
Abstract
Plague causes periodic epizootics that decimate populations of prairie dogs (PDs) (Cynomys), but the means by which the causative bacterium (Yersinia pestis) persists between epizootics are poorly understood. Plague epizootics in PDs might arise as the result of introductions of Y. pestis from sources outside PD colonies. However, it remains possible that plague persists in PDs during interepizootic periods and is transmitted at low rates among highly susceptible individuals within and between their colonies. If this is true, application of vector control to reduce flea numbers might reduce mortality among PDs. To test whether vector control enhances PD survival in the absence of obvious plague epizootics, we reduced the numbers of fleas (vectors for Y. pestis) 96–98% (1 month posttreatment) on 15 areas involving three species of PDs (Cynomys leucurus, Cynomys parvidens in Utah, and Cynomys ludovicianus in Montana) during 2000–2004 using deltamethrin dust delivered into burrows as a pulicide. Even during years without epizootic plague, PD survival rates at dusted sites were 31–45% higher for adults and 2–34% higher for juveniles compared to survival rates at nondusted sites. Y. pestis was cultured from 49 of the 851 flea pools tested (6882 total fleas) and antibodies against Y. pestis were identified in serum samples from 40 of 2631 PDs. Although other explanations are possible, including transmission of other potentially fatal pathogens by fleas, ticks, or other ectoparasites, our results suggest that plague might be maintained indefinitely in PD populations in the absence of free epizootics and widespread mortality among these animals. If PDs and their fleas support enzootic cycles of plague transmission, there would be important implications for the conservation of these animals and other species.
Introduction
This study was motivated by our concern for the general decline in prairie dogs (PDs) (Cynomys), the precarious status of associated species (including federally-endangered black-footed ferrets, Mustela nigripes), and federally threatened Utah PDs (UPD) (Cynomys parvidens). Our concern about plague, however, extends beyond these particular species to other species of mammals with shrinking populations in western North America (Biggins and Kosoy 2001a, 2001b); even large-scale population reductions of such species might escape notice under cursory monitoring.
All species of PDs are highly susceptible to plague (Ecke and Johnson 1950, Ubico et al. 1988, Barnes 1993). Most discussions of plague in PD populations focus on the catastrophic effects of epizootics. A prevalent conceptual model of plague in PD communities involves periodic invasion of colonies by Y. pestis organisms that come from other sources (other areas and/or host species) when the densities of PDs and presumably other rodents reach appropriate levels to sustain rapid transmission, resulting in nearly complete population collapses and subsequent local disappearance of the bacterium due to lack of hosts (Cully 1993, Fitzgerald 1993). It is difficult to imagine that PDs could be responsible for long-term maintenance of Y. pestis when epizootics commonly remove >90% of the animals. However, we observed persistence of plague for 23 years (unpublished data) in a complex of PD colonies occupying a 200 km2 area near Meeteetse, Wyoming (Forrest et al. 1985). During that period, many individual colonies within the complex suffered local population collapses with subsequent recoveries. On a larger scale, the coincidence between the emerging range of Y. pestis in North America and collective ranges of ground dwelling scirurids that form relatively dense populations (Wayson 1947), as well as failure of plague to become established in the eastern United States, continue to be topics of discussion.
Although epizootics of plague that cause high mortality of PDs over wide areas are a commonly noted phenomenon, plague may be maintained between epizootics by continued slow rates of transmission within the PD community. Other postulated mechanisms (not mutually exclusive) for maintenance of Y. pestis (Gage and Kosoy 2006) include survival of the bacterium in the soil, persistence of the bacterium in fleas, and maintenance of the bacterium in reservoir species of mammals in which Y. pestis is relatively nonlethal and causes chronic infections. Although Y. pestis has been identified as the probable cause of many population crashes of PDs, little is known about the ecology of plague in PD communities, particularly the interepizootic maintenance of Y. pestis populations and its potential effect on PDs and associates as an enzootic disease.
We use the term enzootic to describe disease occurrence at less than epizootic levels. This definition requires an effect of disease rather than simply the presence of Y. pestis within the community of hosts. It does not presume persistence or long-term stability of host populations and does not preclude involvement of multiple host species. The term epizootic itself is not defined with explicit limits, but usually refers (e.g., Cully 1993, Lincoln et al. 1998) to an epidemic in nonhuman animals affecting a high proportion of the population over a wide area. Plague epizootics typically cause widespread mortality among highly susceptible rodent species and occasionally nonrodents, including various lagormorphs and certain carnivores, most notably black-footed ferrets and felids. If plague is being maintained at enzootic levels in PD populations, it would be expected to cause chronic mortality rates in these animals. If this is indeed the case and most cases of plague among PDs are acquired via the bites of infectious fleas (Gage and Kosoy 2005, Eisen et al. 2006), it would seem reasonable to propose that controlling the flea vectors of plague should result in decreased plague transmission (Seery et al. 2003, Hoogland et al. 2004) and, therefore, increased survival rates of PDs even in the absence of catastrophic epizootics of plague. We evaluated this possibility by controlling the fleas in 15 areas considered to be active plague foci. Recognizing that other explanations could account for correlations we might detect, we also attempted to collect additional data on presence of plague and causes of mortality of PDs.
Materials and Methods
We conducted this study on white-tailed PDs (WTPD) (Cynomys Leucurus), UPDs, and black-tailed PDs (BTPD) (Cynomys Ludovicianus). Study areas (Fig. 1) were selected within regions where plague had been detected previously, but where epizootic plague had not recently occurred. An exception was East Creek, a UPD colony that had declined before our study began. We augmented the low UPD population at East Creek by releasing 94 UPDs there in 2001. This study was conducted during 2000–2005, but durations varied among the 15 study areas (Fig. 2). Experimental sites were up to 50 ha each, encompassing entire PD colonies or portions thereof. We selected two sites (dusted and nondusted) on each area that had generally similar physiographic and biotic attributes such as slope, aspect, burrow density, quantity and type of vegetative cover, and initial PD density. Matched sites were in close proximity to each other, either by splitting colonies into two parts or by using similar colonies that were separated by <11 km.
General locations of clusters of study areas in the western United States where flea control experiments were conducted on WTPDs (Cynomys leucurus), UPDs (Cynomys parvidens), and BTPDs (Cynomys ludovicianus). WTPD, white-tailed prairie dog; UPD, Utah prairie dog; BTPD, black-tailed prairie dog.

Experimental design, extent of study (horizontal bars), timing of capture–recapture trapping (vertical bars), and notes on status of prairie dog (PD) colonies involved in flea control experiment. Flea control was on separate dusted and nondusted colonies (separate) or on single colonies with both treatments (split). Study sites were in eastern Utah (WTPD), southern Utah (UPD), and north-central Montana (BTPD). The study was restricted at some areas by funding shortfalls (A), collapses of PD populations where plague was confirmed in carcasses or fleas (B), population collapses where there was no sampling for plague (C), and high levels of PD mortality due to recreational shooting (D).
We attempted to reduce the incidence of plague at treated sites by reducing the populations of fleas that transmit the disease. We controlled fleas by infusing a powdered 0.05% deltamethrin formulation (DeltaDust®, Bayer Environmental Science, Research Triangle Park, NC) into burrows during 14 July–24 October of each year with pressurized applicators (Beard et al. 1992, Seery et al. 2003), at a rate of about 4 g per burrow.
We captured PDs with Tomahawk traps set during daylight hours using an arrangement that spatially allocated traps based on PD densities within study sites. Trapping was discontinued when temperatures exceeded ∼35°C. Most trapping was accomplished during June to August (median date = 11 July) with an average annual effort of 2188 trap-hours per site. We recorded sex and age of trapped individuals, and ear-tagged them for individual identification using a #1 monel fingerling fish tag in each ear (Fagerstone and Biggins 1986). If flea and blood samples were to be collected, we anesthetized PDs in an induction chamber with isoflurane. The induction chamber consisted of a 4.25-L glass or clear plastic jar, with a wad of cotton fixed to the inside of the lid. The cotton was moistened with 1.5–2.0 mL of isoflurane immediately before placing the subject in the jar. We removed the animal after all motor activity ceased (30–60 s). We collected blood from PDs on a Nobuto paper strip via toe nail clipping and spent about 30 s combing fleas from each animal captured (Karhu 1999). On several occasions, we also collected fleas from burrows using a flannel cloth attached to a plumber's cable (Seery et al. 2003). We froze fleas or held them <2 weeks in a normal saline solution containing a surfactant (Tween 80). Blood and fleas were tested for Y. pestis at the U.S. Centers for Disease Control and Prevention (Fort Collins, CO). Blood samples were tested via passive hemagglutination assay. Fleas from individual hosts were pooled by species and sex, and emulsions were tested by mouse inoculation and subsequent isolation. Direct fluorescent antibody examinations were done on tissues from a few PDs found dead. Procedures used in this study were approved by the Institutional Animal Care and Use Committee at the Fort Collins Science Center (U.S. Geological Survey).
Interruptions in trapping, caused by high levels of PD shooting in Montana, eruptions of epizootic plague near the end of the study period, and funding shortfalls (Fig. 2) prevented us from having the intended set of five trapping occasions at all sites. We assessed survival for intervals that were 1 year in length (the horizontal lines of Fig. 2), creating a capture history matrix that had year 5 as the final year for each site, regardless of calendar year. Thus, time effects were relevant to age changes in animals but not to changes in calendar years. Because dusted and nondusted colonies were paired using ecological similarities and because we sought to avoid an unbalanced experimental design, deletion of one colony also caused deletion of its pair during that year.
During the first year of the study at UPD and WTPD sites, we assessed flea control about 1 month after dust was applied. For all years except the final year, we assessed flea control efficacy on all sites about 10 months after dust application. We used Mann–Whitney tests to compare overall flea abundance on PDs from dusted and nondusted sites.
We conducted visual counts of PDs for 3–5 days (Fagerstone and Biggins 1986) at each study site, using maximum counts as indices to PD populations present. To avoid the effect of epizootics when assessing survival rates, we did not include data from sites if either the dusted or paired nondusted site had visual counts of PDs that declined >90% compared to the previous year. Low counts (often zero or nearly zero) were sometimes associated with evidence of plague (Table 1), supporting the assumption of epizootic plague as the cause of the population collapse. Visual counts typically detect about half the PDs present during a counting session (Biggins et al. 2006); lack of PDs during a visual count does not necessarily mean that none was present, but we considered this a population collapse, with epizootic plague assumed to be the cause.
Number of samples testing positive for plague, followed by total numbers (in parentheses) of flea pools or PDs tested. Tests were done on pooled fleas collected from PDs and burrows, serum from live PDs, and tissues from dead PDs (NT = no tests).
PD, prairie dog.
For assessments of annual survival, we first conducted an omnibus evaluation, using a combined dataset for all three species of PDs. We also evaluated each species separately because our study sites were widely separated geographically, were different ecologically, and the three species are biologically distinct in several important ways (e.g., hibernation, social behavior) (e.g., Lehmer and Biggins 2005). We used the iterative numerical optimization routines of program MARK (White and Burnham 1999) on a dataset with complete capture histories for each PD captured at any time during the study (which included multiyear consecutive recaptures and alternate year recaptures) to simultaneously estimate probabilities of capture and survival. We modeled survival to assess variation due to flea control (dust, the primary treatment), species, age, sex, and various interactions among these variables. We attempted to simplify general models to the most parsimonious submodels that explained meaningful variation (Lebreton et al. 1992). Aikaike's Information Criterion (AIC), adjusted for small sample sizes (AICc), was used for model selection (Burnham and Anderson 1998), but likelihood ratio tests provided additional information regarding effects of our primary treatment and covariates. We considered models with the lowest AIC value and models with AIC values within 3 units of the lowest value as competitive, but present all models within 5 AIC units of the highest ranking model.
Results
We collected 6882 fleas from PDs and their burrows. Most were Oropsylla tuberculata, Oropsylla hirsuta, Pulex simulans, and Thrassis francisi (the latter from UPDs at high elevations). Flea control using deltamethrin was highly effective. About 1 month after treatment, captured UPDs had 98% fewer fleas on dusted sites compared to nondusted sites (χ
2 = 86.319, p < 0.001) and WTPDs had 96% fewer fleas after 25 days (χ
2 = 21.737, df = 1, p < 0.001). About 10 months postdust application, UPDs had 85% fewer fleas compared to UPDs on nondusted sites (χ
2 = 85.742, df = 1, p < 0.001), WTPDs had 63% fewer fleas (χ
2 = 104.644, df = 1, p < 0.001), and BTPDs had 45% fewer fleas (χ
2 = 4.158, df = 1, p = 0.041). On nondusted sites, WTPDs were more heavily parasitized by fleas (
Plague detection rates were generally low. Y. pestis was cultured from fleas taken from UPDs, BTPDs, and their burrows only during 2005 (Table 1) and only from colonies that had undergone a population collapse that year (Fig. 2). However, Y. pestis was cultured from 40 pools (of 528 pools tested) of the 4096 fleas taken from WTPDs and their burrows (Table 1). Fluorescent antibody tests detected plague in tissues from 8 of the 54 PDs found dead (Table 1). Similar to flea results, the plague-positive UPD and BTPD carcasses were associated with the colonies that had undergone population collapses in 2005 (Fig. 2). At most sites, few of the 2631 PD samples tested had serum antibodies for plague (Table 1); in 2001, however, 19 of the 48 UPDs tested carried antibodies for plague at the Tom Best Spring colony.
The effective sample size for survival analyses was 2411 PD-years, derived from captures and recaptures of 2211 marked PDs. The sample was heavily weighted toward UPDs (1071 PDs, 1195 PD-years), compared to 710 WTPDs (754 PD-years) and 430 BTPDs (462 PD-years). Power of our WTPD analyses was compromised by the low number of recaptures (51), in contrast to 95 recaptures of BTPDs and 217 recaptures of UPDs.
The omnibus evaluation of PD survival, with pooled data from all species (Table 2), provided strong support for the effect of flea control on survival of PDs, even though epizootic plague did not occur on the study areas during the periods evaluated. All competing models (Table 2, Models 1–3) retained dust, and the effect of dust was underscored by removing it from Model 2 (Table 2) (χ 2 = 17.365, df = 1, p < 0.001). Comparisons of Model 2 (Table 2) with several reduced submodels having pooled species (combining categories of groups), ages (also combinations of groups), and sexes demonstrated influences of those variables (species effect, χ 2 = 79.572, df = 4, p < 0.001; age effect, χ 2 = 26.908, df = 3, p < 0.001; sex effect, χ 2 = 28.412, df = 1, p < 0.001). The differences in survival rates among species were strongly influenced by the low estimated rates for WTPDs (Fig. 3) compared to UPDs (Fig. 4) and BTPDs (Fig. 5). The group × sex interaction in the top-ranking model (Table 2, Model 1) also was influential (Model 1 vs. Model 2, χ 2 = 11.560, df = 5, p = 0.041). That effect was in part due to interactions of species and sex, further justifying separate modeling by species of PD.
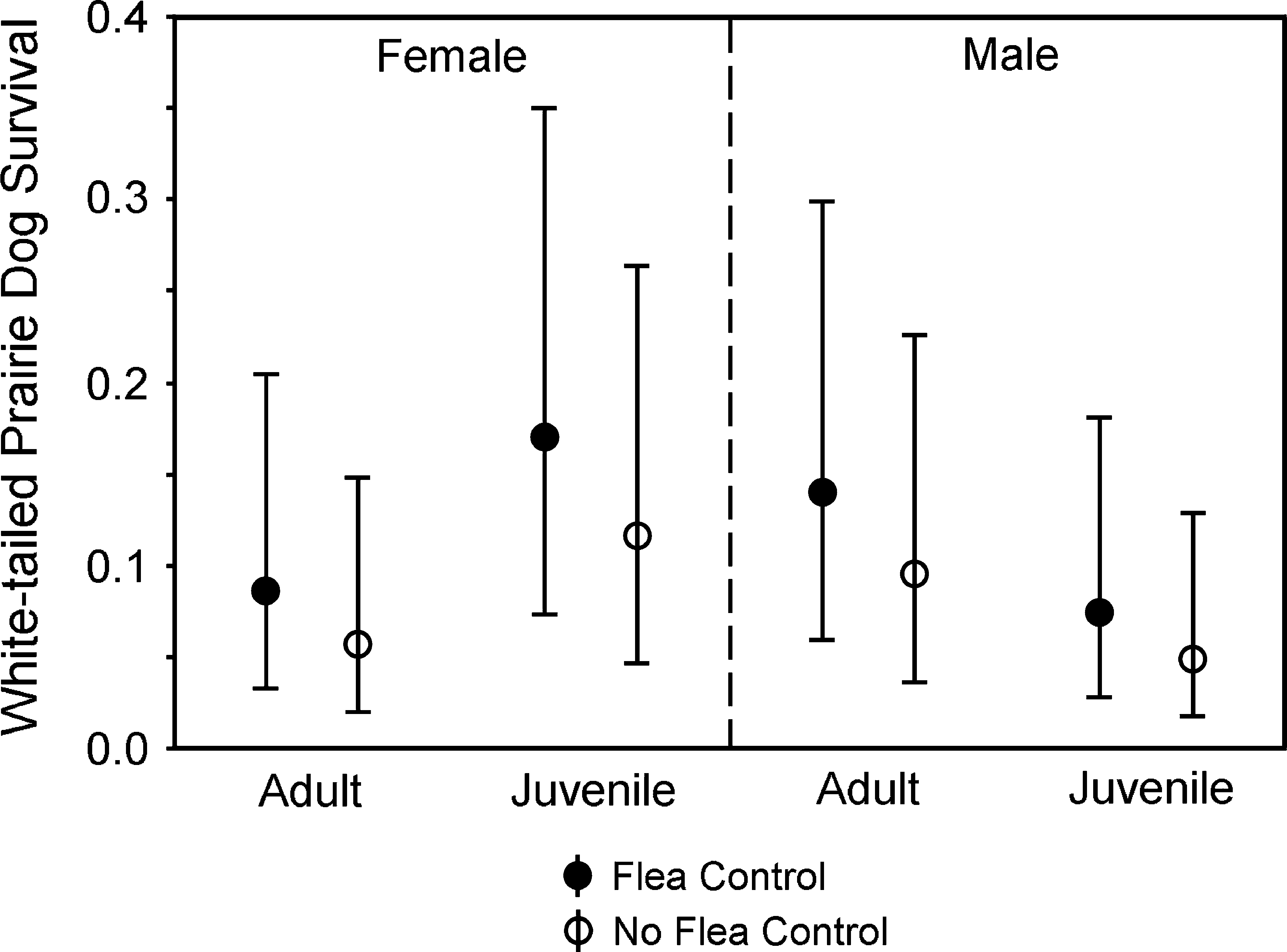
Survival rates (means and 95% confidence intervals) for WTPDs at sites with and without flea control.
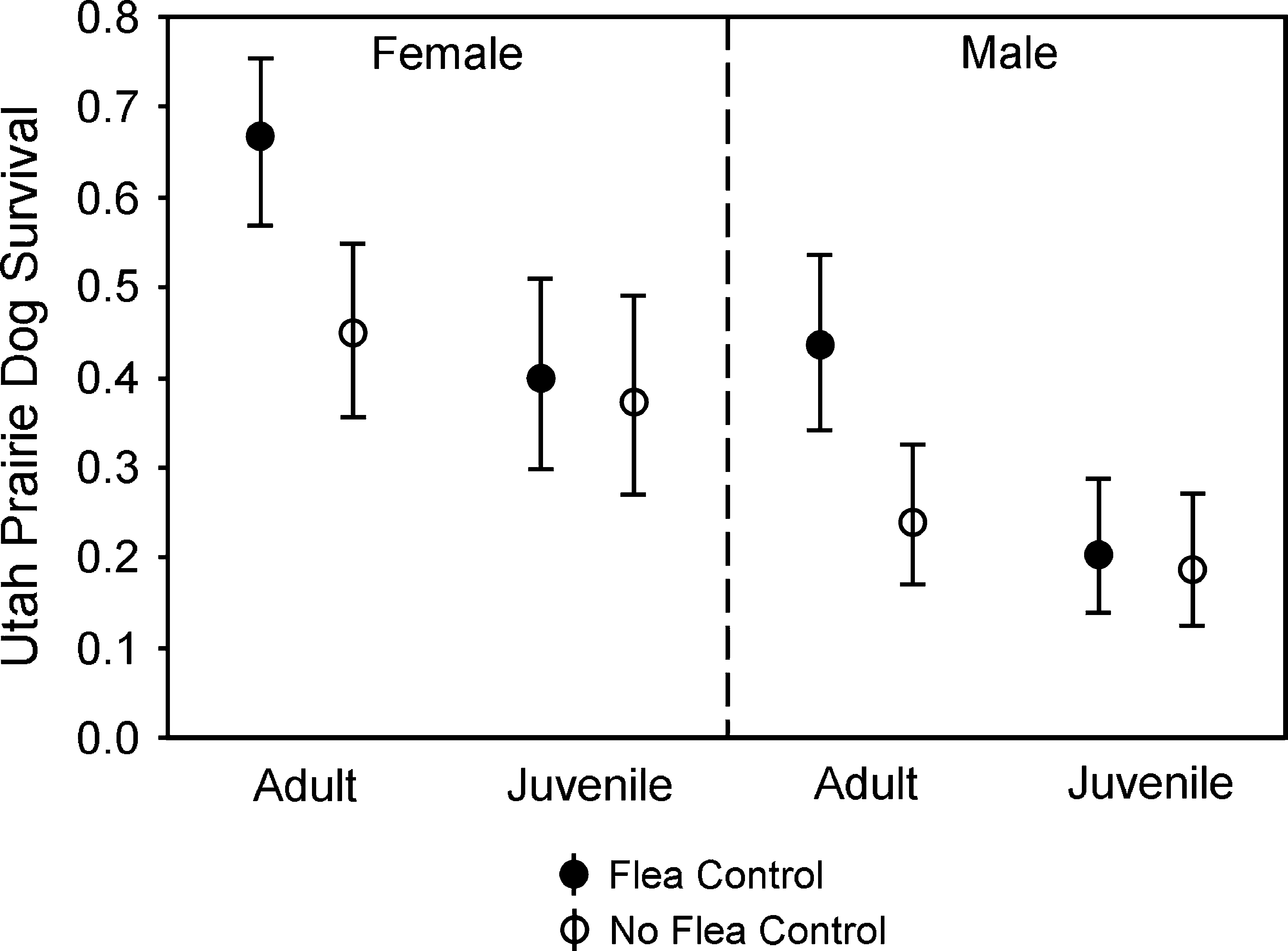
Survival rates (means and 95% confidence intervals) for UPDs at sites with and without flea control.
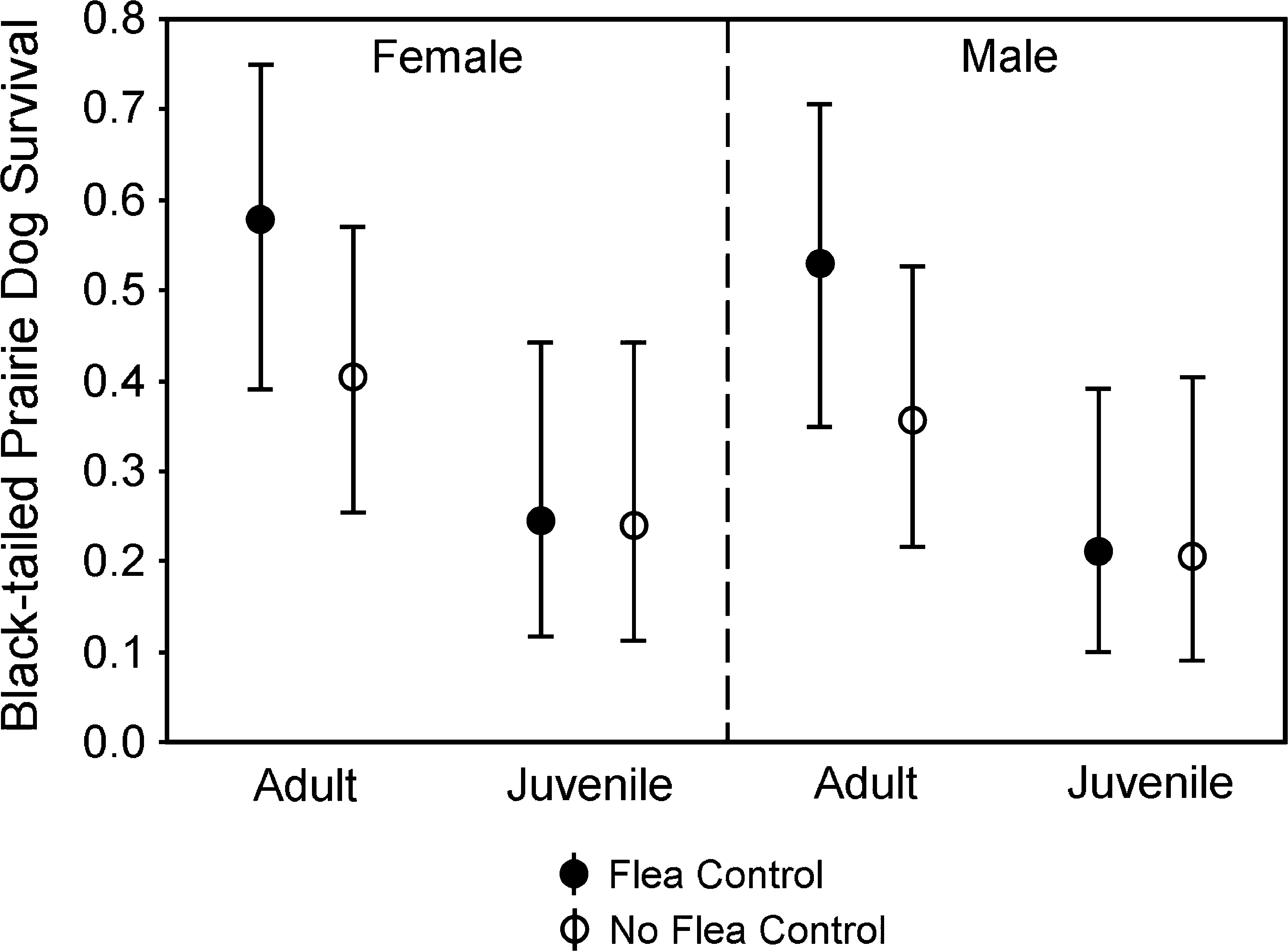
Survival rates (means and 95% confidence intervals) for BTPDs at sites with and without flea control.
This summary presents the highest ranked model and models within 5 AICc units for an omnibus evaluation of all species collectively and presents models from evaluations of each species separately.
Model parameters are Φ = probability of survival, p = probability of capture; (.) = a parameter-specific constant. (,) and (*) indicate additive and interactive effects of variables, respectively. There were two categories of age; juveniles in their first year were separated from adults during their first capture occasion but were combined with adults thereafter. In the omnibus model of all species, group had six categories consisting of three species and two ages. Those groups were collapsed into various combinations of ages and species in models that are not shown because of high AICc values.
AIC, Aikaike's Information Criterion.
Effects of flea control were supported during modeling of each species of PD separately, with the top-ranked model for each species including the effect of dust (Table 2, Models 5, 20, and 27). Survival rates for PDs at dusted sites were 31–45% higher for adults and 2–34% higher for juveniles compared to respective survival rates at nondusted sites (Figs. 3 –5).
Flea control seemed less influential for WTPDs than for the other species (Fig. 3), although as noted above, power was reduced for the WTPD analyses because of low numbers of recaptures. There were numerous competitive models (Table 2, Models 5–15), and dust was not represented in several of them. Removal of dust from the preferred model (Table 2, Model 5, vs. Model 6) provided similarly weak support for its effect (χ 2 = 2.147, df = 1, p = 0.145). The interaction age × sex in the preferred model (Model 5) likely resulted from lower survival of juvenile males compared to juvenile females (χ 2 = 5.252, df = 1, p = 0.022) in contrast to similar survival rates for adults of each sex (χ 2 = 1.174, df = 1, p = 0.279) when age groups were evaluated separately.
Flea control was influential in the models of survival for UPDs (Fig. 3) and BTPDs (Fig. 4). Because the preferred model for UPDs (Table 2, Model 20) contained an age × dust interaction and that model also was competitive for BTPDs (Model 31), we used it to produce estimates of survival for both species (Figs. 3 and 4). Reduction of models to exclude dust substantially weakened the explanatory capability for both the UPD (χ 2 = 11.334, df = 1, p < 0.001) and BTPD models (χ 2 = 3.578, df = 1, p = 0.059). Because of the interaction, age groups were assessed separately for both UPDs and BTPDs using likelihood ratio tests to evaluate submodels as noted above. The influential interaction of age × dust for UPDs (χ 2 = 4.274, df = 1, p = 0.039) was due to a greater degree of improvement in survival (i.e., a steeper slope, Fig. 3) produced by flea control on adults (χ 2 = 11.667, df = 1, p < 0.001) than by flea control on juveniles (χ 2 = 5.817, df = 1, p = 0.016), although both age groups benefitted. The trend was similar for BTPDs (Fig. 4), with noticeable improvement in survival when fleas were controlled on adults (χ 2 = 4.965, df = 1, p = 0.026) but no detectable change in survival due to flea control on juveniles (χ 2 = 0.010, df = 1, p = 0.922).
Probability of capture (p), often regarded as a nuisance variable when the primary focus is on survival rates, is intrinsically interesting. The omnibus modeling and most of the models for each species of PD did not support differences in the probability of capture (p) among species or between sexes, with an overall estimate of 0.429 (95% confidence interval = 0.355–0.510).
Discussion
The consistent lack of effect of variables on p suggests that we were reasonably successful in standardizing the trapping procedures among species. We attempted to keep procedures constant among groups, especially between dusted and nondusted sites within pairs. Indeed, in future studies of this nature, if the interest is primarily in assessing treatment effects within species rather than estimating true survival, estimating p might not be necessary and power might be gained by reducing numbers of parameters estimated in the most general models.
Fleas were effectively controlled by the annual treatments of deltamethrin, with estimated reductions of 96–98% 1 month after treatment and reductions of 45–86% 10 months after treatment. We do not understand, however, why fleas were more abundant on WTPDs than on UPDs or BTPDs, or why flea abundance on dusted WTPD sites was relatively high 10 months after treatment compared to species at the other sites. Because the level of flea abundance at even the dusted sites for WTPDs was higher than levels that appeared to depress survival on the nondusted UPD and BTPD sites, we speculate that we might not have been successful in sufficiently reducing plague effects on WTPDs at dusted sites. Indeed, our observations of more abundant fleas on WTPDs were associated with the lowest survival rates (note the differences in scale of y-axis for Fig. 3 compared to Figs. 4 and 5) and highest detection rates for plague (Table 2) relative to the other PD species.
These relationships are consistent with predictions of Lorange et al. (2005), who modeled the correlation between flea abundance and plague transmission rates and postulated threshold levels of flea density for transition from an enzootic state to epizootic outbreaks. Our average of about 9 fleas collected per WTPD is well above their predicted infestation level to support enzootic plague and near their threshold of 9.4 fleas per host for eruption of epizootic plague. Although intriguing, this alignment with predictions should be cautiously interpreted as their work was with blocking fleas (Xenopsylla cheopis) on laboratory mice compared to common PD flea species that purportedly block less frequently and can transmit Y. pestis without blocking (Eisen et al. 2006, Wilder et al. 2008). Also, our flea sampling did not attempt to capture all fleas; rather, it was intended to provide an index for flea abundance.
Age-specific patterns of survival for UPDs and BTPDs were strikingly similar. One plausible explanation involves indirect effects that result in compensatory losses and increased PD population turnover, which might develop in the following manner: under the premise that (1) enzootic plague causes PD mortality and (2) it is indiscriminate regarding age of victim, it would initially reduce density of both juvenile and adult PDs. The reduction in density of both age classes could create less pressure for dispersal by our juvenile age class, defined as yearlings by the time of their dispersal (Hoogland 1995), increasing their survival and retention rates on the colony. The vacancies caused by losses of adults to plague could be filled by those yearlings that might otherwise have dispersed, leading to a population with more young animals when plague is present than when absent. Thus, the net benefit of reducing fleas might be greater for adults than for juveniles (Figs. 3 and 4) because juveniles replace adults more often when plague is present, but are more often lost to the population through dispersal when plague is absent (i.e., there is no net benefit from increasing survival of PDs that will be lost to other causes). By the time we captured PDs in summer, more yearlings (marked as juveniles) would have dispersed away from dusted than from nondusted colonies under this scenario. The phenomenon might be expected to become increasingly pronounced as the incidence of plague increases. There would be a point, however, at which production of juveniles would not equal mortality rates and the population would decline.
In contrast to the scenario presented above, we could expect direct effects of fleas to result in lower survival for young PDs compared to adults, as reported for other small mammal species (Lehmann 1992, Van Vuren 1996, Hawlena et al. 2006). It thus seems difficult to use direct flea-induced mortality to create an explanation for our observed age × dust interaction and attendant survival estimates.
Our failure to detect plague at many of our study areas might generate doubt regarding its presence. Was the increased PD survival we observed on areas where insecticide was applied due to control of multiple ectoparasites and/or other disease(s), as opposed to reducing the effects of plague? For the following reasons, we think enzootic plague is the most plausible explanation. (1) In the absence of epizootics, plague was detected in fleas, plague antibodies were detected in PD serum, and plague-killed PDs were found. (2) Evidence of the presence of plague was commonly demonstrated in carnivore serosurveys in or near the WTPD (B. Zwetzig, personal communication) and BTPD study areas (Matchett et al. 2009). (3) Nested polymerase chain reaction (PCR) methods have detected apparent enzootic plague near our BTPD study areas (Hanson et al. 2007), although these results were not confirmed by mouse inoculation or other widely used assays. (4) Eruptions of epizootic plague in 2005 at three UPD sites (nondusted) and a BTPD site suggested presence of local enzootic plague in rodent populations, with eruptions perhaps synchronously triggered by favorable climatic factors (precipitation and temperature) (Gage et al. 2008). (5) Direct effects of fleas on other species of rodents often seem benign (e.g., Khokhlova et al. 2002), but when detectable, significant direct effects have been demonstrated only for juveniles (Lehmann 1992, Van Vuren 1996, Hawlena et al. 2006), a trend contrary to our data.
Finally, the most compelling evidence for enzootic plague in PD communities came from a concurrent study of black-footed ferrets, done on study areas that were near our BTPD study areas in Montana. Survival of ferrets in that study was improved 83% by flea control and 91% by an experimental plague vaccine (Matchett et al. 2009), despite lack of epizootic plague in the PD colonies supporting those ferrets. Ferrets are highly susceptible to plague (Rocke et al. 2004, Godbey et al. 2006) and are obligate predators of PDs. It is likely that ferret exposure to plague was from fleas or consumption of PDs or other small mammals killed by plague.
Our general failure and failure of others (e.g., Holmes 2003, Holmes et al. 2006, Matchett et al. 2009) to detect plague in PDs, other rodents, and fleas of rodent communities at these Montana study areas do not convincingly demonstrate absence of plague. In fact, the opposite seems true considering that general carnivore surveys showed evidence of nearly constant presence of plague from 1993 to 2007 in the vicinity of our BTPD and black-footed ferret study areas (Matchett et al. 2009). In addition, using a nested PCR technique, Hanson et al. (2007) reported detecting plague in fleas on 60% of the active PD colonies they sampled during the same time period as our study and at study areas near ours and those of Holmes (2003).
Although the direct effects of fleas seem an unlikely explanation for the altered PD survival in our studies and we believe Y. pestis infections during interepizootic periods might be a significant but not overwhelming source of mortality, we acknowledge that unrecognized pathogens other than Y. pestis could be transmitted to PDs by fleas, ticks, or other ectoparasites and cause mortality. Wild-caught PDs have been reported to be infected with Francisella tularensis (Petersen et al. 2004). Ticks are typically considered efficient vectors of F. tularensis and we occasionally found these acarines on PDs and in PD burrows. Fleas also can transmit tularemia through mechanical means (contaminated mouthparts), which is an inefficient process and unlikely to result in high rates of host to flea to host transmission (Hopla 1974). PDs also frequently are infected with various genotypes of Bartonella washoensis subsp. cynomisii. Other bacteria within this genus have been demonstrated to be transmitted by fleas and B. washoensis subsp. cynomis has been identified in PD fleas by sequencing of PCR-amplified gene products (Stevenson et al. 2003), suggesting that PD fleas could transmit these bartonellae. However, unlike Y. pestis and F. tularensis, B. washoensis subsp. cynomysii has not yet been shown to be pathogenic in PDs and, therefore, seems an unlikely source of mortality in our studies. In addition to the above bacterial species, other vector-borne agents, including as yet undiscovered viruses, bacteria, and rickettsiae, could have been present in our study area and might cause mortality in PDs. Searching for such agents, however, was outside the scope of our study, which was intended only to determine whether vector control could increase survivorship among these animals.
If enzootic plague persists in PD communities, several mechanisms might be involved, alone or in combination. There is potential for soils deep within burrows to provide a reservoir (Baltazard 1964, Ayyadurai et al. 2008, Eisen et al. 2008). Fleas themselves might maintain Y. pestis for long periods (Gage and Kosoy 2005), perhaps interacting with soils (Krasnov 2008) and influenced by their inefficient transmission of Y. pestis (Lorange et al. 2005). Chronic mortality at the population level does not imply chronic infection of individual PDs by Y. pestis. Because of their high susceptibility, PDs probably do not maintain Y. pestis through chronic infections of individuals, although, hypothetically, resistant individuals might be exceptions. The concept of enzooticity of Y. pestis within complexes of PD colonies is not incompatible with other concepts of movement and amplification, but Y. pestis might be less reliant on those mechanisms once it has become enzootic. Also Y. pestis likely benefits from being transported by mammals and their fleas in terms of broad scale spread, and local epizootics might amplify the organism sufficiently that the risk is elevated on the scale of complexes of PD colonies without total decimation of the larger PD population. We hypothesize that source-sink dynamics are also operating in relatively large scales of space and time. For example, plague might be persistent (even within PD communities) where the climate is typically optimal, but might migrate periodically into peripheral habitat for Y. pestis when conditions are most favorable. Conditions conducive for plague propagation appear correlated to weather phenomena at several scales (Parmenter et al. 1999, Enscore et al. 2002, Stenseth et al. 2006, Snäll et al. 2008).
The ramifications of enzootic plague to wildlife conservation are wide reaching. Epizootics are so devastating that their effects are dramatic and obvious. The associated drama tends to draw our attention away from what may be even more important aspects involving enzootic plague. Enzootic plague might have sufficient influence that populations of PDs and other species might not persist over moderate time spans in the absence of catastrophic losses due to epizootic plague. The black-footed ferret in Montana exemplifies a species that is vulnerable to enzootic plague because of its sensitivity to the disease and perhaps its ability to find and consume plague-killed animals, even if the latter are uncommon.
The comparative ease of observing epizootic plague might convince us that failure to detect the disease during interepizootic periods can be interpreted wrongly as absence of the disease. The overall difficulty of detecting plague should provide a warning for those dedicated to wildlife conservation and human health. For species that are difficult to monitor and those that have been poorly monitored, even epizootic plague might occur unnoticed (Biggins and Kosoy 2001a). Hazards from plague may exist even where there have never been epizootics that caused widespread and readily detectable levels of mortality among local rodent hosts. These two points should motivate us to reassess the possible role of plague in population declines of multiple species of conservation concern in the western United States such as the pygmy rabbit (Brachylagus idahoensis), the northern Idaho ground squirrel (Spermophilus brunneus), the riparian brush rabbit (Sylvilagus bachmani riparius), and carnivore species such as the Canada lynx (Lynx canadensis), as indicated by numerous samples collected from dead lynx at reintroduction sites in Colorado and later submitted for analysis to the Centers for Disease Control and Prevention (Wild et al. 2006, CDC unpublished data). Many rodent taxa in the western United States, as summarized by Hafner et al. (1998), should also be considered in this context with special attention to ground-dwelling sciurids occupying fragmented habitats within plague-endemic regions (e.g., Tamias umbrinus nevadensis, Spermophilus lateralis wortmanni).
Footnotes
Acknowledgments
We appreciate assistance from the Centers for Disease Control and Prevention for flea identification and for testing of flea pools, blood samples, and carcasses. We are deeply indebted to our many crew leaders and technicians who assisted them (n = 70). R. Reading and B. Miller of the Denver Zoological Foundation arranged logistical support for the study. Funding was provided by several federal agencies, including the Fish and Wildlife Service, the Geological Survey, and the Bureau of Land Management, supplemented by additional funds from the Utah Division of Wildlife Resources and the Utah Department of Natural Resources Endangered Species Mitigation Fund. Additional in kind support was provided by the Bryce Canyon National Park, Dixie National Forest, and Bureau of Land Management offices in Utah (Vernal, Cedar City, Richfield, and Torrey), Colorado (Meeker), and Montana (Malta). We appreciate constructive reviews of this paper by R. Matchett, D. Eads, P. Stevens, and L. Ellison. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Disclosure Statement
No competing financial interests exist.