
Abstract
Plague is a bacterial (Yersinia pestis) disease that causes epizootic die-offs in black-tailed prairie dog (Cynomys ludovicianus) populations in the North American Great Plains. Through their grazing and burrowing, prairie dogs modify vegetation and landscape structure on their colonies in ways that affect other grassland species. Plague epizootics on prairie dog colonies can have indirect effects on species associated with colonies. The mountain plover (Charadrius montanus) preferentially nests on black-tailed prairie dog colonies and is thus negatively impacted by the loss of prairie dogs. We studied the effects of plague and colony spatial characteristics on the occupancy of 81 prairie dog colonies by nesting plovers in Phillips County, Montana, during a 13-year period (1995–2007). We used a robust design patch occupancy model to investigate how colony occupancy and extinction and colonization rates were affected by plague history, colony size, and colony shape. Here extinction and colonization rates refer to the probability that a colony loses/gains plovers in a subsequent nesting season, given that it had/lacked plovers in that breeding season. Colony occupancy was best explained by a model with no annual variation or plague effects. Colony extinction rates were driven by a combination of a quadratic of colony area, a 3-year plague response, and a measure of colony shape. Conversely, colonization rates were best explained by a model with a 4-year plague response. The estimated annual proportion of colonies occupied by plovers was 0.75 (95% confidence interval = 0.57–0.87). Estimated extinction probability ranged from a low of 0.07 (standard error [SE] = 0.02) in 2002 to a high of 0.25 (SE = 0.03) in 1995; colonization probability ranged from 0.24 (SE = 0.05) in 2006 to 0.35 (SE = 0.05) in 2000. Our results highlight how a bird that depends on prairie dogs for nesting habitat responds to plague history and other spatial characteristics of the colony. Ultimately, this information will broaden our understanding of the impact of plague on species other than the host.
Introduction
Patch dynamics of the prairie dog (Cynomys sp.), an important and widespread herbivore sometimes described as a keystone species (Koford 1958, Paine 1969, Miller et al. 1994, Stapp 1998, Kotliar 2000), are of interest in the North American Great Plains. Prairie dogs are colonial, semi-fossorial rodents whose intensive herbivory and burrow creation alter plant community and soil characteristics on their colonies (Whicker and Detling 1988, Baker et al. 2000). In Montana, United States, where we conducted this study, colonies are typically <40 ha in size and separated from one another by <10 km. Colonies provide a home for a wide range of other vertebrate (Lomolino and Smith 2003, Smith and Lomolino 2003) and invertebrate (Bangert and Slobodchikoff 2006) species that have adapted to this particular habitat. The mountain plover (Charadrius montanus) is one of a suite of bird species that associates with prairie dog colonies, alongside the burrowing owl (Haug et al. 1993), ferruginous hawk (Bechard and Schmutz 1995), and others. The plover is a localized breeding bird of the western North American Great Plains (Knopf and Wunder 2006) where it nests in a variety of grasslands and shrublands, especially in areas disturbed by prairie dogs (Knowles et al. 1982, Dinsmore et al. 2005). At the northern limit of its range in north-central Montana, the plover relies on active black-tailed prairie dog (Cynomys ludovicianus) colonies to survive because they provide a mix of bare ground and short vegetation suitable for reproduction (Dinsmore et al. 2003).
Prairie dogs in north-central Montana are highly susceptible to plague (Collinge et al. 2005), which is caused by the bacterium Yersinia pestis and is spread primarily by the bite of an infected flea (Barnes 1982, Gage and Kosoy 2005). Prairie dogs experience short plague epizootics that result in widespread die-offs, interspersed with periods of plague inactivity (Antolin et al. 2002). Plague epizootics are an important driver of regional prairie dog population dynamics in areas where plague occurs (Antolin et al. 2002, 2006, Hartley et al. 2009). In Montana where this study was conducted, a major epizootic affecting >80% of the colonies occurred in 1992–1995 and smaller epizootics affecting fewer colonies have occurred during subsequent years (Collinge et al. 2005). Typically, plague eliminates >95% of the prairie dogs in <1 week (Cully 1997, Cully and Williams 2001, Pauli et al. 2006) and vegetation height increases with the removal of grazing (Hartley et al. 2009), although sometimes the recovery is slower. Most plague events in north-central Montana occur during the plover breeding season (May–July; personal observation). Because the plover begins nesting in April to May, plague does not appear to affect their occupancy of a colony until the following nesting season. In subsequent years the plague event causes plovers to avoid the affected colony and shift to other unaffected colonies to nest because of alterations in vegetation height. If prairie dogs recolonize a colony, plovers may later return but only after the habitat has again become suitable. On the western Great Plains, plover nesting habitat closely tracked the area occupied by prairie dogs and nest numbers declined rapidly (within 1–2 years) on plague-affected colonies (Augustine et al. 2008). Thus, prairie dogs provide important habitat for nesting mountain plovers and plague is a primary mechanism for changing the spatial arrangement of prairie dog colonies.
Here we describe the patch dynamics of the mountain plover during a 13-year period on a large complex of plague-affected black-tailed prairie dog colonies in southern Phillips County, Montana. We modeled the occupancy dynamics of prairie dog colonies by plovers as a function of plague history, colony size, and colony shape in an attempt to better understand plague effects on plovers. We discuss our findings as they relate to understanding the indirect impacts of plague on a migratory bird during its nesting season.
Materials and Methods
Study area
We studied mountain plovers and black-tailed prairie dogs from 1995 to 2007 on a 3000-km2 area in southern Phillips County in north-central Montana (47°40′–47°55′N,107°35′–108°30′W; Fig. 1). The study area is bounded by the Missouri River to the south, the Sun Prairie and Content roads to the east, Beaver Creek to the north, and Highway 191 to the west. Approximately 2250 km2 of the study area is in public ownership with the Bureau of Land Management (BLM, Malta Field Office) and the U.S. Fish and Wildlife Service (USFWS, Charles M. Russell National Wildlife Refuge). This area was a mixed-grass prairie typified by flat-topped ridges dissected by shallow coulees and sagebrush flats (Knowles et al. 1982, Olson and Edge 1985, Dinsmore et al. 2002). Primary plant species on active black-tailed prairie dog colonies included fringed sagewort (Artemisia frigida), plains prickly pear (Opuntia polycantha), blue grama (Bouteloua gracilis), needle-and-thread grass (Stipa comata), and Sandberg bluegrass (Poa secunda). Mean elevation was approximately 930 m.
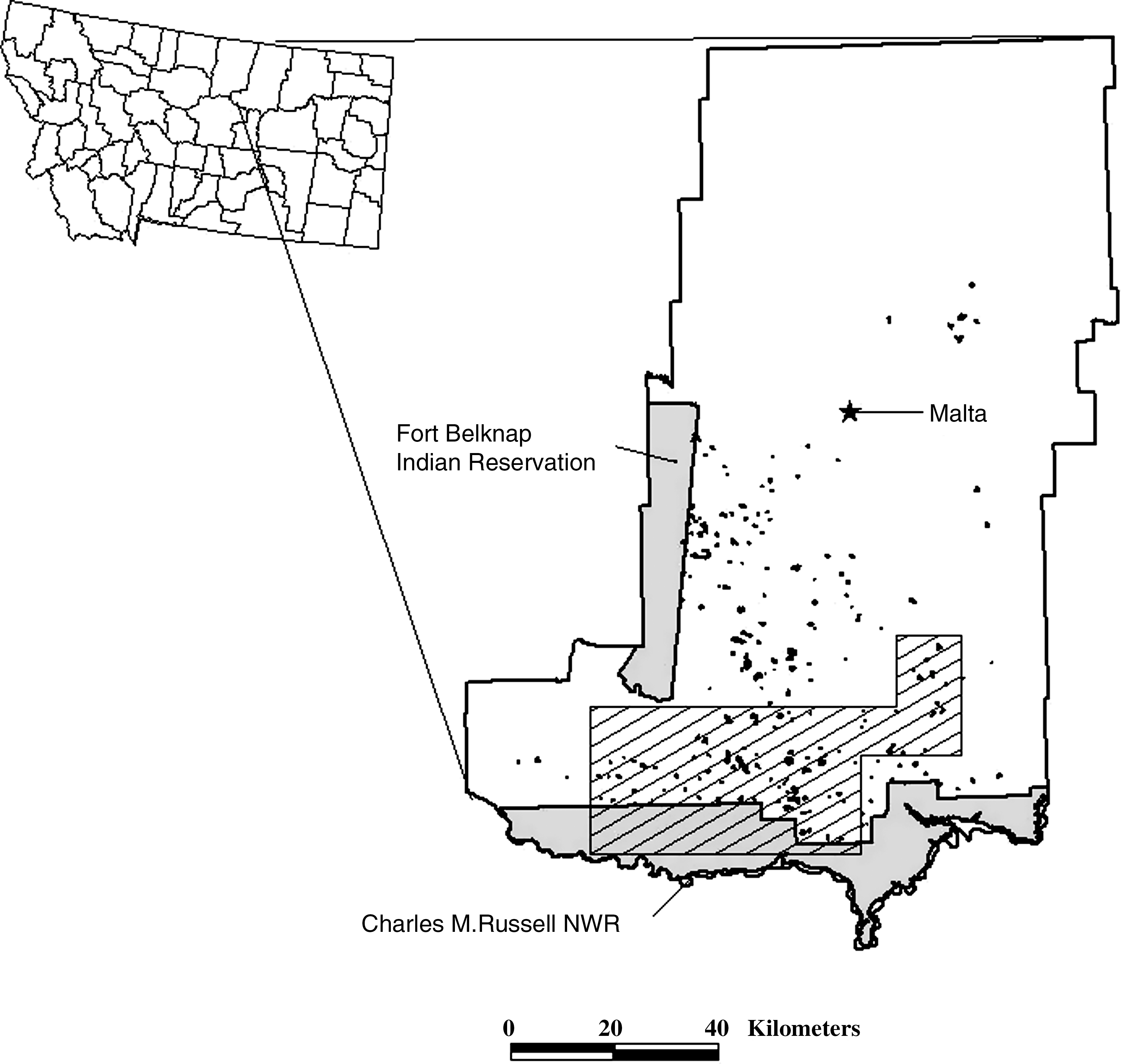
Map of Phillips County, Montana, showing the 2000 distribution of black-tailed prairie dog colonies (in black), Charles M. Russell National Wildlife Refuge, and Fort Belknap Indian Reservation. The stippled region represents the study area.
Within this region we studied mountain plovers exclusively on black-tailed prairie dog colonies. We defined a colony as a specific location on the landscape that was occupied by prairie dogs for ≥ 1 year since 1995. Plovers arrive in early to mid-April and quickly set up a nesting territory on a colony. Our color banding data indicate that individual adult plovers remain faithful to a prairie dog colony through the season of a plague epizootic and then move to another colony in a subsequent year (Skrade 2008). This delayed movement away from a plague-affected colony is likely because plague occurs after most birds have initiated nests, at a time when they are too invested in that year's nesting attempt to abandon. We continued to monitor inactive prairie dog colonies in subsequent years to check for prairie dog recolonization and because plovers occasionally used them when habitat conditions were favorable (e.g., low vegetation and bare ground) even in the absence of grazing. New prairie dog colonies were formed nearly every year and we attempted to survey them soon after they were established.
Surveys for plovers
All active (and some inactive) prairie dog colonies were searched for the presence of at least one mountain plover more than three times each year by vehicle, once or more in each of three sampling periods (May 20–June 10, June 11–30, and July 1–20) spanning the primary plover nesting season. Previous work has shown that plovers seldom nest on inactive colonies (Dinsmore et al. 2003, 2005), so our surveys emphasized active colonies. For these analyses, we used only the first visit within each sampling period as an indication of occupancy for that period. During a survey, the observer traversed the entire colony by vehicle, often stopping periodically to scan for adult plovers with binoculars. We did not standardize survey time to colony area because we also looked for nests (part of a separate study), although large colonies took longer to search than small colonies. Because we were only interested in presence–absence (occupancy) data, the number of plovers detected on a survey is unimportant. The class of occupancy models we used in our analyses (see below) assumes demographic closure (e.g., no births, deaths, emigration, or immigration of plovers) during the nesting season. Surveys were only for adult plovers, so births were not an issue. Further, our previous work at this site has shown high survival of adult plovers during the nesting season and strong fidelity to nesting colonies until dispersal begins in late July (Dinsmore and Knopf 2005, Dinsmore 2008).
Prairie dog colony characteristics
We relied solely on observational data to document plague occurrence on individual colonies (see Collinge et al. 2005). For this study we defined a plague event as a >90% reduction in colony area in a single year. This definition may miss smaller epizootics that did not affect an entire colony, although previous work has shown that plague typically causes this level of response in black-tailed prairie dogs (Cully 1997, Cully and Williams 2001). Consequently, this definition should produce conservative study results of the influence of plague on mountain plovers because our models only include obvious epizootics. We obtained spatial data for all active prairie dog colonies from BLM and USFWS staff and our own efforts. From these sources, we obtained two types of data: (1) the area (in hectares) of each active prairie dog colony and (2) Geographic Information System (GIS) coverages showing colony boundaries. Below we discuss how we collected and subsequently used each source of data.
Annual measures of prairie dog colony area were obtained by using a portable global positioning unit to delineate the boundaries of each active colony. A complete census of all known active colonies was done in 1998, 2000, 2002, 2004, and 2007. In other years we obtained a measure of the area of each colony in one of four ways: (1) from an incomplete census of colonies in the study area, (2) by extrapolating area based on an estimated rate of change in a sample of one-third of all known active colonies, (3) by using the midpoint of colony area in adjacent years, or (4) from other ancillary sources (e.g., a plague event that eliminated a colony). We set the colony area equal to the mean of all other colonies that year in instances where we were unwilling to estimate colony area using one of the four approaches above. This effectively removes a colony from the calculation of area effects for that year.
Data from prairie dog colony mapping efforts were converted to shapefiles (only for colonies that were actually mapped in the field). We used ArcGIS to calculate the area of each mapped colony. We used FRAGSTATS (Version 3.3; McGarigal and Marks 1995) to calculate colony spatial features of interest with a 10-m grid cell size. Because we lacked shapefiles for many years, we calculated spatial characteristics only for colonies that were actually mapped in the field.
Occupancy modeling
We used the robust design patch occupancy model (MacKenzie et al. 2002) in Program MARK (White and Burnham 1999) to investigate plague impacts to mountain plovers. This model generates annual estimates of the proportion of prairie dog colonies occupied by plovers (Ψ), a colonization rate of prairie dog colonies by plovers (γ), an extinction rate of plovers using colonies (ɛ), and the probability of detecting plovers on prairie dog colonies (p). Both ɛ and γ are conditional probabilities. Thus, γ is the probability that a colony not occupied by plovers in year t will be occupied by plovers in year t + 1 and ɛ is the probability that a colony occupied by plovers in year t will become unoccupied in year t + 1. Collectively, these parameters can be used to describe the complex patch dynamics facing a mountain plover in a plague-affected region.
A priori predictions
We developed a list of biological factors to explain how plague history and the resultant changes to colony size and shape influenced the occupancy of black-tailed prairie dog colonies by nesting mountain plovers. Below, we identify three primary sources of variation in occupancy and state our hypothesis regarding each of them.
(1) Plague history. The first known epizootic in Phillips County was in 1992 and several smaller epizootics have occurred more recently (Collinge et al. 2005). We believed that a colony with a history of plague was less likely to be colonized by plovers and exhibited a greater probability of extinction by plovers than a colony not affected by plague. This could occur because plovers remember colonies where the vegetation is taller and less suitable for nesting, colony area declined (see below), or because of unknown plague effects that somehow diminish a colony's suitability for plovers. Although we did not directly measure vegetation height or composition after epizootics, previous research has shown that vegetation height does increase on colonies after a plague epizootic in prairie dogs (Hartley et al. 2009). We surmised that as the time (in years) since a plague event increased, annual extinction rates of plovers should decrease while colonization rates of plovers should increase if a colony has been recolonized by prairie dogs. We further believed that a plague effect might be either acute (immediate loss of plovers after an epizootic) or chronic (immediate loss of plovers followed by a slow recolonization). To model this, we included four sets of covariates for each colony. The first set accounted for the presence or absence of plague on a colony in each of the 13 study years (1995–2007), for example, 0000100000000 for a colony that was decimated by plague in 1999. The remaining three sets of covariates accounted for the time since a plague epizootic, for example, 0000432100000 for a colony that was decimated by plague in 1999. Note that the nonzero numbers in these last three sets of covariates are simply dummy variables and do not force any directionality in the plague effect. We modeled these chronic effects by looking at 2-, 3-, and 4-year time lags since plague and hypothesized that both extinction (ɛ) and colonization (γ) rates were most likely to be explained by long-term chronic effects of plague. Plague history was intended to account for the effects of plague that were not directly accounted for in the annual measures of colony size (see below).
(2) Prairie dog colony size. The size of a prairie dog colony is thought to be an important factor in predicting occupancy by mountain plovers. Early work in Montana suggested that plovers selected colonies between 6 and 50 ha in size (Olson-Edge and Edge 1987). Colony size is reduced after a plague event, although recolonization can be rapid. We believed that larger colonies were more likely to be colonized and less likely to be vacated by plovers. We modeled this as a linear and quadratic effect on both extinction (ɛ) and colonization (γ) rates to account for the possible preference for mid-sized colonies identified in the earlier study.
(3) Colony edge effect. Our personal observations of nesting mountain plovers suggested that there might be a slight tendency for plovers to nest along a colony edge, rather than in the center of a colony. While we had no evidence that this colony characteristic was a result of plague, we could envision an increase in colony edge as a result of prairie dog recolonization after an epizootic. We hypothesized that occupancy might be positively related to increased edge within a colony. To model colony edge effects we computed two simple shape indices for each colony. We calculated a simple patch shape index (PSI) and the perimeter to area ratio for each colony. Both metrics resulted in year-specific covariates for each colony, except 4 years (1997, 2003, 2005, and 2006) when we had little or no spatial data. The shape index takes on values ≥1; a value of 1 indicates a patch that is maximally compact (e.g., a square) and patch irregularity increases with higher values (McGarigal and Marks 1995).
The patch occupancy model does not currently have a goodness-of-fit test, and we were unable to rigorously test for a lack of independence in the data (overdispersion).
Model selection and parameter estimation
We evaluated competing models to explain plague impacts to plover occupancy of prairie dog colonies using the methodology of Burnham and Anderson (2002). We ranked the set of candidate models using Akaike's Information System (AIC), corrected for small sample bias (AICc; Akaike 1973, Burnham and Anderson 2002). We did not adjust for extrabinomial variation in these data; there is currently no method for estimating the extrabinomial variation in the patch occupancy model. We ranked all models in the candidate model set relative to the model with the lowest AICc value. Generally, models with ΔAICc values <2 have strong support, whereas those with ΔAICc values >10 have little support (Burnham and Anderson 2002). Normalized Akaike weights (wi ) were also computed for each model, provided another means of directly evaluating the strength of evidence for each model, and were useful for computing parameter estimates that reflected model selection uncertainty (Burnham and Anderson 2002). Instead of presenting parameter estimates from a single “best” model, we model averaged (Burnham and Anderson 2002) parameter estimates across the entire model set. Finally, we used the logistic regression equation from the best model to make predictions about the effects of plague, colony area, and colony shape metrics on extinction and colonization rates.
Results
We examined the influence of plague history and colony spatial characteristics on occupancy of black-tailed prairie dog colonies by mountain plovers using a total of 81 prairie dog colonies in the 13-year study period (Fig. 2). Colony size ranged from 0 to 246 ha; the mean across all colonies and all years was 27.88 ha (standard deviation = 31.99).
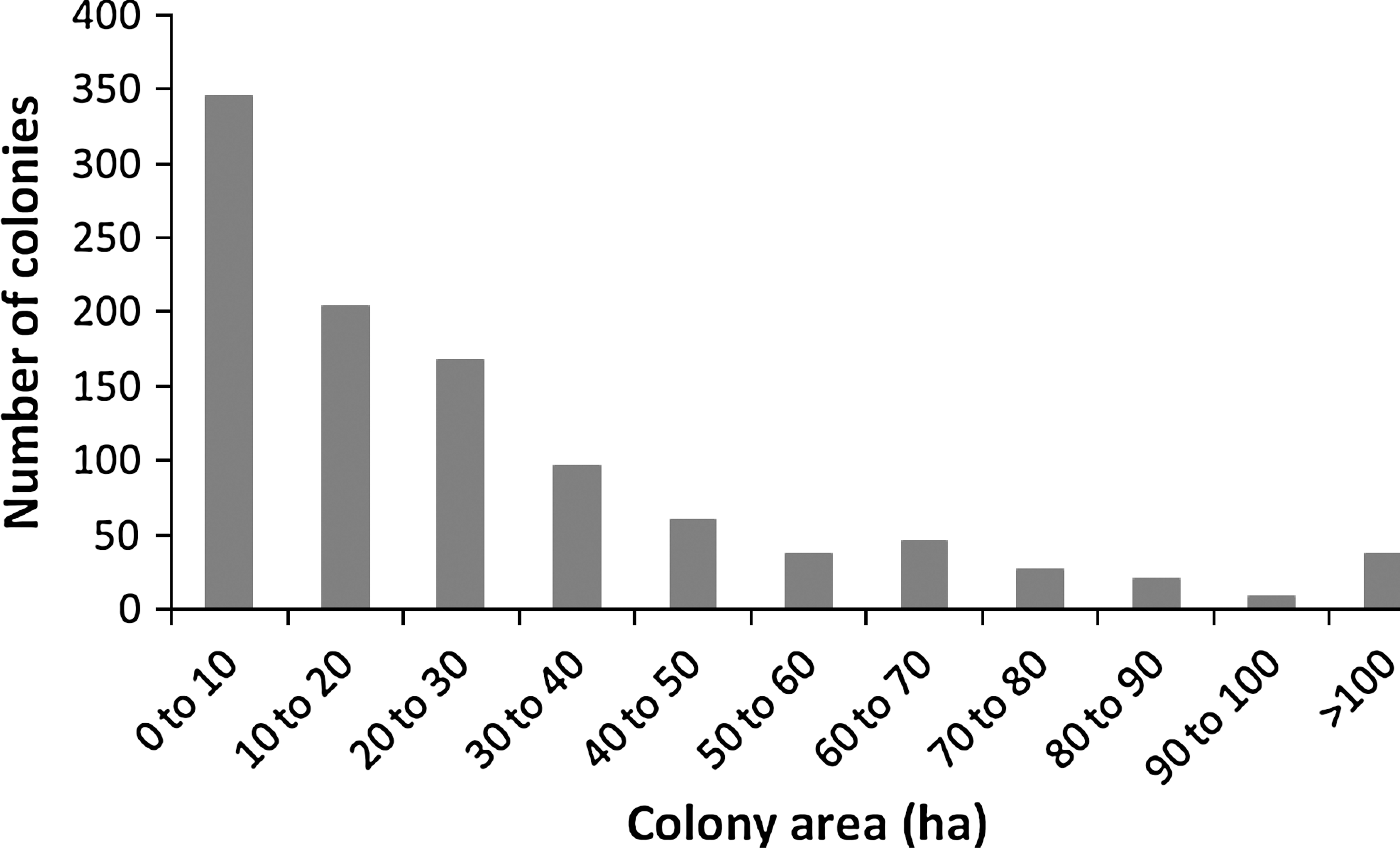
Distribution of black-tailed prairie dog colonies by size (hectares) in southern Phillips County, Montana, 1995–2007. Note that each of the 81 colonies visited during this study could contribute as many as 13 values (1 per year) to this distribution. The first column includes 63 colonies that were inactive because of a plague epizootic (area of zero).
Modeling results
On the basis of our analyses, plague history and colony size were the most important variables explaining occupancy of prairie dog colonies by plovers (Table 1). There was strong evidence that resighting probability varied by survey period within year; models with the first two periods equal were not well supported. In the best model, ψ was constant across years; ɛ contained the effects of quadratic area, a 3-year plague response, and PSI; and γ contained a 4-year plague response (Table 1). All competitive (ΔAICc <2) models contained the same quadratic area and 3-year plague effects on ɛ. From the best model, ɛ decreased as a function of time since plague (
All models in this table had ψ constant across years and capture probability differing by session within year, so only effects on ɛ and γ are listed in the model column. Only models with a ΔAICc <5 are shown. Model effects included a quadratic effect of area (Area2), 1- to 4-year plague effects (1PL, 2PL, 3PL, or 4PL), a patch shape index (PSI), perimeter-to-area ratio (PAR), or no effect (.).
The best model had an AICc value of 2252.15.
Parameter estimates
The detectability of plovers declined across the three sampling periods within years, dropping from 0.85 (standard error = 0.02) in the first period to 0.64 (standard error = 0.02) in the third period. The estimated proportion of colonies occupied by plovers was 0.75 (95% CI = 0.57–0.87) annually. Model-averaged estimates of ɛ and γ diverged early in the study period and then converged toward the end, presumably in response to major plague events at the beginning and end of the study (Fig. 3). Colonization probability increased through 2001, then declined; extinction probability declined through 2003 and then increased (Fig. 3). Finally, we used the logistic regression equation from the best model to predict time-since-plague responses in ɛ for prairie dog colonies of differing sizes (Fig. 4). We also used this model to predict the response in g, which increased from 0.05 one year after plague to 0.31 four years after plague.
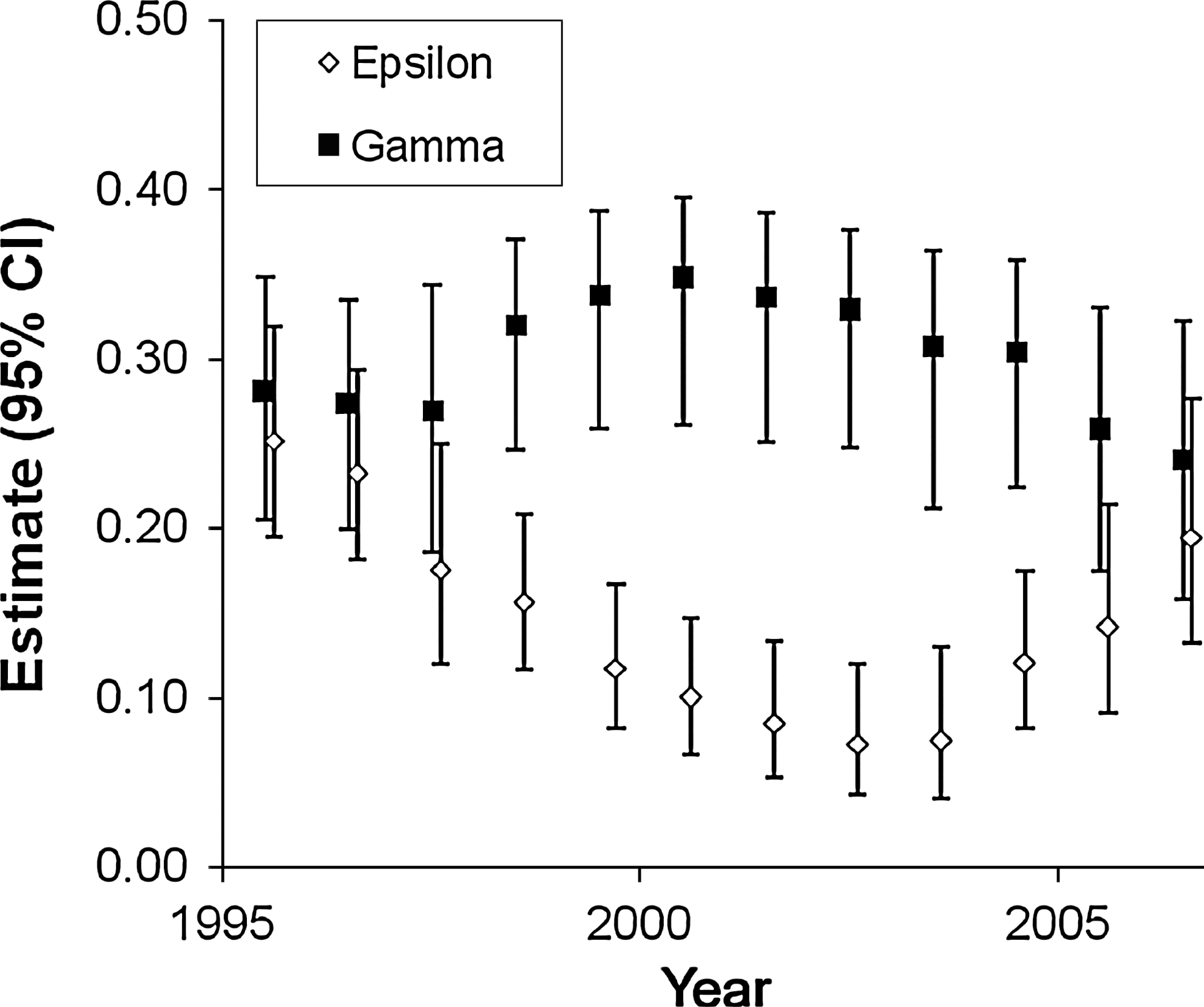
Model-averaged estimates (95% confidence interval) of extinction (epsilon) and colonization (gamma) probabilities for mountain plovers occupying black-tailed prairie dog colonies in southern Phillips County, Montana, 1995–2007.
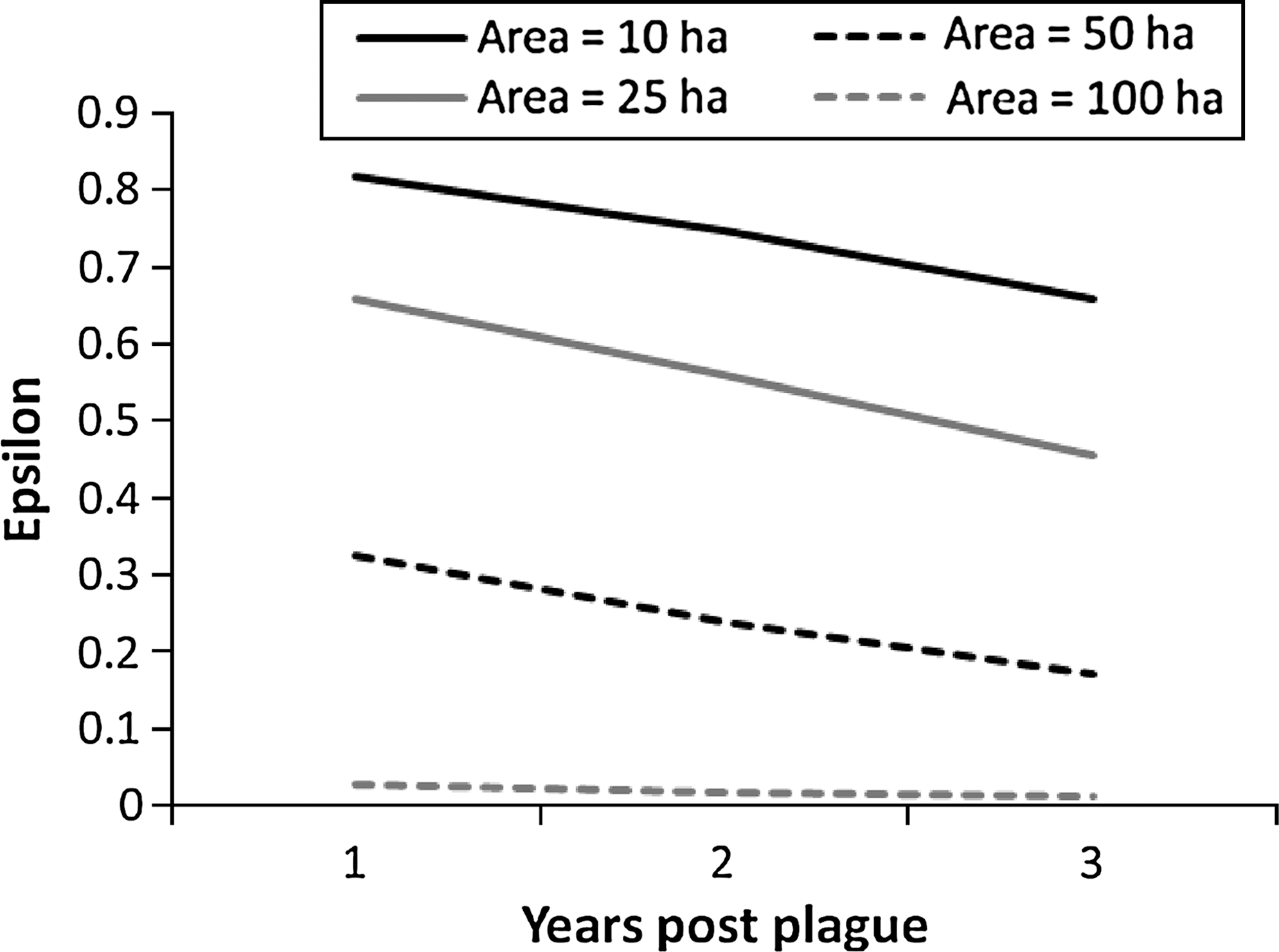
Predicted time-since-plague responses in extinction probability (ɛ) of mountain plovers using prairie dog colonies in southern Phillips County, Montana, 1995–2007. Estimates were generated using the logistic regression equation for the best model and prairie dog colonies ranging from 10 to 100 ha.
Discussion
Our study documents the indirect impacts of plague in prairie dogs on mountain plovers by looking at patterns of colonization and extinction of plovers as they relate to occupancy of black-tailed prairie dogs in Montana, United States. The annual proportion of colonies occupied by plovers was constant during the study period, but patterns of plover colonization and extinction for colonies were strongly affected by plague history and colony size.
We made important assumptions regarding data collection and analyses that may have affected our findings. As noted earlier, we relied on observational data to identify plague events. We also note that prairie dog colony area represented only an index of plover habitat. A measure of the area of a prairie dog colony does not account for factors such as prairie dog density, vegetative differences, or differences in topography such as slope and elevation, all of which may be important to plovers (Knopf 1996). Our sample of 81 colonies was large enough to address our primary research questions, although our results may have been affected because we had few large colonies compared to those that were small to mid-sized. While plague is clearly an important mechanism driving occupancy of prairie dog colonies by plovers, other agents like recreational prairie dog shooting (Vosburgh and Irby 1998) could also contribute to this pattern. Finally, we reiterate that our study includes only presence–absence data; a more detailed analysis that incorporates plover abundance or density might produce slightly different inferences about the effects of plague on mountain plovers.
Indirect impacts of plague to plovers
Our study documents the indirect impacts of plague in prairie dogs on the mountain plover during its nesting season on the northern Great Plains. The proportion of colonies occupied by plovers did not vary between years, although the specific subset of colonies occupied differed between years as a result of plague. Plague is not known to have direct, lethal impacts to plovers, but its indirect effects alter both the amount and distribution of plover nesting habitat in this region. Each of these affects plover persistence, which is a concern because of its heightened conservation status (Dinsmore 2000, U.S. Department of the Interior 2002, 2003).
Plague is lethal to prairie dogs and often eradicates an entire colony (Menkens and Anderson 1991, Cully 1997, Cully and Williams 2001). With the disappearance of prairie dogs after an epizootic come multiple changes in the habitat, on the former colony vegetation height increases with the removal of grazing (Weltzin et al. 1997), the insect community is altered (Bangert and Slobodchikoff 2006), and other changes may occur. Each of these has important implications for the mountain plover. The plover prefers to nest in short-grass habitats with a substantial bare ground component. This condition is met on an active prairie dog colony, but on an inactive colony the vegetation can quickly flourish and bare ground disappears making the colony less attractive to a plover. The plover's diet is diverse, and during a single breeding season study in Colorado it was comprised of items from 90 taxa, predominantly ground-dwelling beetles (Order Coleoptera) and grasshoppers and crickets (Order Orthoptera) with darkling beetles (Eleodes sp.) comprising 22% of the diet (Baldwin 1971). A later study (Knopf 1998) concluded that the diet was probably more flexible than lone study and incidental observations hinted. It seems likely that Coleoptera in particular may be less abundant after a plague event because their steady food supply (dung) is missing; refugia (burrows) also disappear, but more slowly over a period of several years. In response to vegetative and other changes, the insect community on an extinct prairie dog colony is characterized by greater arthropod abundance relative to an active colony (Bangert and Slobodchikoff 2006). The timing of changes in the insect community is largely unknown, and it is possible that such changes occur gradually in the years following a plague epizootic. The often profound habitat changes and shift in the food supply cause plovers to quickly abandon a plague-affected prairie dog colony (Dinsmore et al. 2005). As suggested by Bangert and Slobodchikoff (2006), insect community differences between active and inactive prairie dog colonies, and between colonies and adjacent grasslands, may be a bottom-up community process with important implications to insectivorous vertebrates such as the mountain plover.
Plague impacts the spatial arrangement of colonies at a larger scale by its removal of individual prairie dog colonies. Since the introduction of plague in the early 1900s (Barnes 1993), black-tailed prairie dog colonies have begun to act as meta-populations (Roach et al. 2001) subject to local extinctions and recolonization. Recolonization may result from a combination of survivors that develop antibodies to Y. pestis (Pauli et al. 2006) and immigrants from surrounding colonies (Cully and Williams 2001, Johnson and Collinge 2004, Antolin et al. 2006), although the latter is probably more important. In northeastern Colorado, patterns of extinction due to plague have been positively linked to cool and wet summers when the El Nino Southern Oscillation is active (Stapp et al. 2004). Recolonization is rapid in drier and warmer years and continues until conditions are suitable for another plague epizootic. Much has been written about dispersal in prairie dogs (Hoogland 1995), including the possibility that infected individuals may be responsible for the spread of the disease to unaffected colonies. As a result of the mechanisms for prairie dog dispersal, isolated colonies exhibit longer persistence and a lowered risk to plague (Lomolino et al. 2003, Stapp et al. 2004) than spatially aggregated colonies.
A recent study using many of these same prairie dog colonies (Augustine et al. 2008) found that declines in prairie dog colony area as a result of plague corresponded to rapid (<2 years) declines in mountain plover nesting activity. This is presumably a response to vegetative (Hartley et al. 2009) and possibly other changes that negatively impact the plover. In plague-affected prairie dog complexes in Montana and Colorado, Augustine et al. (2008) concluded that plover nesting habitat closely tracked the area occupied by prairie dogs, although not all prairie dog colonies provide plover nesting habitat because of important microhabitat features such as vegetation structure (Olson and Edge 1985), amount of bare ground (Knopf and Wunder 2006), and slope (Graul 1975).
In this study we further demonstrated negative effects of plague to the plover by examining how they depart and later recolonize colonies affected by plague. Plovers quickly vacate plague-affected colonies (Dinsmore et al. 2005); the predicted rate of extinction increases more than 10-fold as colony area decreases from 100 to 10 ha. We attribute this rapid extinction rate on small colonies to the plover's preference for larger colonies (Olson-Edge and Edge 1987) and the ease with which it can search for and locate other more suitable colonies nearby. Conversely, plovers colonize plague-affected colonies more slowly than they vacate them (a 4- vs. 3-year lag), and this parameter was not affected by colony area. We attribute the lack of an area effect to the conditional nature of this parameter—very few large colonies are unoccupied and thus available to be colonized. Predicted colonization rates increased more than sixfold during a 4-year postplague time period. Plovers appear unlikely to colonize those colonies that are recently impacted by plague, but that probability rapidly increases with time since plague. Colony shape was not a strong predictor of either rate, except for a weak suggestion that plovers were more likely to vacate the most regularly shaped colonies, which have a lower edge to area ratio. The extinction and colonization rates of plovers we report herein, coupled with an understanding of how they are affected by plague history and colony attributes, should prove useful for better understanding the effects of plague on a nonhost species.
Implications for plover conservation
The mountain plover was recently denied listing under the U.S. Endangered Species Act (U.S. Department of the Interior 2003), although they are still of conservation concern throughout their breeding range, including Montana (Montana Natural Heritage Program 2008). Historically, the plover was an associate of the prairie dog ecosystem throughout its range, although the current association with prairie dogs is most evident in the northern portions of its range (e.g., Montana; Knowles et al. 1982, Dinsmore et al. 2005) and in Colorado (Dreitz et al. 2005). As suggested by Augustine et al. (2008), plover persistence in many areas may be closely linked to the persistence of prairie dogs. Plague, along with recreational shooting and poisoning, has been identified as a key threat to prairie dog persistence (Miller et al. 1990). We suspect the plover has the ability to disperse in response to changing environmental conditions (e.g., rainfall), although the introduction of plague may have altered selective pressures and altered their site fidelity and dispersal patterns. There have been multiple approaches to conserving continental prairie dog populations, including strategies to combat the spread of plague. One approach has been to treat prairie dog colonies with an insecticide to kill fleas (Karhu and Anderson 2000). Such treatments are effective for 1 year, expensive, and have indirect effects on the insect community. A study on black-tailed prairie dog colonies in Colorado found significant declines in Coleoptera as a result of treatments (Karhu and Anderson 2000); this Order is an important food item for the plover (Knopf 1998). Stopping the long-term spread of plague is a challenge that is hampered by uncertainty in vectors for transmission between colonies. Many possible vectors have been identified, but none has been documented in field studies. Plague may persist for >1 year in an affected colony, although recent field studies have both confirmed (Hanson et al. 2007) and refuted (Holmes et al. 2006, Salkeld and Stapp 2008) this claim. Uncertainty in mechanisms for plague transmission, especially as they relate to flea ecology, and the long-term ability of plague to persist in affected colonies are key unanswered research questions.
Throughout the northern part of its breeding range, the mountain plover is closely tied to active prairie dog colonies. A recent study of the plover's inter-year site fidelity and responses to plague (Skrade 2008) suggests that an understanding of prairie dog colony dynamics is crucial to future management of the plover in Montana. The current focus on prairie dog conservation (Miller et al. 1994, 2007), including management of plague, may result in recovery of prairie dogs in other parts of the plover's breeding range. Detailed information on plover responses to plague may eventually prove useful to managers interested in developing multispecies approaches to the conservation of the prairie dog ecosystem.
Footnotes
Acknowledgments
We are grateful to the U.S. BLM (Malta Field Office, Montana), Fort Carson Military Reservation (U.S. Department of Defense, Colorado), the USFWS (Colorado Fish and Wildlife Management Assistance Office), the Biological Resources Division of the U.S. Geological Survey (Fort Collins Science Center, Colorado), the Denver Zoological Foundation, Montana Fish, Wildlife, and Parks, and the Fort Belknap Tribal Community for financial support. The staff at Charles M. Russell National Wildlife Refuge supplied additional logistical support. Grensten, JJ, Knopf, FL, and Rosenlund, BD, assisted with multiple aspects of the study. Brees, A, Ely, DC, Hanks, T, Jorgensen, JG, Matchett, R, McQueary, C, Schmitz, R, Skrade, PDB, Veseth, D, Veseth, F, Vosburgh, T, and Wilcox, CT, provided field assistance. We thank Matovitch, B, Robinson, D, and Robinson J, for allowing us access to their lands.
Disclosure Statement
No competing financial interests exist.