
Abstract
The tick Ixodes pavlovskyi is taxonomically, morphologically, and ecologically close to the taiga tick Ixodes persulcatus, a major host and vector of Borrelia spirochetes. The recent range of I. pavlovskyi is disjoined into the Western Siberian and Far Eastern parts, with this tick being almost always sympatric with I. persulcatus. A total of 56 unfed adult I. pavlovskyi ticks from the biotope where this species was absolutely dominant (within the city limits of Tomsk, Western Siberia) and 50 I. persulcatus ticks from the vicinity of this city, where I. pavlovskyi was almost absent, were collected by flagging in May–June 2006, at the seasonal peak of their abundance. The guts and internal organs of individual ticks were inoculated into the Barbour-Stoenner-Kelly (BSK) medium. Thus, 35 Borrelia isolates were obtained and identified by means of polymerase chain reaction–restriction fragment length polymorphism analysis and sequencing of the rrfA–rrlB spacer and, selectively, an rrs gene fragment. The Borrelia infection rate in I. pavlovskyi (35.7 ± 12.8%) was almost the same as in I. persulcatus (30.0 ± 13.0%). Such a high infection rate in I. pavlovskyi shows that this vector can itself maintain natural foci of borreliosis, regardless of very low abundance or even absence of I. persulcatus. In both foci compared, Borrelia garinii prevailed in ixodid ticks (31 isolates). Three B. afzelii VS461 isolates were obtained from I. pavlovskyi. Therefore, independently of whether I. pavlovskyi or I. persulcatus is the main vector, B. garinii of two genomic groups, 20047 and NT29, can circulate in a natural focus, but B. garinii NT29 is more closely associated with I. persulcatus. Moreover, two isolates (one from I. pavlovskyi and one from I. persulcatus) proved to be completely identical to B. garinii ChY13p from I. persulcatus collected in China. The hypothesis is that these and other similar isolates described previously comprise an individual genomic group of B. garinii.
Introduction
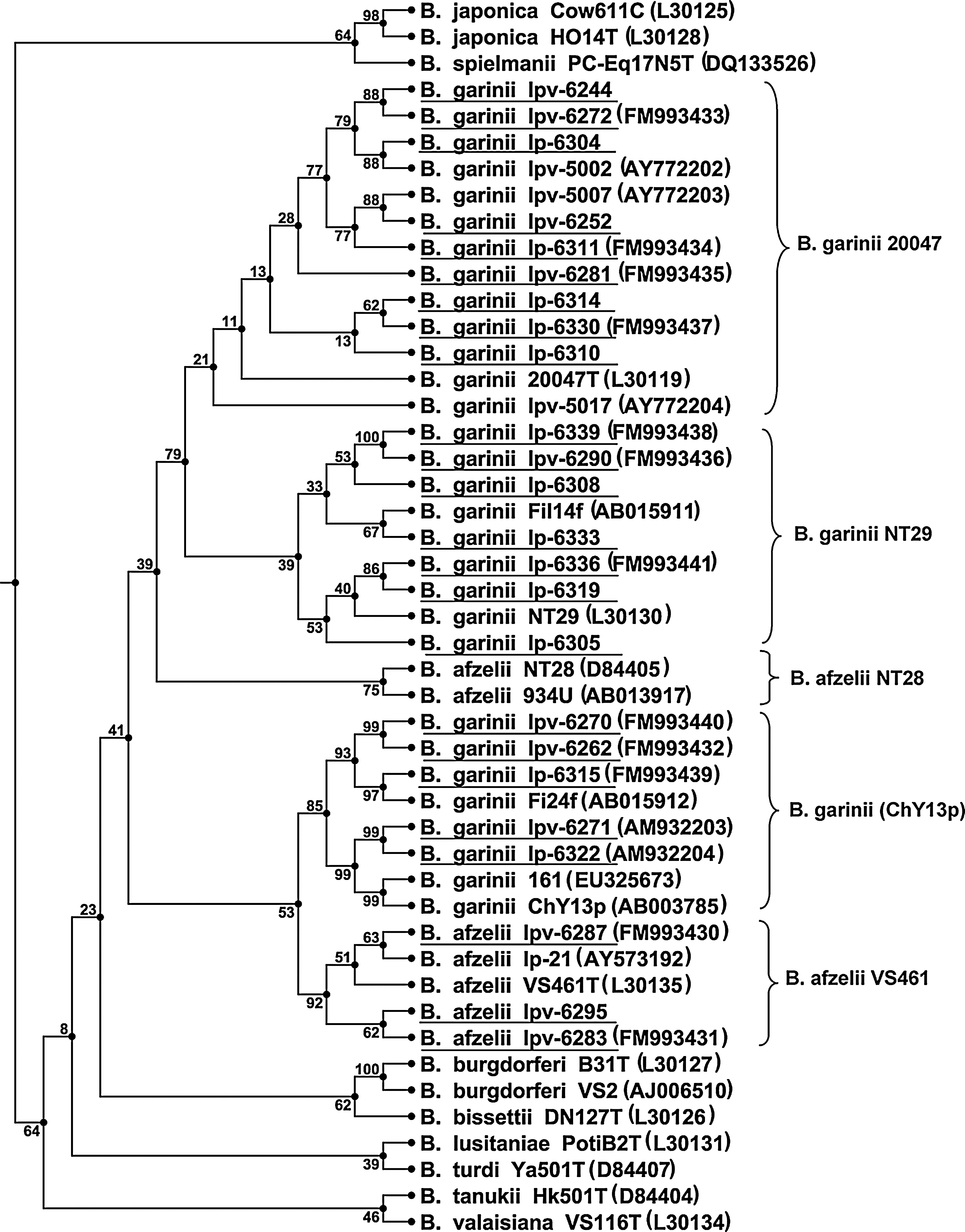
Dendrogram of similarity in the rrfA–rrlB spacer sequence between Borrelia burgdorferi sensu lato and 24 isolates (underlined) from Ixodes pavlovskyi and I. persulcatus ticks collected in the city of Tomsk and its vicinity, Western Siberia. For designations of isolates, see Isolation of Borrelia section in text; GenBank/EMBL/DDBJ accession numbers are shown in parentheses.
A few years ago, permanent populations of I. pavlovskyi ticks were revealed along with I. persulcatus populations in several mutually isolated biotopes within the city limits of Tomsk, Western Siberia (Romanenko 1999, 2005a, 2005b, Romanenko and Chekalina 2004). A phase-contrast microscopic analysis showed that ticks of both species were infected by Borrelia (Romanenko and Pankina 2005). Here, we present the results of the study aimed to isolate Borrelia from I. pavlovskyi and I. persulcatus ticks collected in Tomsk, to genotype the resulting isolates, and to compare the diversity of Borrelia in the two tick species.
Materials and Methods
Study area
Field work was done in two distant biotopes in which different tick species prevailed. Biotope 1 was a park forest (an old cemetery) in the southern outskirts of the old, large city of Tomsk, which lies in the extreme southeast of the West Siberian Plain (56°30′N, 84°58′E), in the zone of transition from dark taiga and pine forests to small-leaf forests. The pattern of vegetation in this biotope was close to a natural open, mature, mixed forest with a diverse herbaceous layer. In late May 2006, when ticks for this study were collected, the abundance of adult I. pavlovskyi reached 50 individuals/km (ind./km), whereas that of I. persulcatus was much lower (only 1–2 ind./km). Biotope 2, which included young pine and deciduous forest areas with well-developed herb–shrub layer, was 12 km southeast of Tomsk (Romanenko 2007). In contrast to biotope 1, the abundance of taiga ticks in it was high (up to 61 ind./km in early June), whereas I. pavlovskyi ticks were absent, although single individuals of this species had occurred there in previous years.
Ticks
On the whole, 106 unfed adult ticks—21 female and 35 male I. pavlovskyi from biotope 1, and 17 female and 33 male I. persulcatus from biotope 2—were collected by flagging in May–June 2006, at the seasonal peak of their abundance. Only adult ticks were collected, because the larvae and nymphs of these species are attached to the flag very rare.
Isolation of Borrelia
Live ticks were dissected individually. The gut and internal organs of each tick were inoculated into the Barbour, Stoenner, Kelley-medium (BSK) medium (Sigma, St. Louis, MO), and the cultures were incubated at 33°C for up to 30 days. A total of 35 isolates were obtained and designated according to the source tick species, Ipv (from I. pavlovskyi) or Ip (from I. persulcatus). These isolates are stored at the Museum of Borrelia in the Laboratory of Vectors of Infections, N.F. Gamaleya Research Institute for Epidemiology and Microbiology, Moscow.
Identification of isolates
All Borrelia isolates were identified by means of polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) analysis and sequencing of the rrfA–rrlB spacer region and a fragment of the rrs gene. DNA was isolated with commercial Proba-NK kits (DNK Tekhnologiya, Moscow, Russia). The rrfA–rrlB spacer was PCR-amplified and sequenced with primers IGSb1 (5′-AGCTCTTATTCGCTGATGGTA-3′) (direct) and IGSa2 (5′-CGACCTTCTTCGCCTTAAAGC-3′) (reverse) (Derdakova et al. 2003).
Amplification procedure, carried out in a Tertsik thermal cycler (DNK Tekhnologiya), involved initial denaturation at 95°C for 5 min and 30 cycles of denaturation at 94°C for 1 min, annealing at 65.8°C for 1 min, and elongation at 72°C for 2 min. The amplicons were purified using a Wizard PCR Preps DNA Purification System (Promega, Madison, WI) and treated with Tru1I (MseI) restriction enzyme (MBI Fermentas, Vilnius, Lithuania) as recommended by the manufacturer. Restriction fragments were resolved by electrophoresis in 16% polyacrylamide gel (19:1) (Bio-Rad, Richmond, CA) in TBE buffer at 100 V for 8 h. To estimate their size, the pUC19 DNA/MspI (HpaII) Marker 23 (MBI Fermentas) was loaded on each gel. The gels were stained with ethidium bromide (5 μg/ml) and photographed in UV light using a DNA Analyzer Videosystem (Moscow, Russia). To confirm the results of genotyping for some isolates, additional amplification of an approximately 1500-bp fragment of the rrs gene was performed with primers 16Sf1 (5′-ATAACGAAGAGTTTGATCCTGGC-3′) (direct) and 16Sr (5′-CAGCCGCACTTTCCAGTACG-3′) (reverse) (Masuzawa et al. 1999). The amplification procedure involved initial denaturation at 95°C for 5 min; 35 cycles of denaturation at 93°C for 1 min, annealing at 50°C for 1 min, and elongation at 72°C for 1 min; and final elongation at 72°C for 9 min. For more complete sequencing of the rrs fragment, the procedure was performed with primers covering different regions of this gene (Le Fleche et al. 1997): 16Sf1 and S5 (5′-GAGGAATAAGCTTTGTAGGA-3′) (442–462 bp), S9 (5′-ATTAGATACCCTGGTAGTCT-3′) (784–803 bp), or S13 (5′-AGGAAGGTGAGGATGACGTC-3′) (1169–1188 bp).
The sequencing of the rrfA–rrlB spacer in 24 isolates and the rrs gene fragment in several isolates using an ABI PRISM BigDye Terminator v.3.1 reagent kit (Applied Biosystems, Lincoln, CA) and the analysis of reaction products in an ABI PRISM 3100-Avant Genetic Analyzer (Applied Biosystems) were performed at the Genome Collective Access Center (Engelhardt Institute of Molecular Biology, Russian Academy of Sciences, Moscow, Russia;
Results
Borrelia were isolated from approximately one-third of all ticks collected. It is noteworthy that the infection rate of I. pavlovskyi ticks from biotope 1 (where the abundance of I. persulcatus was very low) proved to be almost the same as that of I. persulcatus (the main Borrelia vector in the greater part of Eurasia) from biotope 2, where I. pavlovskyi ticks were absent: the difference in these parameter is statistically insignificant (Student's distribution: td = 0.65) (Table 1).
According to the results of PCR-RFLP analysis, the greater part of all isolates (31 out of 35) obtained from both tick species were B. garinii. However, 15 out of 20 isolates from I. pavlovskyi were B. garinii 20047, whereas spirochetes of this subgroup in I. persulcatus were no more frequent than B. garinii NT29. It is relevant to note in this context that three isolates from I. pavlovskyi collected in the southwestern Altai (Gorelova et al. 2001) also belong to subgroup B. garinii 20047.
Only one isolate from ticks of each species was identified as B. garinii (ChY13p). Only three isolates, all from I. pavlovskyi ticks, were classified as B. afzelii (Table 1). This Borrelia genospecies is less common in the study region, but it would have also been revealed in I. persulcatus ticks if the number of inoculations was greater.
The sequencing of the rrfA–rrlB spacer in 24 isolates from the Tomsk region, the degree of homology between the resulting sequences, and their comparison with the sequences deposited in GenBank/EMBL/DDBJ databases confirmed the results of Borrelia genotyping by the PCR-RFLP method. The spacer sequence in isolates classified as B. garinii 20047 (Fig. 1) showed the highest homology (97.7–99.2%) to that in the type strain of this subgroup (GenBank accession no. L30119); in another group of B. garinii isolates, this sequence proved to be 99.2–100% homologous to that in subgroup NT29 (GenBank accession no. L30130). Five isolates formed a distinct, separate cluster together with B. garinii ChY13p (GenBank accession no. AB003785) and Borrelia with a similar spacer structure from northeastern China and Japan (GenBank accession nos. EU325673 and AB015912), with isolates Ipv-6271 and Ip-6322 showing 100% homology to B. garinii ChY13p. The attribution of isolates Ipv-6272, Ipv-6290 and Ip-6319, Ipv-6271 and Ip-6322, and Ipv-6283 to a appointed genomic subgroup (Fig. 1) was confirmed by the results of sequencing of the rrs gene fragment. Within the genomic subgroups, Borrelia from Tomsk showed complete homology in the rrfA–rrlB spacer sequence, irrespective of whether they were isolated from I. pavlovskyi or I. persulcatus ticks.
Discussion
The high Borrelia infection rate in I. pavlovskyi ticks confirms the opinion that this vector is a good host for spirochetes and is involved in the epizootic process (Gorelova et al. 2001). Moreover, the data presented above provide evidence for the existence of natural foci in which taiga ticks are scarce or absent, and the role of the main arthropod host and vector of Borrelia is played by I. pavlovskyi. Larvae and nymphs of this species have a wide range of feeding hosts, mainly small mammals and birds. Unlike I. persulcatus, adult I. pavlovskyi ticks readily attack birds (Ushakova and Filippova 1968, Filippova 1971, 1977, Sapegina 1972). Therefore, birds as well as small mammals may be major reservoir hosts of Borrelia within the I. pavlovskyi range, both in the foci where this species occurs together with the taiga tick (Gorelova et al. 2001) and in the absence of the latter. On the other hand, adult I. pavlovskyi is less aggressive toward humans and attacks them much less frequently than does I. persulcatus (Kovalevskii et al. 1975). Attacks on humans by larvae and nymphs of I. pavlovskyi tick are not described. The epidemic significance of I. pavlovskyi is apparently much lower than that of the taiga tick even at a similar level of their abundance. Reliable data allowing an objective assessment of the role of I. pavlovskyi ticks in the transmission of Borrelia to humans are as yet absent.
On the whole, the results of genotyping the isolates obtained in this study show that B. garinii prevail in ixodid ticks from both natural foci (31 out of 35 isolates). This genospecies is also more frequent in other regions of Western Siberia (Fomenko et al. 2006, 2008), in contradiction to the previous opinion that B. afzelii is more widespread there (Rudakova et al. 2001).
In the Tomsk region, as in the Cisural region of Eastern Europe (Nefedova et al. 2007), B. garinii of two subgroups, 20047 and NT29, may circulate simultaneously in a natural focus, irrespective of whether the main vector is I. pavlovskyi or I. persulcatus. However, B. garinii NT29 appear to be more closely associated with I. persulcatus, as is the case with Borrelia of this genomic subgroup in the rest of their range (Postic et al. 1997, Korenberg et al. 2002). Moreover, both I. pavlovskyi and I. persulcatus ticks yielded Borrelia isolates with the nucleotide sequences of the rrfA–rrlB spacer and rrs gene fragment identical to those in B. garinii ChY13p from I. persulcatus collected in China (Li et al. 1998, Takada et al. 1998). Completely or highly homologous sequences were also found in Borrelia isolated from small mammals and I. persulcatus ticks from China, Japan, and Russia (Western Siberia) and also from Haemaphysalis flava tick nymphs collected from migrating birds in Japan (Ishiguro et al. 2000, 2005, Fomenko et al. 2006, 2008). With respect to the rrfA–rrlB spacer sequence, our isolates Ipv-6271 and Ip-6322 proved to differ from B. garinii 20047 and NT29 (no more than 89.8 and 88.7% homology, respectively), being closer to B. afzelii VS461 (93.8% homology). It may well be that all these isolates, which form a well-defined separate cluster in the dendrogram (Fig. 1), represent an individual genomic subgroup (its tentative name, ChY13p, is parenthesized in the Table 1 and Fig. 1). However, this hypothesis requires special, more detailed substantiation, which is beyond the scope of this paper.
Footnotes
Acknowledgment
This study was supported by the Russian Foundation for Basic Research, project no. 07-04-00286.
Disclosure Statement
No competing financial interests exist.