infection (and consequently the lowest densities of SNV-infected deer mice) were in the altitudinal and climatic extremes of desert and alpine tundra, while the highest densities and prevalences were at middle altitude habitats including Great Basin scrub and pinyon-juniper woodland. These are also the biomes that are most densely populated in that area by humans (Mills et al. 1997). Habitat distributions of other North American hantaviruses are largely unstudied.
Cross-sectional studies of the ecology of SNV, BCCV, and El Moro Canyon virus helped delineate mechanisms of rodent-to-rodent virus transmission. The association of infection with rodent age suggested horizontal transmission, as had been observed for SEOV and the Norway rat. More frequent infection in males than in females (male bias in antibody prevalence) and the association of wounding with infection (especially in males) implicated aggressive encounters among males as a likely mechanism of transmission (Mills et al. 1997, Douglass et al. 2001, McIntyre et al. 2005, Calisher et al. 2007). The existence of differences in the degree of male bias in antibody prevalence among hosts of different species suggests different mechanisms of transmission or differences in the frequency of various mechanisms of transmission among species (Mills et al. 1999a, Calisher et al. 2007); other possible mechanisms, including venereal transmission (Hinson et al. 2004), vertical transmission, mutual grooming, communal nesting (Calisher et al. 1999, Pearce-Duvet et al. 2006), and environmental contamination have not been carefully investigated.
Longitudinal Studies
Long-term studies are necessary to understand the seasonal and year-to-year patterns in hantavirus–host dynamics as well as to identify the environmental variables associated with those changing dynamics. In 1994, longitudinal mark–release–recapture studies were initiated in four states with a relatively high incidence of HPS: Arizona, Colorado, Montana, and New Mexico (Fig. 3) (Mills et al. 1999b, Douglass et al. 2001). Over 13 years, these studies accumulated more than 1 million trap nights and 44,000 rodents captured. These and other long-term ecological studies (Boone et al. 2002, McIntyre et al. 2005, Safronetz et al. 2008) show that many environmental factors interact to influence host population dynamics and hantavirus transmission in North American rodents (See Fig. 4 for summary schematic).
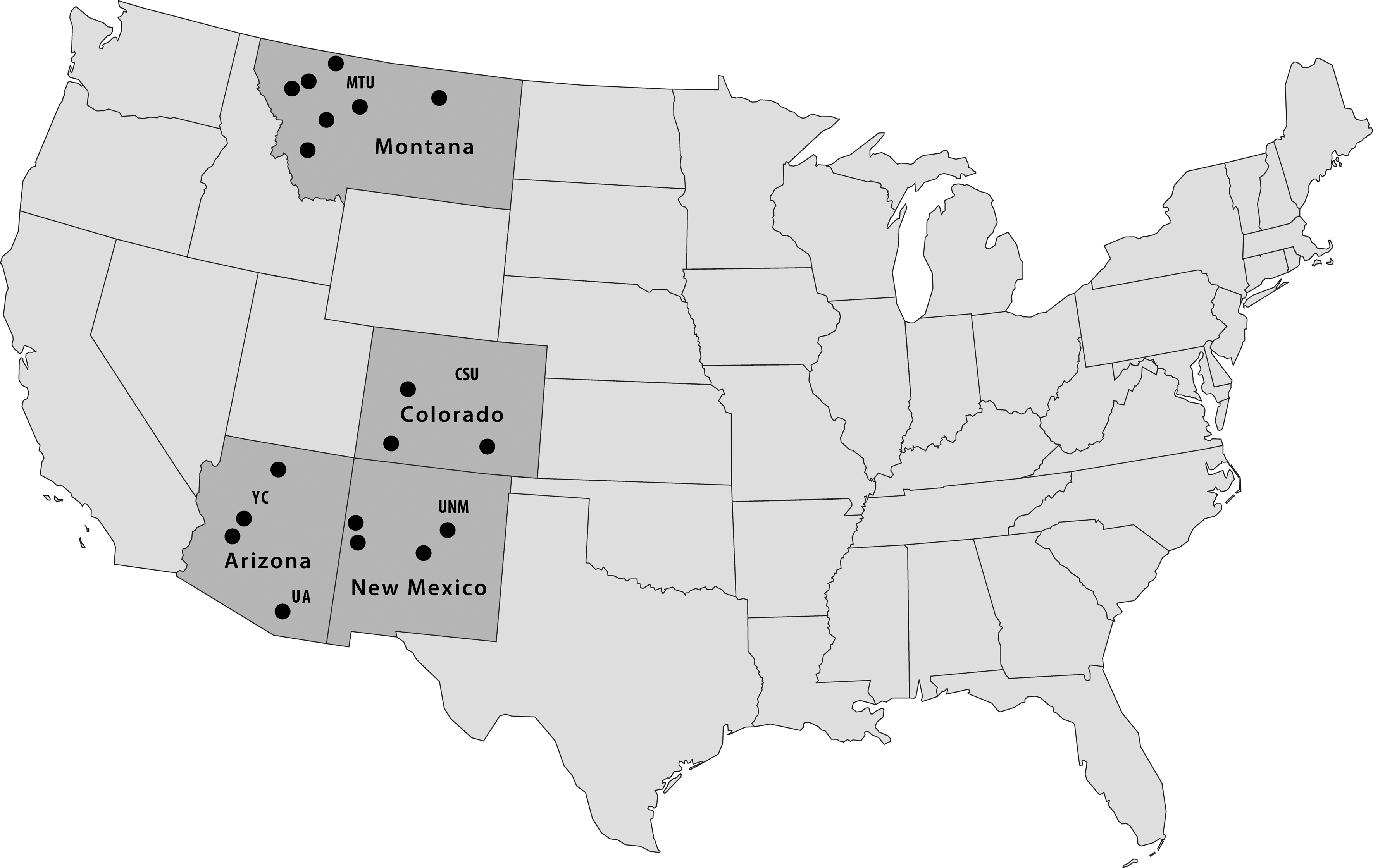
Locations of long-term mark-recapture studies conducted in the United States, 1994–2006. MTU, Montana Tech University; CSU, Colorado State University; YC, Yavapai College; UA, University of Arizona; UNM, University of New Mexico.
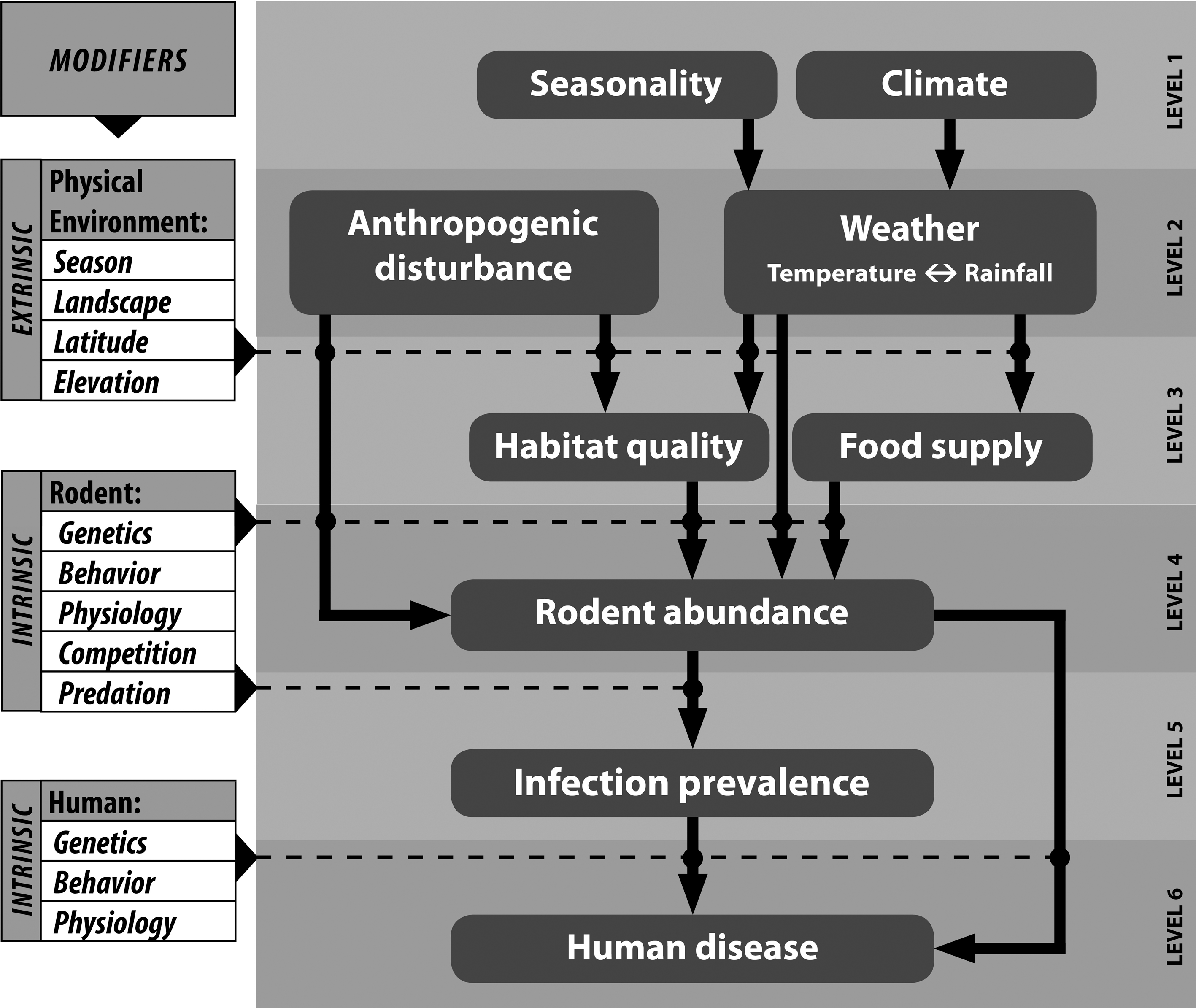
Components of a bottom-up trophic cascade influencing the risk of HPS to human populations. Seasonality and climate affect local weather which, in turn, affects habitat quality and food quality and availability as experienced by rodent host populations, resulting in increases or decreases in reproductive success, growth rates, and survivorship. Rodent abundance is also affected by anthropogenic disturbance, both directly and through its effects on habitat quality. Because hantaviruses are horizontally transmitted in a density-dependent manner, infection prevalence is controlled by rodent abundance. The risk of human infection is a function of the number of infectious rodents in the environment, which is a function of both population abundance and prevalence of infection in the host population. Monitoring of components at each level (right side of graph) can have predictive value for human infection. Monitoring of lower level components provides the greatest lead time, while monitoring of higher level components provides the most accurate risk prediction. “Modifiers” on the left are variables that are extrinsic or intrinsic to host and human populations that are rarely monitored. These variables may have important influences that occur at the intersections of levels. For example, the end effect of weather on habitat quality will depend upon the landscape (e.g., soil type and cover, slope, topography, and vegetation); the effects of rodent abundance on infection prevalence are influenced by the host's genetic susceptibility, level of stress, reproductive condition, risk behavior, and interactions with other species. This is a simplified schematic and potential but poorly understood connections (e.g., the effects of level 2 and 3 components on infection prevalence) and feedback loops (e.g., negative effects of rodent abundance on food supply and habitat quality and potential negative effects of infection prevalence on rodent abundance) are omitted.
For example, increases in reproductive success brought about by environmental regulators such as precipitation, habitat quality, and food availability influence hantavirus transmission rates within rodent populations (Mills 2005). The El Niño Southern Oscillation (ENSO) event that brought increased rainfall to the southwestern United States in 1992–1993 was hypothesized to have resulted in increased population densities and increased transmission of SNV in deer mouse populations via a bottom-up trophic cascade, leading to the 1993 outbreak of HPS (Engelthaler et al. 1999). Because no one was monitoring deer mouse populations in the southwestern United States at the time, this hypothesis remained untested until another ENSO event occurred in 1997. Investigators observed increased rainfall followed by increased vegetative cover and finally dramatically increased abundance of deer mice at trapping sites. These events were followed by 3 years of increased numbers of HPS cases in the southwestern United States (Yates et al. 2002, Mills 2005). Further studies demonstrated that the effects of rainfall can vary, depending on its interaction with other variables, including location, altitude, vegetation type, temperature, and season. For example, at a grassland site in southeastern Colorado, increased rainfall had positive effects on deer mouse populations when temperatures were warm, but negative effects when temperatures were low (Calisher et al. 2005a). Thus, although the component “weather” is represented by a single box in Figure 4, the ultimate effect of weather as a cascade component depends upon the interaction of multiple variables that may act synergistically.
Habitat alteration or disturbance resulting in biodiversity loss can lead to local extinction of populations of rodent species that are habitat and dietary specialists and to population increases in rodents of opportunistic species, many of which are hantavirus reservoirs (Mills 2006). Hantavirus transmission in these high-density host populations is likely to increase, leading to greater risk of infection to humans. These processes in host populations may contribute to the observed negative relationship between host diversity and prevalence of infection with hantaviruses (Mills 2005, 2006). These relationships may only be apparent when long-term averages of rodent population densities and prevalences are taken into consideration.
Host populations in disturbed habitats demonstrate altered population dynamics that may lead to increased risk of infection for humans. For example, peridomestic deer mouse populations had longer breeding seasons and higher SNV infection prevalence than sylvan populations on a Montana ranch. These differences might be related to the more favorable microenvironment in and around buildings and potentially longer survival of virus deposited inside buildings (Kuenzi et al. 2001). This favorable (for mouse and virus) indoor environment might also help explain why HPS cases in the United States are frequently associated with inhabiting or cleaning closed structures with active infestations of deer mice (Armstrong et al. 1995). Although deer mice are frequent inhabitants of houses and outbuildings in rural areas, other hantavirus hosts in North America (e.g., hispid cotton rats and rice rats) only rarely are found in such environments. This contrasting behavior might help explain the relative rarity of HPS cases associated with hispid cotton rats and rice rats. This same logic suggests caution in concluding that hantaviruses associated with some North American hantavirus host taxa that have not been associated with HPS (e.g., Reithrodontomys spp. [harvest mice], Microtus spp. [voles], and even the Soricomorpha [shrews and moles]) are not pathogenic for humans.
Another important concept elucidated by long-term ecological investigation is the role of population density in transmission and maintenance of hantaviruses. The prevalence of infection by hantaviruses in rodent host populations frequently does not show synchronous density dependence. In fact, the highest prevalence of infection may occur during the season when the population density is lowest (Boone et al. 1998, Douglass et al. 2001). An explanation for the lack of synchronous density dependence may be that the highest host population densities occur at the end of the breeding season, when the population consists of a high proportion of young (and therefore uninfected) individuals. It has been proposed that the highest prevalence of infection might occur in the spring of the year, before the young of the year have entered the population and the population age structure is oldest. Finally, it is hypothesized that the prevalence of infection in this population is proportional to the peak population density attained during the previous breeding season. This delayed-density-dependent prevalence of infection, which was proposed to explain changes in the prevalence of Puumala virus infection in bank voles (Niklasson et al. 1995), was hypothesized as a universal phenomenon for horizontally transmitted rodent-borne pathogens in temperate habitats (Mills et al. 1999a). Delayed density dependence was recently demonstrated in Montana deer mice. SNV antibody prevalence peaked in spring and was proportional to the peak density of deer mouse populations the previous autumn (Madhav et al. 2007). Although spring prevalence in females was much lower than in males when autumn densities were low, prevalence in females increased more rapidly at higher densities, approaching the prevalence in males at the highest densities. This suggests that female deer mice may frequently interact with other deer mice only at relatively high population densities (Madhav et al. 2007).
In contrast to SEOV in Norway rats (Childs et al. 1987b, 1988a), native North American hantavirus rodent host population densities and infection prevalences may vary greatly among years. The virus may even become extinct in some local populations (especially sites with low population densities in marginal habitats), only to be reintroduced from nearby source populations a few months or years later (Abbott et al. 1999, Calisher et al. 2005b, Kuenzi et al. 2007). This suggests that a metapopulation structure or source-sink dynamic may influence the overall pattern of hantavirus dynamics and persistence (Glass et al. 1998, Boone et al. 2000) and that habitat heterogeneity (both spatial and temporal) might serve as indicators of changing patterns of risk.
Predictive Modeling
Because there is no vaccine and no specific treatment for HPS, the best current defense measure is prevention of infection. Effective intervention relies on the identification of times and places of increased risk (Mills and Childs 1998). An important goal of longitudinal studies of hantavirus host populations has been to identify environmental factors associated with increases in host population density to develop predictive models of HPS risk for humans.
Data from longitudinal studies of SNV and deer mice demonstrated an association among increases in rodent population density at specific sites, increased virus transmission in host populations, and increases in human cases of HPS (Yates et al. 2002). Knowledge of this association allowed prediction of increased risk of human disease and the publication of advanced warning (CDC 1998, 1999), possibly resulting in reduced morbidity and loss of life. These predictions represent the use of simple models based on direct observation of rodent population trends. Although useful, such models, based on observations at levels 4 and 5 (Fig. 4), provide short lead-time and are possible only for local areas where rodent populations are being monitored. To develop models that provide greater lead-time and cover wider geographic areas, researchers have focused on environmental conditions at lower levels of the bottom-up trophic cascade hypothesized to lead to increases in rodent population density and on the use of broad-coverage satellite data capable of monitoring components at levels 2 and 3 (Fig. 4).
Because of their universal availability and long lead-time, weather data (especially rainfall) were among the first potential predictors to be explored (Yaffee et al. 2008). Although state-of-the-art forecasting tools were employed, addition of weather variables to simple density-dependent models based on counts of deer mice did not improve predictability. Part of the explanation for this result may be that the same amount of rainfall can have highly divergent effects on vegetation depending upon numerous factors, including location, slope, topography, soil type, vegetation type, and time of year. More successful models have circumvented the variable effects of rainfall by using vegetation or indicators of vegetation quality as a predictor variable.
Environmental conditions monitored by satellites identified sites likely to support increased abundance of infected deer mice in a given season (Glass et al. 2000, 2002, 2006). Some locations showed persistently favorable environmental conditions over several years (Glass et al. 2007). These sites are hypothesized to serve as “refugia”—areas where deer mice and SNV thrive—while unfavorable conditions elsewhere result in low deer mouse population densities and low prevalence (or local extinction) of SNV. These refugia would represent sources for deer mouse repopulation and reintroduction of SNV when local conditions improve (Yates et al. 2002).
Loehman et al. (manuscript in preparation) used gross primary productivity (GPP) derived from the moderate resolution imaging spectroradiometer to predict deer mouse abundance at Montana trapping sites for 5 years. Rodent abundance tracked GPP closely for 4 years, but rodent populations decreased the 5th year while GPP continued to rise. Factors not included in the model (e.g., predation, competition, or intrinsic population factors that may operate at high population densities) may have been more important in the 5th year than the factors included in the model.
Clearly, extrinsic and intrinsic factors affecting rodent population dynamics and hantavirus transmission are myriad (Fig. 4). These factors do not act independently and their interactions will add further complexity to predictive models. We believe that useful models are possible, but their development will require the considerable multidisciplinary efforts of mathematicians and ecologists and they must be based on truly long-term monitoring of rodent populations and environmental variables. These monitoring programs will be measured in decades rather than a few years.
Laboratory Studies of Hantavirus Infection in the Principal Host
Laboratory studies using pathogenic American hantaviruses in the natural host must be conducted in biosafety level (BSL)-4 laboratories or in natural outdoor enclosures by investigators wearing appropriate personal protective equipment (Botten et al. 2000). As a further complication, SNV is difficult to grow in cell culture and deer mouse–to–deer mouse SNV virus transmission in the laboratory setting has been challenging to demonstrate (Botten et al. 2002).
In the earliest laboratory studies of North American hantaviruses, Hutchinson et al. (1998, 2000) used BCCV and hispid cotton rats to confirm patterns observed in Old World hantavirus/host systems, including transient viremia, long-term presence of infectious virus in organ tissues, long-term shedding of virus in urine and saliva, and horizontal transmission of virus between cage-mates. One finding, however, contrasts with other field and laboratory studies. For other hantaviruses, pups of infected dams acquire protective maternal antibody, either in utero or by breastfeeding, and vertical transmission appears not to occur (Zhang et al. 1988, Dohmae and Nishimune 1995, Mills et al. 1997, Borucki et al. 2000, Botten et al. 2002, Kallio et al. 2006). BCCV, however, appears to have been transmitted from infected dams to offspring. Cotton rats used in these studies were a subspecies from the midwestern United States (Hutchinson et al. 2000), where BCCV does not occur. In light of more recent findings concerning associations between hantaviruses and cotton rats at the subspecies level, additional experiments are needed.
For SNV in the deer mouse, investigators (Botten et al. 2002) demonstrated a very different pattern from those described above. In contrast to studies with Hantaan, Seoul, Puumala, and BCCVs, SNV shedding was not detected in urine or feces, although viral RNA was detected intermittently in saliva. Intracage transmission, which was easily demonstrated for other hantaviruses, was detected in only 1 of 46 mice exposed. The apparent poor efficiency with which SNV was shed by its host may be an important factor in the relatively low incidence of human infection (Safronetz et al. 2008).
Above we suggested that male bias in the prevalence of hantavirus infection in hosts of many species was due to gender-specific differences in behaviors that serve as risk factors for transmission, such as aggressive interactions. We also suggested that the correlation between scars and infection is because aggression leads to infection. Recent laboratory studies suggest that the cause-and-effect relationship between aggression and infection may be complex. Male Norway rats infected with SEOV were more aggressive than were uninfected males, and aggressive males had more virus in their tissues than did less aggressive males (Klein et al. 2004b). Differences in host immune response to infection also may impact the likelihood that infected individuals remain infectious. After inoculation with SEOV, male rats had more viral RNA in target organs and shed viral RNA longer than did females. This was associated with sex-related variation in the expression of genes involved with immune function (Klein et al. 2004a). The expression of key transcriptional factors and genes that encode for proinflammatory, antiviral, T cell, and Ig superfamily proteins was higher in females than in males. Conversely, males had higher expression of heat shock protein genes, suggesting elevated cellular stress. These patterns indicate that infection in males may be longer and more productive than similar infections in females. Lehmer et al. (2007) reported experimental evidence that male deer mice were less immunocompetent than females and suggested that this condition helps explain the commonly observed higher infection prevalence in males than females.
Studies suggest that regulatory T cells contribute to the persistence of hantaviruses in rodents (Easterbrook et al. 2007, Schountz et al. 2007). These cells, which suppress proinflammatory responses, were more numerous during the persistent phase of SEOV infection in male Norway rats. Inactivation of regulatory T cells caused a reduction in SEOV RNA in target organs and to decreased shedding of viral RNA in saliva (Easterbrook et al. 2007). Results consistent with regulatory T cell effects have been described in a similar system for deer mice (Schountz et al. 2007).
Synthesis and Future Directions
Over the last 14 years, the increase in our understanding of North American hantavirus–host ecology has been dramatic. Nevertheless, many challenges remain. As studies continue, relatively simple patterns that first emerged have been shown to be inaccurate or incomplete or time- or site-specific. For example, the initial generalization that ENSO events and increased rainfall lead to increases in host population density and increased risk of human infection may be true only for the arid Southwest. The effect of rainfall on host populations cannot be considered in isolation; its interactions with many other environmental variables must be considered to achieve a predictive understanding (Mills 2005). Other patterns deduced after a few years of study, such as the linear relationship between GPP and deer mouse population density in Montana, were shown to be incorrect after longer-term investigations (Loehman et al., manuscript in preparation). Indeed, because of the stochastic nature of environmental conditions and host responses to environmental conditions, some associations (such as the negative relationship between rodent community diversity and prevalence of hantavirus infection) were only discerned by using long-term averages (Mills 2005).
Observations from laboratory studies do not necessarily hold when applied to field conditions. For example, transmission of SNV among deer mice was rare in the laboratory (Botten et al. 2002), but appears to occur readily in the field. Transmission in nature may require specific factors (e.g., environmental stressors affecting immune function, territorial boundaries, and population dominance hierarchies) that were not replicated in the laboratory. Although transmission of BCCV to offspring was observed in the laboratory (Hutchinson et al. 2000), there is no evidence that it occurs in the field (Glass et al. 1998). Nevertheless, laboratory studies can be important for helping to interpret field observations (e.g., aggression among hosts leads to infection, but the converse may also be true [Klein et al. 2004b]). The widely held assumption (principally derived from results of laboratory studies) that hantaviruses do not have important effects on host fitness has been challenged by a field study that suggests both a population effect and an individual physiological response of infection: survival of infected (antibody-positive) young deer mice was lower than that of uninfected mice, and recently infected (seroconverting) deer mice gained less weight than mice that did not seroconvert (Douglass et al. 2001, 2007). A laboratory study demonstrating that chronically infected deer mice had weaker immune responses than uninfected mice provided a possible mechanism for this reduced health (Lehmer et al. 2007). Laboratory and field studies are complementary, and accurate conclusions often require both. Finally, field studies in natural environments should be supplemented with studies in disturbed peridomestic environments where most cases of HPS are acquired (Kuenzi et al. 2001).
The recent finding of several species of insectivores that appear to host hantaviruses in many parts of the world, including North America (Arai et al. 2007, 2008), challenges our long-held assumptions about the origin and cospeciation of hantaviruses and muroid rodent hosts and opens a new field of investigation: Is there a clade of hantaviruses that has cospeciated and radiated along with the insectivore lineage? Did the insectivore hantavirus lineage derive from the muroid lineage or vice versa? Are insectivore-associated hantaviruses pathogenic for humans? Although the similarity of phylogenetic trees provides strong evidence for codivergence of hantaviruses and hosts, this is not the complete story. Virus spillover in rodents is common, even across family lines (e.g., Childs et al. 1994), and host jumping between subspecies, species, and genera has apparently occurred (Vapalahti et al. 1999, Plyusnin and Morzunov 2001). In North America, cross-species host jumping events have been proposed for New York virus (Morzunov et al. 1998) and for Limestone Canyon virus (Sanchez et al. 2001). Cross-taxon jumps at higher taxonomic levels should be less common, but may have occurred, especially among syntopic species. It is highly unlikely that hantaviruses associated with muroid rodents and with insectivores diverged from a hantavirus associated with a common ancestor to both groups. Nevertheless, a much more recent host jumping event between these two lineages with similar ecological niches is possible. Extensive sampling and detailed phylogenetic analyses will be needed to reveal the true explanations for the patterns observed.
The practical end to ecological studies of hantaviruses is not the understanding of hantavirus–host ecology in itself, but the application of this understanding to the development of risk-reduction interventions aimed at decreasing the burden of human disease. Pathogen and host identification, delineation of endemic areas and high-risk habitats, and specification of seasonal risk patterns have obvious practical value and have been achieved for several areas in the United States. Predictive models are beginning to show increasing success (CDC 1998, 1999, Glass et al. 2006, Loehman et al. [manuscript in preparation], Luis et al. [manuscript in review]), but their continued improvement depends upon the establishment, maintenance, and continuity of long-term monitoring programs.
The need for future studies is great. Ecological studies are still lacking in the eastern United States, where SNV, New York virus, Monongahela virus, BCCV, or Bayou virus are common, and we do not know what host or virus factors are responsible for the lower incidence of human disease in the eastern United States. Most human cases of HPS are acquired from exposure to virus in peridomestic environments, and host population ecology in these habitats is different from that in more natural, undisturbed settings, yet ecological studies in these environments are still rare. The utility of field studies has been limited by the inability to control population and environmental variables, while laboratory studies suffer because of their inability to replicate natural conditions. Future field studies might employ outdoor enclosures that would provide seminatural conditions while allowing the control of some population and environmental variables such as population density, age structure, sex ratios, the availability and quality of food and water, and the presence of competitors and predators (Bagamian et al., unpublished data).
Several HPS cases in the United States have occurred very close to the border with Mexico (CDC, Special Pathogens Branch, unpublished data) and rodent species that are known to host hantaviruses that cause HPS in the United States (P. maniculatus, Peromyscus leucopus, and S. hispidus) occur in large areas of Mexico; yet, no HPS cases have been reported in Mexico. Mexico is home to a much higher diversity of sigmodontine and neotomine rodents than the rest of North America (Musser and Carleton 2005), and preliminary serosurveys have reported hantavirus antibodies in rodents of at least eight species (Mantooth et al. 2001, Suzan et al. 2001, Chu et al. 2008, Castro-Arellano et al. 2009). Nevertheless, except for Playa de Oro virus (Chu et al. 2008), the viruses associated with these hosts remain largely uncharacterized. The diversity, distribution, and natural history of hantaviruses in Mexico and their importance for human disease are almost completely unstudied. Our understanding of the diversity and ecology of North American hantaviruses will remain incomplete until intensive studies in Mexico are undertaken.
Although laboratory studies of SNV are difficult for reasons already mentioned, they are sorely needed. Effects of infection on hosts, the relative efficiency of potential mechanisms of transmission, host–virus specificity and the potential of secondary hosts, pathogenesis, viral shedding, viral recrudescence, and many other problems are best investigated in the controlled environment of the laboratory.
Improvements in the nascent field of predictive modeling for hantaviruses are also needed and will require the continuation, perhaps for decades, of long-term monitoring programs for host populations and environmental variables. Unfortunately, traditional funding agencies do not generally support long-term studies. We hope that the increasing recognition by ecologists and public health officials of the value of long-term monitoring programs (e.g., Institute of Medicine Workshop on Global Climate Change and Extreme Weather Events: Understanding the Potential Contributions to the Emergence, Reemergence and Spread of Infectious Disease [Institute of Medicine, 2008]) will lead to the development of needed long-term funding sources.
Footnotes
Acknowledgments
We thank Craig Manning (CDC) for assistance with art and graphics. Pierre Rollin and Barbara Ellis (CDC) and Charles Fulhorst (University of Texas Medical Branch) provided helpful comments that improved the article.
Disclaimer
The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the Centers for Disease Control and Prevention or the Department of Health and Human Services.
Disclosure Statement
No competing financial interests exist.