
Abstract
Ivermectin (IVM) is a chemically modified macrocyclic lactone of Streptomyces avermitilis that acts as a potent neurotoxin against many nematodes and arthropods. Little is known of IVM's effect against either blood-feeding Phlebotomus sand flies, or the infective promastigote stage of Leishmania transmitted by these flies. We injected hamsters subcutaneously with two standard IVM treatments (200 and 400 μg/kg body weight) and allowed cohorts of Leishmania major-infected Phlebotomus papatasi to blood-feed on these animals at various posttreatment time points (4 h, 1, 2, 6, and 10 days). Infected and uninfected sand flies that bit treated and untreated hamsters served as controls. Serum levels of IVM in low- and high-dose–treated hamsters were determined at the five time points. Sand fly mortality following blood feeding was recorded at 24-h intervals and, in relation to IVM treatment, was time and dose dependent. Mortality was most rapid and greatest among infected flies that fed nearest to time of dosing. Mean survival of infected sand flies after feeding on untreated hamsters was 11.5 days, whereas that of infected sand flies that fed 4 h, 1 day, or 2 days posttreatment on high-dose–treated hamsters (400 μg/kg) was 1.6, 2.1, and 2.7 days, respectively. Infected and uninfected sand flies that blood fed 6 days following low-dose IVM treatment (200 μg/kg) still experienced significantly greater mortality (p < 0.02) than controls. Promastigotes dissected out of surviving flies that fed on IVM-treated hamsters showed typical motility and survival. Moreover, 21.7% of IVM-treated hamsters developed lesions after being fed upon by infected sand flies. L. major promastigotes appeared to be tolerant to ng/mL blood levels of IVM that caused significant mortality for up to 10 days posttreatment in blood-feeding P. papatasi.
Introduction
Ivermectin (IVM), a semisynthetic macrolide antibiotic, is an anti-infective drug widely used in veterinary medicine against a variety of parasitic infections (Drummond 1985, Campbell 1989). At sublethal doses, IVM has been found to reduce the life span and fecundity of mites, beetles, horn flies, stable flies, and mosquito species (Miller et al. 1986, Focks et al. 1991, Mahon et al. 1993, Nasr et al. 1996). IVM has been the drug of choice for treatment of onchocerciasis, loaiasis, and lymphatic filariasis (Taylor and Greene 1989, Ottesen et al. 1990, Chippaux et al. 1998) and also shows efficacy against intestinal nematodes (Wen et al. 2008). IVM is relatively nontoxic to most vertebrates, and so it has been used to lower filaremia in humans, thereby significantly reducing the transmission of filariasis (Boatin et al. 1998, Foley et al. 2000). In veterinary medicine, wildlife management, and even in aquaculture, IVM has valuable efficacy against a wide range of parasitic nematodes and arthropods (Campbell et al. 1983, Jackson 1989, Strong 1993, Davies and Rodger 2001, León-Vizcaíno et al. 2001, Geary 2005).
Despite an abundance of published literature on the insecticidal and veterinary properties of IVM, little is known about the effect of this drug on vector sand flies and their obligate Leishmania parasites. Avermectin and IVM sensitivities and lethal concentration (LC)50 of laboratory-colonized adult female Phlebotomus papatasi and P. langeroni have been studied by spiking blood or sucrose solutions with the drugs (Kassem et al. 2001) but sand fly responses over time to blood levels of IVM in treated host animals has not been reported. Similarly, sensitivity of cultured Leishmania promastigotes has been reported from in vitro experiments (Rasheid and Morsy 1998), but in vivo sensitivity of Leishmania within sand flies to IVM acquired by blood feeding on treated host animals has not been described. There are no published reports of IVM effect on Leishmania infections in mammals, or on stages of the parasite developing within the sand fly.
To explore the potential of a two-pronged control strategy that targets both the sand fly vector and Leishmania parasites, we examined whether hamsters pretreated with standard veterinary doses of IVM could develop serum levels of the drug that adversely affect blood-fed sand flies, as well as the developing stages of Leishmania within these sand flies.
Materials and Methods
Sand flies
A laboratory colony of P. papatasi established from collections in the North Sinai, Egypt, during 1989, and maintained in the U.S. Naval Medical Research Unit No. 3 insectary was used in all phases of this study.
Animals
Adult Syrian golden hamsters (Mesocricetus auratus), weighing 100 ± 30 g, were used in these experiments. All animals used in this work were cared for and maintained under conditions approved by the Association for Assessment and Accreditation of Laboratory Animal Care International.
Ivermectin
The IVM used in this study was a commercial injectable formulation of IVEEN® labeled for veterinary use (reg. no. 926/95 Vet.) and provided by the Egyptian Company for Chemicals and Pharmaceuticals (ADWIA). This formulation is a sterile solution containing 1% IVM, 40% glycerol, 15% benzyl alcohol, and 44% propylene glycol. Per manufacturer's label instructions the formulation was diluted with propylene glycol for subcutaneous (SC) injection.
Leishmania parasites
The Leishmania major strain (IPAP/EG 89/SI-177) used in this study was isolated in 1989 from wild-caught P. papatasi collected from North Sinai, Egypt, (Mansour et al. 1991) and maintained by serial passage and liquid nitrogen cryopreservation. Cryopreserved parasites were thawed at room temperature and grown to stationary phase in Schneider's Drosophila medium. After 4–5 days, promastigotes were harvested, washed three times in normal phosphate-buffered saline (PBS), and centrifuged at 3000 rpm for 10 min. Washed parasites were resuspended in equal volume of PBS and counted by hemocytometer to determine the number of parasites/mL. Approximately 20 μL (equivalent to ∼6 × 106–7 parasites/mL) of suspension were inoculated into the hind foot-pads of BALB/c mice to produce lesions. After lesions appeared, mice were euthanized and the infected lesions were extirpated and homogenized in PBS. This suspension was mixed with an equal volume of washed hamster blood and used for infecting sand flies via membrane feeding.
Infection of sand flies with Leishmania
Female sand flies (150–200) were fed on the amastigote-blood suspension through chick-skin membrane using water-jacketed glass feeders warmed to 37°C. Blood-fed flies were held for 6–9 days at 27°C with 90% relative humidity, a 12:12 light:dark photoperiod, and provided with 30% sucrose food solution. After 7–10 days of parasite development, promastigote-infected sand flies were fasted for 24 h in preparation for direct feeding on IVM-treated hamsters.
IVM treatment of hamsters and sand fly feeding
The selected 200 and 400 μg/kg body weight dosages of IVM and SC administration followed that of Valarie and Tynes (1998). In accordance with the manufacturer's instructions for control of endo- and ectoparasites in rodents, and to minimize variability associated with oral dosing, each of 10 hamsters was treated with a single SC dose of IVM at 200 μg/kg body weight. A second group of 10 hamsters received the IVM 400 μg/kg dose. At 4 h, 1, 2, 6, and 10 days post-IVM treatment, hamsters were fed upon by uninfected and Leishmania-infected sand flies. Prior to blood-feeding, hamsters were sedated with ketamine (50 mg/kg, IM), and their bellies were shaved to facilitate sand fly blood-feeding. After 30 min all blood-engorged flies were counted and apportioned into two equal groups: one in which mortality was monitored daily, the other from which subgroups were taken for dissection in PBS to determine the appearance and motility of their Leishmania promastigotes. Age-matched infected and uninfected sand flies that fed on untreated hamsters served as controls. Hamsters were visually inspected for developing lesions twice weekly for up to 4 months. Lesions that appeared were aspirated onto slides, air-dried, stained with Giemsa, and examined for the presence of Leishmania amastigotes via 400 × light microscopy.
Effect of IVM on cultured promastigotes
Aliquots (1.0 mL) of counted culture promastigotes were transferred to media containing seven different concentrations of IVM (50, 100, 150, 200, 250, 500, and 750 μg/mL). Samples drawn at 5 min, 1, 2, and 24 h postexposure were placed onto a Neubauer counting chamber and assessed by light microscopy for motility, appearance, and density. Additionally, the hind footpads of two BALB/c mice were inoculated with 0.02 mL of promastigotes taken after 24 h from culture media containing 50 μg/mL of IVM. The two mice were monitored daily for signs of pathology.
Extraction and measurement of IVM drug levels in hamsters
At 4 h, 1, 2, 6, and 10 days post-IVM treatment, 1.0 mL of blood was taken from treated hamsters. The blood was permitted to clot, after which sera were collected and stored at −20°C for later analysis. Extraction and quantitative determination of serum IVM by high-performance liquid chromatography (HPLC) followed the method of Alvinerie et al. (1995), with slight modification. In brief, a 50 μL serum sample was fortified with 25 μL internal standard IVM (100 ng/mL) and then mixed with 525 μL acetonitrile to deproteinize the serum. The sample was vortexed for 2 min and then centrifuged for 15 min at 5000 rpm. The supernatant was loaded onto a solid phase extraction column SPE-C18 (1 cc, 200 μg/mL; Chromaband® C18, Phenomenex, Torrance, CA) that had been equilibrated with 5 mL acetonitrile at a flow rate of 1 mL/min, followed by a 5 mL acetonitrile/water mixture (30:70) containing 100 μL triethylamine at the same flow rate. The supernatant was loaded at a flow rate of 0.5 mL/min after which the solid phase extraction column was allowed to dry via gentle suction at the same flow rate for 2 min. Bound IVM in the column was eluted with two 500-μL passages of pure acetonitrile at 0.25 mL/min. The eluent was collected in an Eppendorf tube and allowed to dry at 60°C under a gentle nitrogen stream. The dry eluent/residue was redissolved in 50 μL pure acetonitrile, followed by the addition of 50 μL diluted N-methyl imidazole (1:1, imidazole:acetonitrile) as a buffer, and 50 μL diluted trifluoroacetic anhydride (1:1, trifluoroaceticanhydride:acetonitrile) to initiate derivatization. This mixture was allowed to react for 1 min to complete derivatization, and then 20 μL of the derivatized sample was injected into a HPLC unit (model LC 10 ADVP; Shimatzu, Kyoto, Japan). Analysis was undertaken isocratically using a reverse phase C18 column (Phenomenex, 250 × 5.2 mm, 10 μm), and an acetonitrile/methanol/water mixture (70:25:5) as a mobile phase at a flow rate of 1 mL/min. The derivatized IVM was detected using a fluorescence detector (model RF-10AXL; Shimatzu, Kyoto, Japan) adjusted to 365 and 470 nm to encompass excitation and emission wavelengths and connected to a Shimatzu laboratory computing integrator (model C-R8A; Shimatzu, Kyoto, Japan). The area of the eluted peak was a function of the concentration of IVM in the sample. The lower level of detection reported for this method is 1 ng/mL.
Data analysis
Data were analyzed with statistical software SPSS Version 10.0 for Windows©. Analysis of variance was used to compare mean times to mortality among blood-fed sand flies that fed upon low-dose–treated, high-dose–treated, and untreated hamsters, and at 4 h, 1, 2, 6, and 10 days post-IVM treatment. Posttesting compared between treatments and times within treatments. Differences between groups in the proportions of flies that blood-fed were compared by chi-square test.
Results
IVM blood levels in hamsters and Leishmania infection
Treatment of hamsters by SC injection with either high or low doses of IVM resulted in serum levels of the drug that were detectable by HPLC over the 10 days of sampling (Fig. 1). Highest serum levels, which averaged 80.2 ng/mL, were measured 4 h posttreatment in the high-dose group. At this same posttreatment time point, highest average serum level in the low-dose group was only 16.3 ng/mL. Low- and high-dose SC treatment levels differed only by a factor of 2, but mean serum levels measured in the low-dose group ranged from 4.7 to 8.6 times lower at the five time points. There was indication of an elimination half life (t ½e) for the 200 μg/kg low dose of <2 days, with little difference between days 2, 6, and 10 in the detectable serum level of drug. Half life of the higher dose was not clear from our sampling points, but little difference was seen between levels on days 6 and 10 posttreatment. Despite differences in mean serum levels of IVM measured in the two treatment groups, the percent decline within groups, from peak levels at 4 h after treatment, until 10 days posttreatment, was comparable and amounted to 64.3% and 62.6%, respectively, for the high- and low-dose groups.
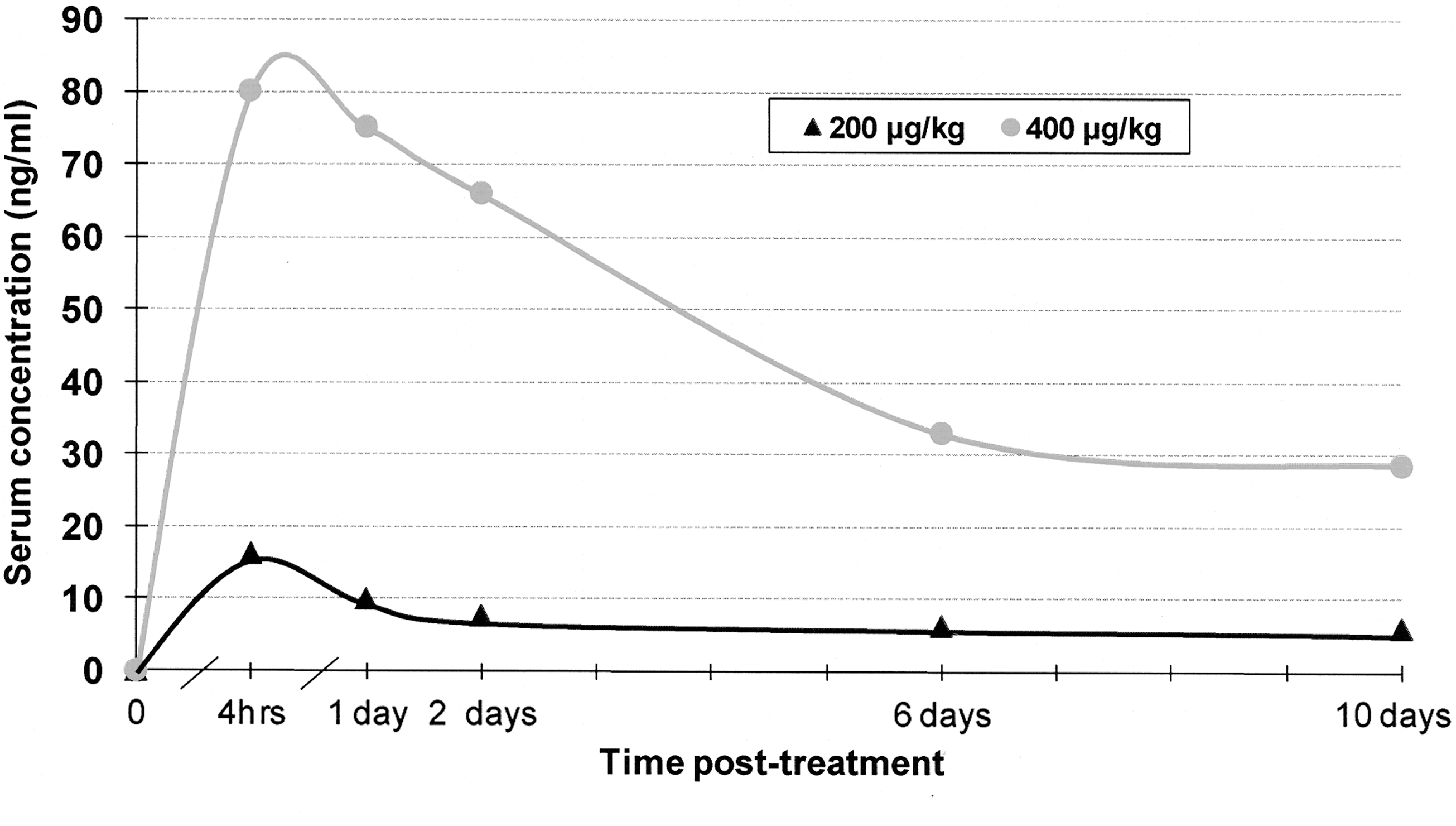
Ivermectin (IVM) serum levels in hamsters: 4 h, 1, 2, 6, and 10 days after 200 or 400 μg/kg subcutaneous (SC) treatment.
L. major tolerance in vivo and in vitro to IVM
On dissection, high promastigote infection rates were found for sand flies that bit treated hamsters at each of the five posttreatment time points. Lesions with confirmed L. major amastigotes developed in 2/11 (18.2%) low-dose–treated hamsters bitten on day 2 posttreatment when serum IVM measured 7.7 ng/mL, and in 3/12 (25%) high-dose–treated hamsters bitten on days 1 (one infection) and 2, when IVM serum levels ranged from 66.1 to 75.2 ng/mL. Lesions in the low-dose group appeared on days 29 and 46 postbiting and in the high-dose animals on days 27, 29, and 56. There was no apparent difference in the occurrence and timing of lesions that developed in untreated control hamsters (one of five hamsters developed lesions on day 67; organs were not tested using PCR to document infections in the other four control animals) that were bitten by infected flies from the same batch.
Culture promastigotes of L. major exposed, in vitro, to IVM concentrations ranging from 100 to 750 μg/mL demonstrated rapid mortality within 2 h and 95–100% mortality by 24 h (Fig. 2). However, promastigotes exposed to IVM at 50 μg/mL, a concentration >600 times higher than the peak serum level (80 ng/mL) measured in hamsters treated with the SC high dose, suffered only slight mortality during the first 2 h of in vitro exposure, and ∼54% mortality after 24 h (Fig. 2). Further evidence of tolerance was seen in the development of lesions in BALB/c mice within 20 days after inoculation of cultured promastigotes that had been exposed, in vitro, for 24 h to this 50 μg/mL IVM concentration.
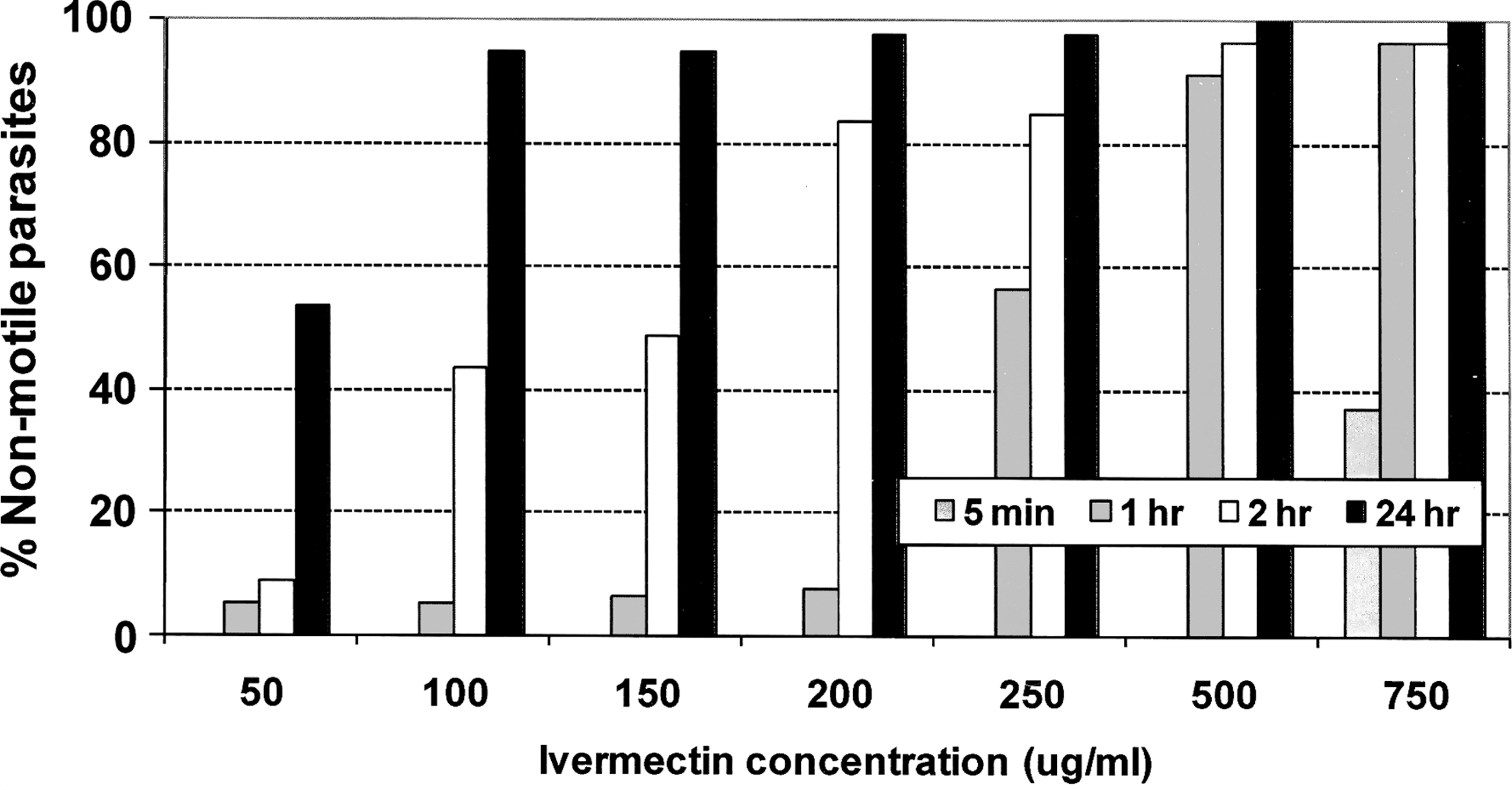
Effect of IVM concentration and exposure time on Leishmania major promastigotes in culture.
Sand fly responses to IVM acquired by blood feeding on treated hamsters
P. papatasi mortality in response to blood feeding on IVM-treated animals was dose and time dependent (Figs. 3a, b and 4a, b). In terms of dose dependence, sand flies that bit animals treated with high IVM dose experienced significantly earlier and greater mortality (p < 0.05) than flies that bit untreated and low-dose–treated hamsters. On each day following blood feeding the cumulative mortality rate of flies that bit high-dose–treated hamsters was consistently higher for each posttreatment blood feeding group than in corresponding groups of flies that bit low-dose–treated hamsters. In relation to both the low- and high-dose IVM treatments, the steepest slope on mortality rate curves, indicating most rapid death, occurred within 2 days following blood feeding on serum levels that ranged from 7.7 to 80.2 ng/mL. For the high-dose IVM treatment, toxic response of infected sand flies was initially slow, with mortality of 20–37% measured at 24 h and mortality ranging from 65% to 100% at 48 h. For the lower IVM dose, and again among the Leishmania-infected flies, this lag was even more pronounced (Fig. 4a, b). This delayed response was not apparent in uninfected sand flies (Fig. 3a, b). Cumulative mortality plots showed that relative to controls, IVM levels in blood 6 days after low-dose treatment had toxic effects against both infected and uninfected flies, but by day 10 there was no observed toxic effect (Figs. 3a and 4a). IVM levels 10 days after high-dose treatment appeared to be toxic only to infected flies (Fig. 4a, b).
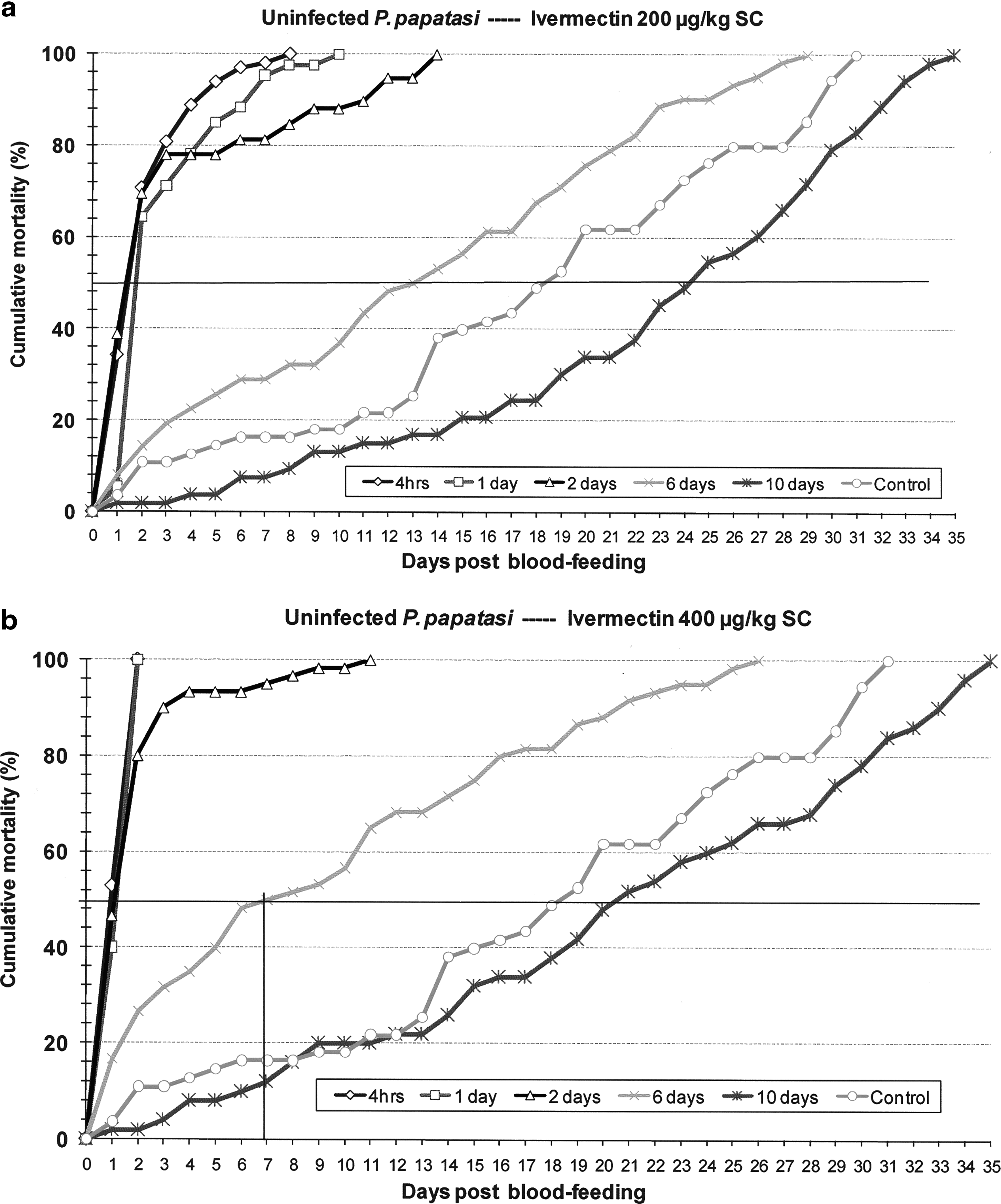
Cumulative daily mortality of uninfected Phlebotomus papatasi after blood feeding on hamsters treated with IVM (
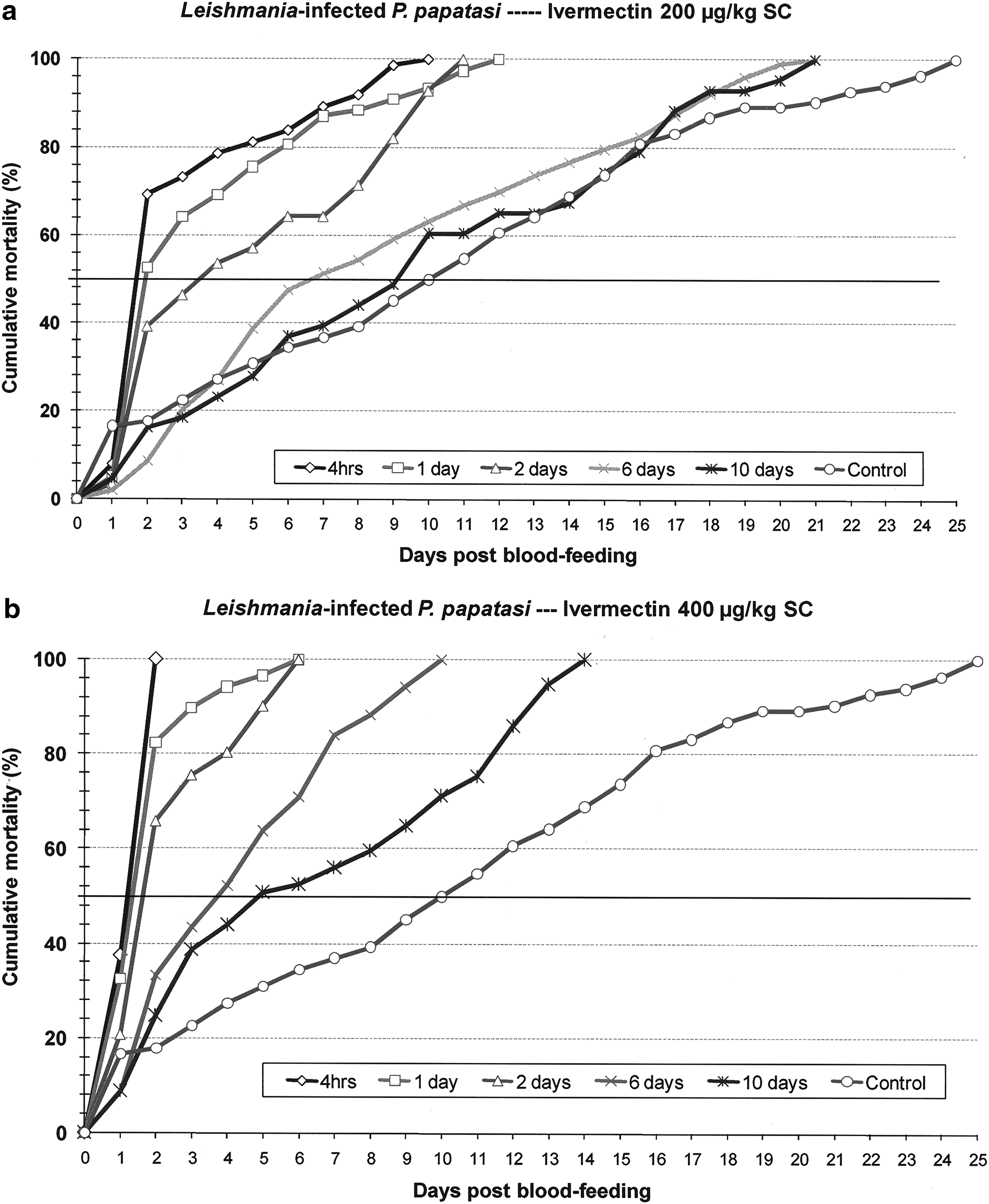
Cumulative daily mortality of Leishmania-infected P. papatasi after blood feeding on hamsters treated with IVM (
Comparison of Leishmania-infected and age-matched uninfected sand flies fed on untreated hamsters showed that mean survival time was significantly reduced by the L. major infection (11.5 vs. 18.7 days; p < 0.005). Complete mortality in these respective control groups was attained on days 25 and 31 post-blood feeding. This effect of the parasite did not translate to consistently reduced mean survival times among groups of flies that fed on IVM-treated hamsters. Despite significantly reduced survival of infected flies fed on untreated control hamsters, both infected and uninfected flies demonstrated acute, and fairly comparable, sensitivity to IVM blood levels that ranged from 7.7 to 80.2 ng/mL (Tables 1 and 2). Mean survival time, equivalent to lethal time (LT)50, of infected sand flies that bit treated hamsters within 2 days of IVM dosing often exceeded that of uninfected flies, but among groups of flies that blood fed 6 and 10 days posttreatment, when IVM levels ranged from 6.1 to 33 ng/mL, mean survival times of uninfected flies greatly exceeded those of infected flies (Tables 1 and 2). Sand fly blood-feeding avidity on IVM-treated hamsters also differed between infected and uninfected sand flies. No difference was observed between feeding rates on low- and high-dose–treated hamsters, but blood feeding rates for uninfected flies was significantly greater than that of Leishmania-infected flies (low dose: 93.7% vs. 81.6%, p < 0.005; high dose: 93.0% vs. 82.3%, p < 0.005).
IVM, ivermectin.
Discussion
Our results provide a body of evidence from which a potential role for IVM in the control of sand flies can be considered. Key but opposing elements revealed were the clear impact upon P. papatasi survival produced by nanogram levels of IVM in the blood of hamster for up to 10 days posttreatment and the high level of tolerance to this drug by both the promastigote and amastigote stages of L. major. Independent of IVM, but with significant impact upon vector survival, we also demonstrated the deleterious effect of parasitism by L. major on the infected sand fly host.
The effect of IVM, in vivo, on our infected and uninfected sand flies appears to differ from that obtained by membrane feeding uninfected, colony-reared P. papatasi with IVM-spiked blood (Kassem et al. 2001). We observed that the infected sand fly mortality associated with blood feeding on IVM serum levels of 7.7–16.3 ng/mL was negligible at 24 h but rose to 38–70% by 48 h. Similarly, for the high-dose treatment, groups of sand flies that blood fed on IVM serum levels of 66–80 ng/mL experienced 24-h mortality that ranged from 35 to 38%, but which rose to 65–100% by 48 h. This curious lag or delayed response to IVM that was seen in six separate cohorts of infected sand flies was unexpected. Based on our determination of significantly reduced survival of Leishmania-infected sand flies, relative to uninfected ones, we had anticipated greater and/or prolonged sensitivity of parasitized flies to IVM, that is, toxic insult beyond tissue damage and nutritional competition. It seems possible that IVM in our in vivo system had bound to plasma proteins and fats in the hamster's blood in such a way as to render it less available than IVM simply spiked into blood for immediate membrane feedings (Klotz et al. 1990). Also important to consider is the likelihood of damage and blockage or barrier effect in the anterior midgut of the flies caused by Leishmania infection, particularly when flies are heavily infected (Schlein et al. 1992, Stierhof et al. 1999, Rogers and Bates 2008). We observed no comparable delay in uninfected blood-fed sand flies that might credit slowed IVM tissue exposure to the peritrophic membrane. The orderly dose- and time-dependent mortality which did occur in our infected flies would seem to attest to blood-feeding success. In comparing between age-matched, infected and uninfected sand flies that fed on untreated hamsters, we observed clearly that the mean survival time of infected flies was significantly lower and thus interpreted this difference to derive from the deleterious effects of their parasite burdens (Rogers and Bates 2008). In this respect, the reduced sensitivity of our infected sand flies and their delayed mortality responses relative to uninfected flies that fed on the same IVM-treated hamsters would seem even more difficult to reconcile. In nature, L. major infections within P. papatasi are predominantly light, and as a result, little or no impact of survival of the vector is deduced (Yuval 1991). Our unnatural laboratory infections, maximized and induced to ensure high frequency and heavy infections in all fed flies, may have produced levels of promastigote infection far exceeding those typically encountered in nature. As such, parasite-induced blockage, barrier effect, competition, and immune responses may have been the main determinants contributing to the delayed toxic response that was observed in our infected sand flies. At lower parasite densities these parasite-mediated conditions would be expected to result in more effective transmission of promastigotes to our IVM-treated hamsters. Although lesions appeared in more than 22% of the treated hamsters, we observed lesions develop in only one of five untreated control hamsters. The high mortality and low transmission lead us to question whether we overinfected our flies. In hindsight, PCR may have successfully demonstrated the presence of parasite DNA in those hamsters that showed no signs of pathology. Alternatively, these induced, maximized infections may have been too large to enable normal transmission by bite.
Our results demonstrate that P. papatasi, one of the most important sand fly vectors of human disease in the Old World, is sensitive to ng/mL levels of IVM that resulted from SC treatment with two standard veterinary doses of the drug. Unfortunately, these relatively large doses and the optimal SC delivery route produced serum levels that had an appreciable toxic effect against uninfected flies of only 6 days. Wild populations of sand flies, lacking the vigor and robustness of colony-reared flies, may be far more sensitive, and as seen with numerous arthropod species, sublethal levels of IVM reduce egg formation, hatch rates, and general fecundity.
SC dosing is not practical for control in the zoonotic cycle, but a more sensible delivery of IVM by oral and percutaneous dosing might be possible with IVM-treated baits or spraying of burrows and preferred food plants. Although these methods could not expect to achieve serum drug levels comparable to SC dosing, and outright lethality to the biting flies, they could, by virtue of a gradual feed-through effect, deliver targeted IVM levels through feces that have profound and potentially long-term toxic effect against the more vulnerable larval stages of the sand flies. Strong (1993) drew attention to feces as the main elimination route of avermectin and the broad adverse effects of the active drug residues on pastureland ecology, but Mascari et al. (2008) demonstrated serious potential of this approach for Middle Eastern Leishmania control. Hamsters fed a diet containing 20–100 ppm of IVM excreted sufficient levels of the active drug in their feces to cause high mortality when fed to P. papatasi larvae.
We did not design our study to measure pharmacokinetics of IVM dosing in hamsters, but the peak serum levels and profile that we determined in these animals from sparse data points after the recommended SC dosing was comparable to those reported for a variety of other mammalian species. IVM pharmacokinetics are reported to differ markedly among species, and to vary according to formulation, route of administration, sex, age, and physiological status (Cerkvenik and Grabnar 2002). Among small mammals, IVM-treated rabbits and rats are reported to develop high, sustained blood levels of IVM for >13 days, whereas higher doses (500 and 1000 μg/kg) in jirds and guinea pigs are almost undetectable after 48 h (McKellar et al. 1992, Sithithaworn et al. 1998).
Nematode and arthropod species also differ widely in their degree of susceptibility to IVM, but higher dipterans are considered to be particularly sensitive. Foley et al. (2000) reported that blood feeding by Anopheles farauti on human volunteers treated with 250 μg/kg IVM, which would have produced blood levels >46 ng/mL, caused 80–100% mortality within 3 days. Mass treatment of IVM for filariasis control in Papua New Guinea produced blood levels that were toxic to mosquitoes of the An. punctulatus group (Bockarie et al. 1999). Studies by Pampiglione et al. (1985) that compared between three mosquito species found the insecticidal effect of IVM to be greater against anophelines than culicines. Follow-on studies that compared IVM susceptibility between Aedes aegypti, Ae. albopictus, and Culex quinquefasciatus found Culex to be most tolerant and calculated that the fecundity of both Aedes species was reduced 50% by IVM blood levels of only 3–4 ng/mL (Tesh and Guzman 1990). Psoroptic mange mites of cattle were similarly susceptible, with fecundity reduced to 50% by blood levels of IVM remaining 24 days after a single 200 μg/kg SC treatment (Guillot et al. 1986). Kassem et al. (2001) reported an LC50 of IVM in P. papatasi of 13 ng/mL, a level more than 50 times and more sensitive than Cx. quinquefasciatus, but found that although sublethal levels of the drug reduced the number of sand fly eggs laid, there was no apparent effect on their hatch rate relative to untreated controls.
Because of the broad efficacy of IVM over many invertebrate species, ranging from protozoa to higher insects, our study also sought to determine whether infective-stage Leishmania promastigotes inside the sand flies would show signs of toxicity associated with IVM blood levels produced by standard SC dosing, and secondarily, whether these IVM levels in the hamster would prevent transmitted parasites from developing into amastigote infections. The absence of any change in the motility and appearance of promastigotes in the blood-fed flies, and the development of multiple infections in both low- and high-dose–treated hamsters suggested that IVM blood levels were ineffective. Our in vitro evaluations, which exposed promastigotes to grossly higher levels of IVM, indicated a tolerance by L. major far beyond blood levels that could be safely achieved by public health and veterinary regimens of this drug. In their important review, Campbell et al. (1983) attributed the IVM insensitivity of flukes, tapeworms, and protozoa to the critical absence of gamma-aminobutyric acid–mediated neurotransmission in these organisms. Although there are reports of IVM efficacy, in vivo, against some species of intestinal protozoa (Njoo et al. 1993, Youssef et al. 1996, Hassan et al. 2001), other in vitro-based investigations have reported very low IVM activity against Leishmania and Plasmodium (Rasheid and Morsy 1998, Nasveld et al. 2003). In retrospect, high-level tolerance to IVM by L. major is not surprising. IVM was in wide use throughout the United States as a regular treatment for the prevention of heartworm in dogs when Gaskin et al. (2002) reported L. infantum infections in 42% of foxhounds in a New York Kennel. When it was also shown that canine leishmaniasis affected foxhounds in 17 other states and 2 Canadian provinces (Duprey et al. 2006), it became obvious that L. infantum was unaffected by long-term exposure to levels of IVM that conferred near absolute protection against Dirofilaria immitis.
In summary, although blood-feeding P. papatasi is highly sensitive and susceptible to levels of IVM that can be attained in blood for 6–10 days by SC dosing, we observed no lethal or suppressive effect upon promastigotes of L. major inside these sand flies, or amastigotes within the IVM-treated hamsters. The relatively low IVM blood levels that were attained in hamsters, combined with the short half-life of the drug, limit the duration of toxic effect in blood-feeding sand flies. Further study is needed to determine whether IVM can be delivered to wild rodents in acceptable oral formulations that generate feed-through levels of IVM in feces, with greater focus and impact on the larval stages of these vectors.
Footnotes
Acknowledgments
The authors thank Fetouh Ali and Ahmed Fawzy of the U.S. Naval Medical Research Unit No. 3 Vector Biology Research Program for their technical support, and are also grateful to Ms. Maria Badra for reviewing and processing this manuscript. This work was supported by the Military Infectious Diseases Research Program, Work Unit No. 6000-RAD1-U-E0305-U0005-04-N3.
Disclaimer
The views expressed in this article are those of the authors and do not necessarily reflect the official policy or position of the Department of the Navy, Department of Defense, or the U.S. Government. The experiments reported herein were conducted in compliance with the Animal Welfare Act and in accordance with the principles set forth in the “Guide for the Care and Use of Laboratory Animals,” Institute of Laboratory Animal Resources, National Research Council, National Academy Press, 1996, and under U.S. Naval Medical Research Unit No. 3 Animal protocol No. 04-04.
Disclosure Statement
Four coauthors are military service members, the first author and another coauthor are employees of the U.S. Government. This work was prepared as part of our official duties. Title 17 U.S.C. §105 defines that “Copyright protection under this title is not available for any work of the United States Government.” Title 17 U.S.C. §101 defines a U.S. Government work as a work prepared by a military service member or employee of the U.S. Government as part of that person's official duties. No competing financial interests exist.