
Abstract
Chikungunya virus (CHIKV) has long been considered to be transmitted to humans by the human-biting mosquito Aedes aegypti, especially in Africa. However, the recent outbreak of CHIKV involved another vector, Aedes albopictus, and serological data in the literature suggest that several species of domestic or human-related vertebrates can be contaminated by this virus. However, the role of Ae. albopictus mosquitoes as potential enzootic vectors for CHIKV has not yet been evaluated. Here we investigate Ae. albopictus feeding and resting behaviors in an area where a CHIKV epidemic recently occurred, which means deciphering host-seeking and feeding behaviors on several vertebrate species, measuring endophagous/exophagous (activity), endophilic/exophilic (resting) behaviors and its diel (24 h, day/night) biting activity. Ae. albopictus was found to have bimodal daily feeding activities and was found to have exophagic (89%) and exophilic (87%) behaviors. Ae. albopictus showed an opportunistic feeding behavior on a wide range of hosts (from cold-blooded to warm-blooded animals), supporting that it can be implicated in various vertebrate–virus pathosystems. However, with equal availability of one of the four vertebrate hosts (calf, chicken, dog, and goat) proposed against human, Ae. albopictus significantly preferred human, supporting earlier data about its high degree of anthropophily. Multiple blood feeding was also reported in every combination (animal/human) offered to Ae. albopictus, enlightening the higher risks to spread an arbovirus to human population because of interrupted feeding. Such catholic behavior suggests that Ae. albopictus may act as a bridge vector for zoonotic viruses. Further epidemiological implications of this issue are discussed.
Introduction
This epidemic wave was characterized not only by its extensive geographic range, but also by its new vector, Aedes albopictus, instead of Ae. aegypti. In Kenya and the Comoros archipelago, Ae. aegypti was the only vector implicated; in Madagascar both Ae. aegypti and Ae. albopictus are present, and in the other islands Ae. albopictus was the sole vector responsible of the transmission of CHIKV (see map in Delatte et al. 2008b). Ae. albopictus originated in Asia (Skuse 1894) and has spread in those last 20 years to many countries in Europe and America, and some countries in the African continent (Benedict et al. 2007), where it replaced Ae. aegypti in some ecological niches (Juliano and Lounibos 2005). One of the consequences of the interaction between this new vector, Ae. albopictus, and CHIKV was the selection of a new CHIKV strain. Indeed, this viral mutation on a membrane protein (E1 226V, instead of 226A) allowed the virus to be more adapted to its vector and increased its replication rate in Ae. albopictus (Schuffenecker et al. 2006, Tsetsarkin et al. 2007, Vazeille et al. 2007).
Although able to transmit a large number of arboviruses (Turrell 1988, Mitchell 1995), the species has generally been considered as a secondary vector because of its zoophilic behavior. Nevertheless, it might be worth noting that Ae. albopictus has been a primary dengue vector in circumstances where Ae. aegypti is rare or absent, like La Réunion (Paupy et al. 2001), or such as Hawaii (Effler et al. 2005) and Macao (Almeida et al. 2005). Since 2005–2007 where its involvement in the CHIKV epidemics as primary vector has been demonstrated (De Lamballerie et al. 2008), Ae. albopictus has became a worldwide concern. Thus, the epidemiological role of Ae. albopictus has clearly been shown for CHIKV transmission from human to human, but its precise role in potential enzootic transmission remains unclear.
The importance of enzootic transmission depends on the trophic behavior of the mosquito for the different vertebrate species, the availability of these species, and ultimately the replication capability of the virus in the vertebrates. CHIKV is well known to replicate in primates (Diallo et al. 1999). First, CHIKV antibodies have been found, with a low frequency, in Africa or Asia in different animal species such as zebu (Guilherme et al. 1996), rodents, or birds (Jupp and McIntosh 1988, Inoue et al. 2003). Further, Ae albopictus is known to feed on several animal species in Asia, where it originates (Smith 1956, Hawley 1988), as well as in the other colonized continents (Hawley 1988). No study has ever been conducted in the islands of Southwest Indian Ocean.
Human-biting rate is a key variable in the basic reproduction (R0) rate of arboviruses. R0 is one of the most important concepts in infectious disease epidemiology, and most often defined as the average number of secondary cases caused by one infectious individual placed in a population of susceptible individuals or more generally as the expected number of secondary cases per primary case in a naïve population (Diekman and Heesterbeek 2000). For a mosquito-borne disease, R0 is generally defined as
To evaluate the importance of zoophilic behavior, biting behaviors are de facto a very important aspect to study, which implies assessing the host choice, the time, and the place of the blood meal. Up to now, no studies on feeding behaviors of Ae. albopictus have ever been done on the populations of the Indian Ocean, and only a few data are available for Ae. albopictus in general. This lack of knowledge might be due to the difficulty to find resting blood-fed Ae. albopictus in nature, which would allow to identify the origin of the blood and thus assess the feeding preference in nature. To better understand host-feeding behavior of Ae. albopictus, we conducted different experiments using populations from La Réunion and targeting its host preference in nonchoice and choice experiments, its diel (i.e., the 24-h physiological time unit made up of 1 day and 1 night) biting activity, and its resting-place behaviors.
Materials and Methods
La Réunion is a tropical French island (2500 km2) situated in the Indian Ocean (21°16′ S, 55°31′ E), east of Madagascar with about 770,000 inhabitants. There are two distinct seasons: a cool and dry winter from May to October, and a warmer, rainy summer from November to April. The habitat is usually composed of houses with gardens, with very few high buildings. Different experiments were carried out in the outskirt of Saint Pierre, in different seasons according to the protocols (see below).
Host preference experiments
Mosquito source
Ae. albopictus used in the two host-feeding experiments were obtained from field-collected eggs in artificial containers from Saint Pierre (La Réunion Island) and kept in insectarium (F0 were used). The insectarium was maintained under approximately 27.5 ± 1°C, 80 ± 20% relative air humidity, and natural photoperiod. For both experiments, 5-day-old mated adult females of the F0 generation and starved from the morning were used. The range of natural daylight (dawn–dusk) vary from about 10 to 14 h during the year. Sunset varies from about 17 h 20 min to 19 h 00 min, and sunrise from 5 h 20 min to 7 h 00 min according to the season.
Nonchoice host feeding on 12 animal species
Experimental design
Tested animals were classified by their sizes into two categories (large and small); according to their category, a different protocol was used. Two specimens of each species were tested. For each large animal (human, goat, pork, cow, and dog) a small cage (length, 15 cm; width, 7 cm; height, 5 cm) covered with white polyester tulle was used. Inside each cage, 10 unfed female mosquitoes were placed. Cages were kept for 90 min on the belly of each animal during the highest biting activity of female mosquitoes: 16 h–17 h 30 min (for the month of the study, period chosen according to the diel biting activity; see Results section). Goats and cows were shaved where the cage was placed; the cage was placed under the belly of the dogs, where hairs are scarce. Each small animal (duck [Anas sp.], chicken [Gallus gallus], rat [Rattus norvegicus], chameleon [Chameleo pardalis], gecko [Hemidactylus sp. and Gehyra mutilata], mouse [Mus musculatus], and shrew [Suncus murinus]) was placed individually in a cage (length, 20 cm; width, 20 cm; height, 20 cm) with 10 female mosquitoes. The animals were kept inside each cage for 90 min from 16 h to 17 h 30 min.
All these experiments were repeated three times for each animal of each species (in total six times per species). After the 90 min of meal access, mosquitoes were caught with an aspirator, frozen at −80°C, and subsequently examined to determine their feeding status.
Statistical analysis
A first Kruskal–Wallis rank sum test was applied to our data set to test if the different rates of blood-feeding mosquitoes were different (at a level of 5%). Then, a rank test, the pairwise comparisons using the Wilcoxon rank sum test (at a level of 5%) with a Bonferroni adjustment, was used to compare the different rates of blood-fed mosquitoes on different hosts. All statistical tests were performed with R software (Team 2004).
Choice host-feeding experiment on four animal species compared to humans
Experimental design
Four animal species were chosen according to the results of the nonchoice experiment and their ability to be manipulated for choice experiments: three were species with the highest biting rates recorded (chicken, dog, and calf ) and one was less bitten (goat). Further, these animal species are often found in backyards of the traditional habitat in Réunion island (except calves, which are present in farms and fields). The number of individuals in each species to be compared to one human was chosen to level out skin surface. To evaluate the equivalent skin surface for each species, we used the Haycock's formula (1978) for human, and the uniform formula of Wang and Hihara (2004) for dog, chicken, goat, and calf. Basic measurements of skin surfaces involving only weight and height used in clinical veterinarian schools were also checked, and gave similar results. Thus, we used 11 chickens of 3.3 kg each; two 8-month-old goats of 20 and 18 kg, respectively; two dogs of 18 and 19 kg, respectively; and a 5-day-old calf of about 45 kg. The three different human volunteers (two females and one male, members of the research group and authors of this article) had an equivalent skin surface. Each human was wearing short pants and a short-sleeve shirt, and laid still on a 80 cm × 2 m mattress with a cloth on the face (to prevent mosquito bites on the face). The hair or feather density covering each animal was considered as equivalent to the clothes of each human, and prevented the volunteers to perform the experiment naked. Each of the three humans was tested individually against each of four animal species.
Each human volunteer was introduced into a large mosquito net (length, 2.90 m; width, 2.10 m; height, 1.80 m). Then, the animal bait was introduced at 1.1 m from the human bait in a cage (length, 1 m; width, 1 m; height, 1.5 m) with large grids (4 × 4 cm, allowing mosquito movements in and out). When both bait species were placed, 110 female mosquitoes were released in the middle of the net for 90 min during the peak activity of Ae. albopictus females (16 h–17 h 30 min). The mosquito net was strapped using a plastic cover to prevent mosquitoes from escaping (Fig. 1). After a period of 90 min, the human present in the net captured all the blood-fed mosquitoes within the net with a mouth-aspirator. The totality of mosquitoes captured never reached the released number; this could be because of unseen tiny holes allowing mosquitoes to escape, or mosquitoes squashed by the animals, or the catcher missing some of them. The order of the tested species for each human was randomized for chickens, goats, and dogs, but not for the calves. Indeed, the availability of the borrowed animal forced us to group the testing within 3 days at different hours of the day.
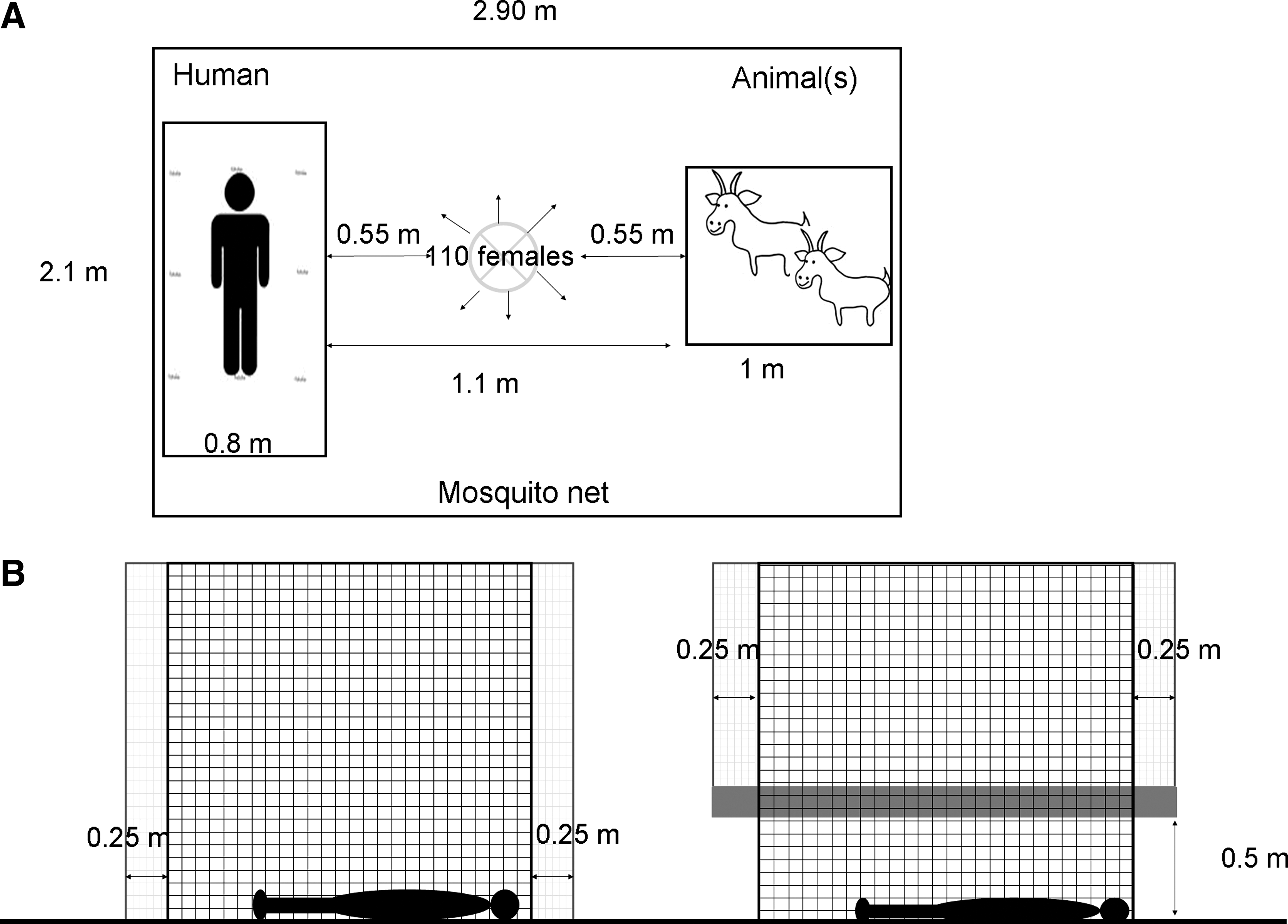
Experimental design for the choice experiment (
After each experiment, all blood-fed mosquitoes were individualized and stored at −80°C. Then, an enzyme-linked immunosorbent assay test was performed on each mosquito for blood meal identification; according to the animal species tested (human, goat, dog, and chicken) different antibodies were used. Each mosquito was ground in phosphate buffered saline; each extract was transferred into two microtiter plates. Then, the specific antibodies against the two animals tested (one in each plate) in the choice experiments were used in an enzyme-linked immunosorbent assay reaction as described in Beier et al. (1988). Positive controls used in each plate were a dilution (1/100) of a sample of blood of each animal species tested. Negative controls were naïve, nonblood-fed Ae. albopictus from our lab rearing, ground in the same conditions as above. Two positive and two negative controls were used. A positive reaction was stated when the optic density recorded was twice the amount of the average of the two negative controls.
All animal studies were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and approved by the Direction des Services Vétérinaires de la Réunion. All human volunteers involved in these experiments had read the protocols and agreed to participate. Female mosquitoes used were from lab rearing and were free of pathogens.
Statistical analysis
For chicken, goats, and dogs, the number of fed Ae. albopictus females per human person (H1, H2, or H3), animal species, and blood origin (human, animal, or both) were analyzed using a Poisson log linear model (PROC GENMOD, link log; SAS, version 9), which is the equivalent to the multinomial response model (McCullagh and Nelder 1998). The explicative variables were the animal species, the human person, and the blood origin. The log of the total number of blood-fed mosquitoes was taken into account as an offset variable. The significance of explicative variables and their interactions were tested by likelihood ratio (p < 0.05). For each significant variable, the exponential of regression coefficient estimates (RC) (also called incidence rate ratio estimates, Dohoo et al. 2003) were used to assess the relative change in the number of fed mosquitoes for one level compared to a reference. These RC are equivalent for counted responses to the odds ratios that we would obtain for binary outcomes (e.g., infected vs. not infected). For example, in the experiment combination of goats and H1, an RC of 1.5 with animal as reference level means that the mosquitoes have 1.5-fold more chance to have fed on the human than on the animal. A similar analysis was run separately for the calf (without the species factor), owing to the fact that the experiments were run in different conditions than with the other species.
Diel biting activity and endophagous/exophagous activity
The experiment was carried out four times (twice during the summer and twice during the winter) in Saint Pierre in a peri-urban area (in 2007).
Experimental design
A two-layer mosquito net was placed in the garden (under a tree to provide shade the whole day) of the chosen house, and a second in the bedroom. The chosen house (four bedrooms, one kitchen, one dining room, and one bathroom) had a large garden and was close to sugarcane fields. The chosen bedroom was facing the back of the garden and was in front of the outside mosquito net. The outside two-layer mosquito net was placed at approximately 15 m from the house (and the window of the chosen bedroom). The two-layer mosquito nets consisted of a first inner mosquito net (length, 2.4 m; width, 1.60 m; height, 1.80 m) sawn to a second layer (length, 2.90 m; width, 2.10 m; height, 1.80 m), which was placed outside. A human was used as bait and left during 24 h in the inner part of the mosquito net (mosquito-proof ) to prevent him to be bitten by mosquitoes. The second mosquito layer was above the first one with 25 cm gap between both mosquito net layers and rolled up at 50 cm above ground to allow the attracted mosquito to come close to the first mosquito layer. Each experiment was started at 8 h am and carried on for 24 h. Every 30 min the outside layer was rolled down to the ground to prevent attracted mosquito to escape. Subsequently, all trapped mosquitoes were caught with a butterfly net. After every capture (every 30 min) the outside layer of the mosquito net was rolled up again (Fig. 1). The same experimental setup was used in parallel in a bedroom of the house. All the captured mosquitoes were placed in 5 mL tubes individualized according to the hour and place of capture (garden or house), and then frozen (−20°C). Mosquitoes were then identified, sex determined, and counted.
To avoid an intrusion effect, the mosquito nets were installed the day before the experimentation.
The endophagic and exophagic rates were calculated. The endophagic rate corresponded to the amount of indoor-captured mosquitoes around the human bait over the total amount of captured mosquitoes. Subsequently, the exophagic rate was the amount of outdoor-captured mosquitoes around the human bait over the total amount of captured mosquitoes.
Resting places of wild Ae. albopictus
Experimental design
In total, 105 houses with gardens were visited within two selected residential areas of La Réunion (one on the East coast and the other on the West coast). To maximize the numbers of collected mosquitoes, within each neighborhood, houses were chosen with some vegetation that provided a shaded environment for resting mosquitoes. Typically, grass lawn with flowers was found in the front, and trees (mango, papaya, and lemon) and aromatic plants were found in the back yard, in addition to hedges of small ornamental shrubs around each garden. Chickens were sometimes reared in the back yard. Mosquitoes were caught between 9 h 00 min and 14 h 30 min, excluding the highest biting activity of females. Outside collections were made by two people, one smoothly beating the vegetation (with a broom stick) and the other right behind with a butterfly net, to catch all the insects coming out. Every parts of each garden were processed as described. Inside the houses, mosquitoes were caught with mouth-aspirators. Immediately after collection, each sample was placed in a 5 mL tube on wet ice in a cooler (gardens' and houses' caughts were separated). Samples were then transported to the laboratory for processing. Subsequently, mosquitoes were sorted under a binocular (counts and sex and species identification were made). In addition, females were examined to determine their gonotrophic status (unfed, blood fed, gravid, or half-gravid).
Results
Host (nonchoice and choice) feeding experiments
Nonchoice experiment
The Kruskal–Wallis test performed on our data set was significant (χ 2 = 38.67, ddl = 11, p < 0.001), which showed a global differentiation between groups of hosts. According to the Wilcoxon pairwise test (p < 0.001) six groups of hosts were found (Fig. 2). A first group of very high rates of blood-fed females on chickens, humans, dogs, and cows with blood-fed mosquito rates above 70% was observed. Then, five other groups were observed with blood-fed mosquito percentages ranging from 2% to 42%. The lowest rates of blood-fed females were observed for the cold-blooded animals tested: geckos and chameleons (Fig. 2).

Percent of blood-fed mosquitoes within each tested vertebrate species that fed on that species in the nonchoice experiment. Columns with the same letter indicate that the results are not significantly different (p > 0.05, Wilcoxon pairwise comparison rank test). Vertical bars represent the standard errors.
Choice experiment
For the experiments with chickens, goats, and dogs, the interaction between human person * animal species * blood origin was significant (p < 0.05), showing that the strength and/or direction of the preference varied according to these factors. Whatever the different animal species/human pairs, mosquitoes significantly preferred human blood to the other tested animals, up to 11.1-fold (Table 1A). Human-fed mosquitoes similarly bit the three persons, except with goats, where H3 was more likely to be bitten than H1 (Table 1B, Fig. 3). Animal-fed mosquitoes were more likely to have fed on goats than on dogs or chickens in experiments with H1 and H2. No differences were seen with H3 (Table 1C). Further, in all the experiments blood feeding on both animals and humans was detected. A significantly higher rate of mixed blood was found with chickens compared to goats in experiments with H1 and H2. Again, no differences were seen with H3 (Table 1D).
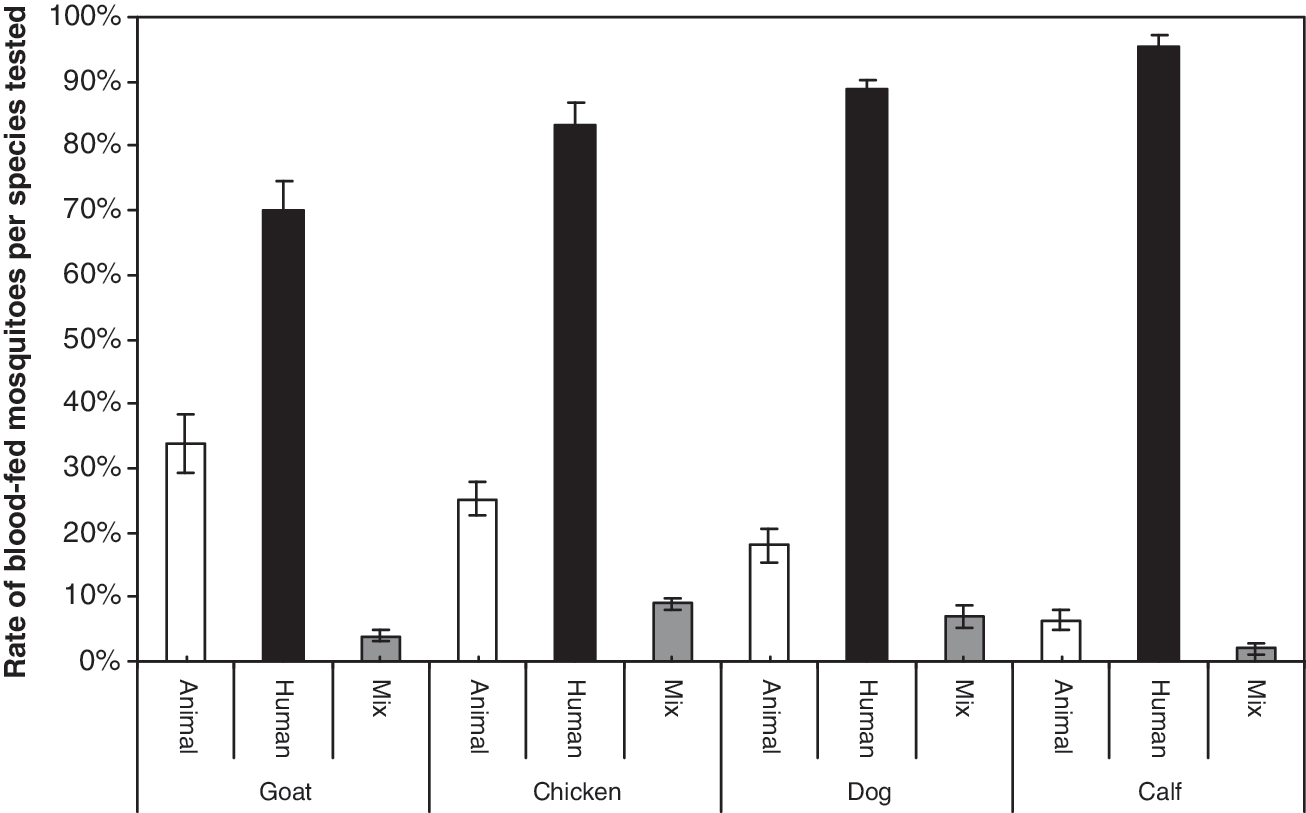
Percent of blood-fed mosquitoes on animal, human, or both (mix) within each tested vertebrate species in the choice experiment performed by enzyme-linked immunosorbent assay tests. Average percentages of blood-fed mosquitoes presented for human and animals include the mix cases. Vertical bars represent the standard errors.
H1, H2, and H3 are the three human volunteers.
The bold type is meant to indicate significant regression fitted to the Poisson model.
Regression coefficient estimate assessing the relative change of the number (A) of mosquitoes in feeding on humans compared to animals, (B) of human fed mosquitoes in feeding on one given person compared to another one, (C) of animal fed mosquitoes in feeding on one species compared to another one, and (D) of mixed fed mosquitoes in feeding on one given species compared to another one. Mixed fed mosquitoes are those that fed on both humans and animals.
p < 0.01.
p < 0.001.
p < 0.05.
For the calf experiment, the only significant variable was blood origin (p < 0.05), showing that mosquitoes highly preferred human to calf (RC = 21.3 [confidence interval: 11.6, 39.0]) and to multiple feeding (RC = 46.8 [confidence interval: 19.3, 113.5]). The human person factor was not significant.
Diel biting activity and endophagous/exophagous activity
In total, 1437 female mosquitoes were captured (159 indoor and 1278 outdoor) and 124 males (5 indoor and 119 outdoor). The endophagous and exophagous activities of Ae. albopictus recorded over 24 h with two-layers mosquito nets at different seasons in La Réunion showed differences at several levels: the time of the year, the hours of the day, and the place of the bait (indoor or outdoor) (Fig. 4). Indeed, the time of the highest activity peak of host-seeking females was depending on the season; it was about 2 h before sunset, that is, at 17 h 30 min in summer (when sunset was at 19 h 00 min) and at 16 h 00 min in winter (when sunset was at about 18 h 00 min). A smaller but important activity peak of female mosquitoes was also registered at 8 h 30 min in every experiment, which coincides with the beginning of the experimentation at 8 h 00 min.

Average number of mosquitoes captured when landing close to human baits on the inner mosquito net during 24 h in the nycthemeral activity experiment (in winter and summer seasons). Graph 1, number of outdoor females; graph 2, number of indoor females; graph 3, number of outdoor males captured according to the time of the day. Note that the three graphs have different y-axis scales. Vertical bars represent standard errors.
The highest percentages of females were collected outside the house (81% and 96% of females caught in summer and winter, respectively). During daytime, from 5 h 00 min to 19 h 30 min, and outside the activity peaks, host-seeking female activity was still observed, but in a lower proportion than during the two peaks. At night, hardly any female was caught inside the house (1, once), but 15% of females were caught outside during the summer and only 1% during the winter. The global exophagic percentage was 89%.
Only five males were caught inside the house during the summer experiments; 93% and 100% of the males were collected outside in summer and winter, respectively. No males were ever caught at night. The highest activity peaks for males were recorded at sunrise (at 6 h 00 min for the summer season and at 7 h 00 min for the winter season) and before the highest activity peak of females.
Resting places of wild Ae. albopictus
As a whole, Ae. albopictus individuals were collected outside or inside in 66% of the 105 visited houses. Among the positive houses, females were collected in 84% of the houses and males in 64%. Over the total number of Ae. albopictus adults caught, we found sevenfold more adults outside (n = 214) than inside (n = 30) the houses. Very few males were caught inside the houses (six houses), whereas 18 houses had females inside, of which only 9 houses had females engorged with blood or were gravid. Forty-two percent of the females caught inside were gravid or blood-fed.
Discussion
Ae. albopictus is the most abundant Aedes species in the Islands of the South West part of the Indian Ocean; in La Réunion, it is present in all human-altered areas up to an altitude of 2000 m in summer and 1200 m in winter (Delatte et al. 2008a). Ae. albopictus was known to be an opportunistic feeder upon most groups of vertebrates, including birds, reptiles, and amphibians, although preferring mammals (Gould et al. 1970, Tempelis et al. 1970, Sullivan et al. 1971, Savage et al. 1993), but because of its opportunistic behavior it was not considered as a good vector to human. Our findings with Ae. albopictus on La Réunion showed that even though the mosquitoes had potentially a high opportunistic feeding behavior on several vertebrate species (nonchoice experiment), when given the choice between a human and an animal bait the human host was by far preferred. However, mixed meals occurred with every animal species. The marked preference for human blood came together with a strong exophagy (outside activity) and exophily (outside resting).
The nonchoice experiment showed that when Ae. albopictus unfed females were put on different vertebrates, they fed on all the tested species ranging from warm-blooded mammals to cold-blooded reptiles, with nevertheless a preference for human, dog, chicken, and cow. In the nonchoice experiment, the percentage of females blood-fed on mammals ranged from 20% to 81%. Surprisingly, Ae. albopictus was able to feed on two cold-blooded predators of insects, geckos and chameleons, at a low rate (2% and 8%, respectively). However, when given the choice between humans and either dogs, chickens, goats, or calves, the females strongly preferred the human host (Fig. 3). Cows, which were found in the nonchoice experiment in the most bitten group by Ae. albopictus, were poorly bitten in the choice experiment compared to humans. The most preferred animal, except humans, in the choice experiment was the goat. To a lower extent, these discrepancies between the nonchoice and choice experiments, especially for the cow and goat animal hosts, might depend on several factors such as the skin odor, convective currents, presence of hairs (the cow was shaved in the nonchoice experiment), skin temperature, physiological status (calf instead of a cow), and movements of animals with mosquito cages on their back (in the nonchoice experiment). Nevertheless, our results strongly suggest that if Ae. albopictus was to have the choice between (equally available) the four vertebrate hosts proposed and a human, the mosquito would preferentially bite the human. This supports earlier data about its high degree of anthropophily from elsewhere in its range, for example, southern Thailand, Macao, and Hawaii (Hawley 1988, Almeida et al. 2005, Effler et al. 2005), and prove that despite its opportunistic feeding behavior it cannot be anymore considered as a poor candidate vector of arboviruses from human to human. In addition, our results are pointing out the ability of Ae. albopictus on La Réunion to have a blood meal despite the lack of its preferred host (human), which might be less nutritive. To further investigate the consequence of feeding on a nonpreferred host, fitness measurement, such as the number of eggs produced and the survival of the offspring depending on the blood nature, should be measured.
A further important result shown by this choice experiment was the propensity of Ae. albopictus to take several blood meals on different hosts, even in a short period of time. This finding is of first interest because it potentially allows the mechanical transmission of arboviruses between different species within a very short period of time. Further, this multiple blood-feeding behavior might also play a role in Ae. albopictus biological parameters. Indeed Rui-De et al. (2008) demonstrated that multiple blood-feeding on two different host species increased the survival of females of Ae. albopictus, so that longer life expectancy and multiple blood feeding are part of the most critical parameters that would enhance an arbovirus spread. Nevertheless, this multiple blood-feeding habit should be checked in the field and its proportion in natural population assessed.
Regarding the field survey realized on La Réunion, Ae. albopictus fed preferentially and rested even more outdoors. Even if captures of resting wild Ae. albopictus are usually considered difficult, if we compare the results obtained in this study on 105 houses, far more adults were captured outside than inside, which enhances the exophilic behavior of Ae. albopictus. The two completely different experiments performed to estimate the exophily (indoor/outdoors captures in 105 houses) and exophagy (24-h captures around a human bait) behaviors of Ae. albopictus showed surprisingly consistent results (i.e., 87% and 89%, respectively). The exophagous feeding behavior had a bimodal distribution peaking in the morning at twilight and 2 h before sunset, which is a common finding with Ae. albopictus (Hawley 1988, Almeida et al. 2005). The late afternoon peak was higher than the morning peak activity in both seasons. The morning peak might be partly explained by the intrusion effect of the human bait in the surroundings (Germain et al. 1972, 1973), despite the precautions made for avoiding it (by installing the mosquito nets the day before). Despite these bimodal peaks of activity Ae. albopictus females had a continuous activity during the 24-h experiments. At night the activity was reduced and not recorded in winter, which can be due to lower night temperatures. In several studies realized on the feeding activity of Ae. albopictus, in rural, urban, or natural habitats, bimodal pattern of feeding activity has been found (Hawley 1988, Almeida et al. 2005). Taylor and Jones (1969) shown that flight activity in Ae. aegypti appears to be controlled by an endogenous rhythm that may be phase-set by both light-on and light-off. In addition, the total activity is correlated with the number of hours of light in a 24-h period. Altogether, light has both a direct effect in determining the amount of activity and an indirect effect through setting the phase of the endogenous rhythm, which agrees with our results. Males were also captured around the human host, which might be explained by a strategy of waiting to copulate with blood-seeking females.
Conclusion and Epidemiological Considerations
In agreement with Savage et al. (1993) we found that Ae. albopictus is an opportunistic feeder that utilizes a wide variety of hosts, with a significant and marked preference for human host. Therefore, it has the potential to become involved in the transmission cycles of indigenous arboviruses but also to increase the number of viral hosts in the case of an arbovirus transmissible to animals. This fact could be particularly the case during a CHIKV epidemic, where animals were found with positive antibodies in several studies (Jupp and McIntosh 1988, Guilherme et al. 1996, Inoue et al. 2003). Ae. albopictus trophic behavior (multiple blood-feeding habit and host range), its ecological plasticity (Delatte et al. 2008a), its tolerance to a wide range of temperatures (Delatte et al. 2009), and its increased range will probably enhance arbovirus circulation and emergence, especially on areas such as African regions where arbovirus sylvatic cycles are common. Recently, Ae. albopictus has been reported from several African countries, such as Nigeria, Guinea, Cameroon, and Gabon (Benedict et al. 2007, Coffinet et al. 2007). Further, all discrepancies found between Ae. albopictus studies on hosts (high variability on host preferences) around the world emphasize the great capacity of Ae. albopictus to host adaptation. Apart from its opportunistic feeding capacity, Ae. albopictus has shown its propensity toward exophagy and exophily; note that Ae. albopictus is called the “forest edge mosquito” (see Hawley 1988) in its native range. Its exophily behaviors might have protected it against the malarial indoor persistent insecticide treatments performed in some countries (Salvan and Mouchet 1994) and might be a reason for its predominance in several countries of the South West Islands of the Indian Ocean (Delatte et al. 2008b).
Epidemiological models have always considered that transmission from human to human was the only way of CHIKV diffusion (Bacaer 2007, Dumont et al. 2008). Our results raise the relevance of addressing other epidemiological models. Indeed, in the case where animals are bitten and able or not to retransmit CHIKV to mosquitoes, it will imply strong modifications of the R0 model (see Introduction). Up to now, according to literature only primates have been shown to multiply the virus, so experimental studies should clarify this point.
Finally, Ae. albopictus trophic behaviors will allow us to improve vector control. Indeed, it showed that indoor sprayings of insecticides will not be efficient to control Ae. albopictus population. Further, the knowledge we gained on its high human-biting behavior and its biting diel cycle will help in promoting personal protection such as the use of repellent and long-sleeve cloths at morning and late afternoon.
Footnotes
Acknowledgments
We would like to thank Fanny Bastien (Institut de Recherche pour le Développement [IRD]), Isabelle Janin (IRD), Jean-Sébastien Dehecq, Direction Régionale des Affaires Sanitaires et Sociales (DRASS), and Philippe Dom (DRASS) for their valuable help in the field work, and P. Gasqui for his help in the statistical analysis. We would also like to thank all the volunteers who stayed still under the double mosquito nets for hours in the diel biting experiments. We also thank Nathalie Becker for correcting the English of our article. This work was partly funded by IRD, DRASS, and the Entomochik and Anichik projects of the Agence Nationale de la Recherché (ANR) Santé Environnement et Sante Travail (SEST).
We thank the CICA-Riviere for loaning the calf for the choice experiment.
Disclosure Statement
No competing financial interests exist.