
Abstract
We report the prevalence of Siberian and Far Eastern subtypes of tick-borne encephalitis virus (TBEV) in Ixodes persulcatus and Ix. pavlovskyi ticks collected in Tomsk and its suburbs during 2006–2008. The TBEV was detected in 5.7% ticks collected in the city, where Ix. pavlovskyi ticks were dominated and 7.5% ticks from suburban foci with prevalence Ix. persulcatus ticks. Genotyping of the virus showed that Siberian subtype (89.5%) is predominant in individual ticks of Tomsk suburbs; however, the proportion of Far Eastern subtype in two urban sites reached 47%. Phylogenetic analysis demonstrated that Siberian subtype variants from individual ticks were quite divergent and original. Only one subclade was found to be similar to Zausaev strain of TBEV, which is the etiological agent of lethal chronic form of tick-borne encephalitis infection. The average level of homology of 5′ noncoding region (5′-NCR) of TBEV in the individual ticks was 95% for Far Eastern subtype and 89% for Siberian subtype of TBEV. Multiple substitutions in 5′-NCR were found in viral RNA derived from individual ticks. The A2 and C1 elements of Y-shaped structure and putative site for viral RNA polymerase were most variable regions for TBEV 5′-NCR. The B1 and B2 elements and the start codon were practically conserved. The viral RNA from three TBEV-infected pig kidney embryo cells after three passages (out of 21 polymerase chain reaction–positive ticks) were found to multiple substitutions in 5′-NCR in comparison with viral RNA from individual parent tick. However, these three variants did not replicate efficiently in pig kidney embryo cells that may be connected with a considerable modification of Y-shaped structure of 5′-NCR. The efficiently replicating isolate Kolarovo had only seven substitutions in the 5′-NCR and typical Y-shaped structure for Siberian subtype of TBEV. Our data support the idea that hypervariability of the 5′-NCR reflects viral strategy to select the fittest RNA molecule for productive viral infection in mammalian and tick cells.
Introduction
Approximately 700 million people in more than 30 countries of Europe and Asia (excluding China) live in the areas where TBEV is endemic. The estimated annual incidence of tick-borne encephalitis (TBE) is 14,000 cases (excluding China). From this number, 11,000 TBE cases occur in Russia (Gritsun et al. 2003a). Data on TBE incidence in China are scarce; the only report describes 3500 cases in 1994 (Suss 2003, Lu et al. 2008). Clinical history of TBE infection in humans varies significantly depending on the viral subtype. The infection with the European subtype commonly manifests as a mild-fever form of the disease with low mortality rate of 0.5–1% (Haglund and Gunther 2003). In contrast, Far Eastern subtype of TBEV causes severe meningoencephalitic and encephalitic form with high mortality rate of up to 30% and long-term neurological consequences among the survivors (Dumpis et al. 1999, Heinz and Kunz 2004). Patients infected with the Siberian subtype of the virus suffer from a less severe disease, in comparison with Far Eastern subtype, but tend to develop chronic TBE (Gritsun et al. 2003b). Geographically, Far Eastern subtype of TBEV is generally distributed in the Far East of Russia, whereas European subtype is mainly found in Europe. Siberian subtype is mainly found in European part of Russia, in Ural Mountains region and Siberia. Previously, in addition to the Far East of Russia, the Far Eastern subtype has been discovered in Siberia and Europe as well as in a mix of the different subtypes in European and Asian parts of Russia (Lundkvist et al. 2001, Pogodina et al. 2007, Zlobin et al. 2007).
The TBE is an infectious zoonotic disease that occurs in the so-called natural foci, high-risk, or endemic areas. The virus is maintained in natural foci in a life cycle linking ticks and wild vertebrate hosts. Although multiple tick species are biologically competent to transmit TBEV in a laboratory, in the natural setting TBEV is predominantly transmitted by Ixodes ricinus in Europe and by Ixodes persulcatus in the area from Ural Mountains to the Pacific coast of Asia (Charrel et al. 2004). Since ticks remain infected throughout their life cycle, the epidemiology of TBE is closely related to the ecology and the biology of tick hosts (Randolph et al. 2000, Gould et al. 2003). The prevalence of ticks infected with TBEV in endemic areas in Europe usually ranges from 0.5% to 5% (Suss et al. 2002), whereas prevalence as high as 40% has been reported in Russia (Gritsun et al. 2003a). Small rodents develop a short 2–3 day viraemic phase, which results in a low viraemic transmission to ticks, suggesting that the virus easily adapts to replication in both tick and mammalian cells.
One of the proposed mechanisms of such adaptation is linked to the secondary structure of the noncoding regions (NCRs) on both ends of the viral RNA. Viruses of the genus Flavivirus have highly characteristic structures of the 5′-NCR and the 3′-NCR. The 5′-NCR and 3′-NCR interact with each other to circularize viral genome, which is proposed to be important for RNA replication and for transmissibility between vertebrate and invertebrate hosts (Thurner et al. 2004, Gritsun and Gould 2006). The 5′-NCRs vary in length, sequence, and specific conformation in the mosquito-borne, tick-borne, nonvectored, and nonclassified flaviviruses (Gritsun and Gould 2007). The essential elements of 5′-NCR of TBEV secondary structure are depicted in Figure 1: Y-shaped structure, two conserved sequences (CS), and the translation start codon (Mandl et al. 1993, Gritsun et al. 1997, Khromykh et al. 2001). The Y-shaped structure has been recently suggested to serve as a promoter for the viral RNA polymerase. Specifically, the A1/A2 elements and the loop of the left shoulder of Y-shaped structure interact with the RNA polymerase (Filomatori et al. 2006). The CS A and CS B together with the corresponding elements of 3′-NCR are required for genome cyclization and panhandle formation (Mandl et al. 1993). Mutations in CS A and CS B block virus replication even if they form predicted stable complementary structures with 3′-NCR (Alvarez et al. 2008). Thus, variability of NCRs may be attributed to the strategy employed by the virus to select RNA molecule capable of productive a viral infection in different hosts (Casati et al. 2006).
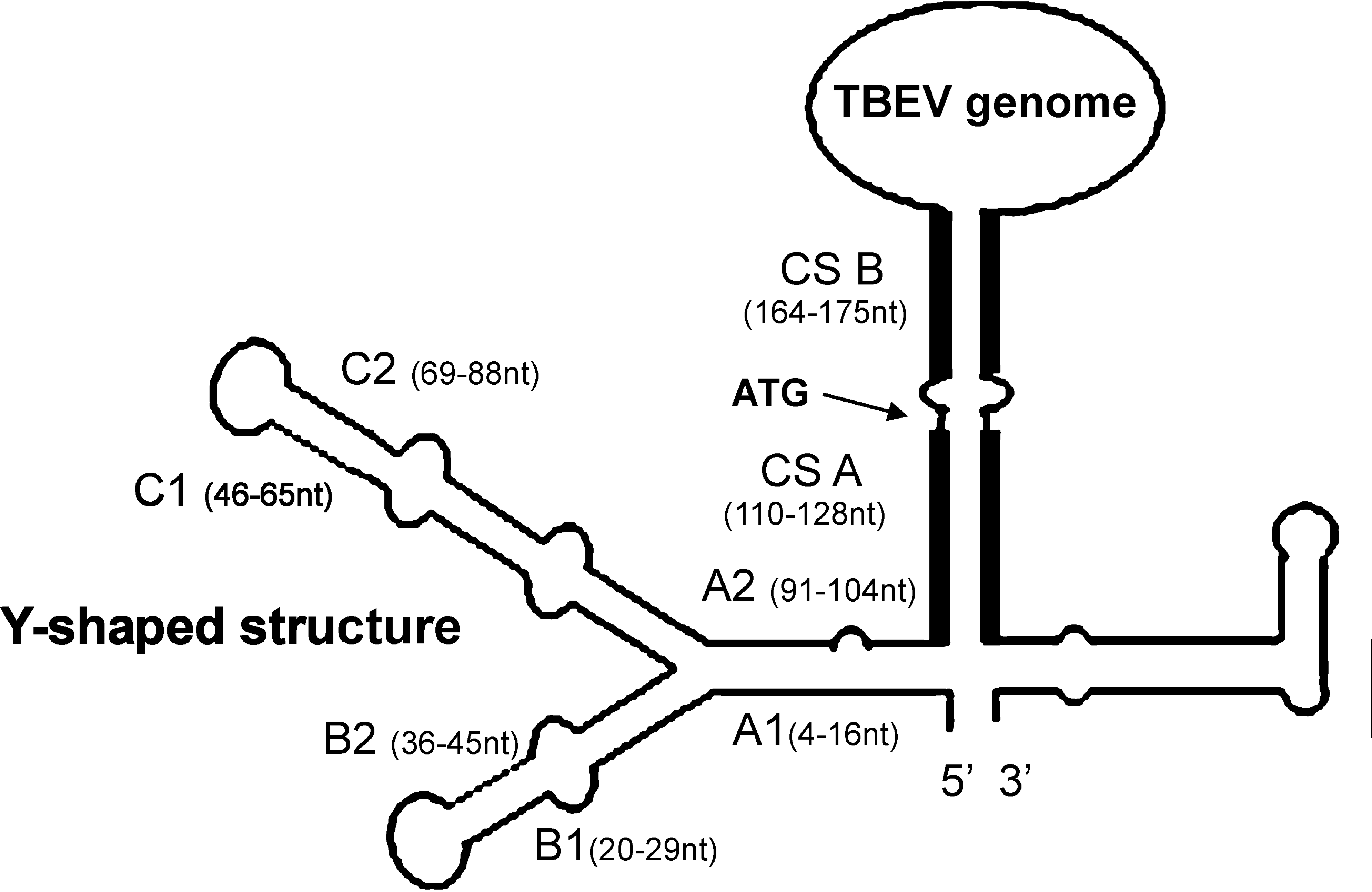
The predicted secondary structure of the tick-borne encephalitis virus (TBEV) genome, calculated by MFold (
Recently, relatively high variability of up to 55.5% in 5′-NCR was reported for the European TBEV subtype derived from the individual Ix. ricinus ticks (Casati et al. 2006). In particular, four RNA regions (CS A, CS B, the folding-stem structure, and the start codon) were shown to display substantial heterogeneity. The authors hypothesized that such variability is linked to the virus adaptation to replicate in mammalian and Ix. ricinus hosts in local natural foci in the central Switzerland. This geographical location is characterized by low incidence of the TBE in humans and high prevalence of TBEV in ticks.
The city of Tomsk with its suburban area (south of Western Siberia) is a highly endemic focus for Siberian subtype TBEV characterized by 10-fold higher morbidity than the average in Russia (Onishchenko et al. 2007). In this study, we present the investigation of the genetic diversity in 5′-NCR region of TBEV found in Ix. persulcatus and Ix. pavlovskyi ticks collected in the Tomsk city and its suburban area.
Materials and Methods
Tick sampling
During spring 2006–2008, ticks Ix. persulcatus and Ix. pavlovskyi (adults, nymphs, and larvae) were collected by flagging or from small mammals. Ticks were stored at −70°C until taxonomic identification, nucleic acid extraction, and virus isolation. The collection was done in two urban and two suburban sites described previously (Moskvitina et al. 2008).
RNA extraction and reverse transcription
Individual ticks were frozen in liquid nitrogen, crushed by plastic pestles, and homogenized in phosphate-buffered saline (200 μL). Total RNA was extracted from 100 μL of the homogenate using RIBO-sorb kit (InterLabService, Moscow, Russia) according to the manufacturer's instructions. Total RNA was transcribed to cDNA using REVERTA-L kit (InterLabService) according to the manufacturer's instructions.
Detection of TBEV cDNA
Polymerase chain reaction (PCR) primers TBE1f (AGATTTTCTTGCACGTGCRTGCGTTTG) and TBE2r (CCCAKCATGCGCATCAAC) were designed based on the multiple sequence alignment of 5′-NCR and protein C-encoding regions of TBEV strains available in GenBank.
Sequencing of PCR products, and structural and phylogenetic analysis
PCR products were separated in 1.5% agarose gel and gel-purified using Wizard SV Gel and PCR Clean-Up System kit (Promega, Madison, WI) according to the manufacturer's instructions. Sequencing was performed using BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Warrington Cheshire, UK) on Applied Biosystems 3130XL genetic analyzer according to the manufacturer's instructions. All samples were analyzed twice in independent experiments. Multiple sequence alignment and phylogenetic studies were performed using Vector NTI Suit 10.0 and MEGA 4. Analysis of the secondary structures and calculations of dG were performed using MFold v.3.2 (
Virus isolation
The tick homogenate diluted 1 to 10 was used to infect a pig kidney embryo (PKE) cells. The infected cells were incubated at 37°C for 4 days in Eagle's minimum essential medium supplemented with 2% fetal bovine serum. The cell culture supernatant was used to infect the next passage of PKE cells. The viral RNA for sequencing was purified from infected PKE cells after the third passage.
Results
Detection of TBEV in ticks
We collected totally 892 Ix. persulcatus ticks (303 female, 280 male, 103 nymph, and 206 larvae) and 914 Ix. pavlovskyi ticks (120 female, 116 male, 145 nymph, and 533 larvae), which were tested for the presence of TBEV by PCR amplification of the 240 nt fragment of 5′-NCR and N-terminus of protein C. Viral RNA was detected in 118 tick samples (in average 6.5%), including 62 positive Ix. persulcatus ticks and 56 Ix. pavlovskyi ticks. The TBEV was found more frequently in nymphs (12.3%) than in imago (8.5% in female and 5.1% in male) or larvae (1.1%). The virus was detected in 5.7% ticks collected at the urban sites and 7.5% ticks from the suburban sites. Ix. pavlovskyi ticks were found predominantly in the urban setting, whereas Ix. persulcatus ticks in the suburban setting.
Phylogenetic analysis
To determine viral subtypes in the infected ticks, we aligned 36 sequences of the amplified fragments from Ix. persulcatus and Ix. pavlovskyi individual ticks, 4 sequences for cell-adapted variants with 2 sequences from Ix. ricinus, and 14 laboratory-adapted TBEV sequences available from GenBank. This analysis demonstrated that 27.8% and 72.2% of these strains belong to Far Eastern and Siberian subtypes, respectively (Figs. 2 and 3). The majority of ticks (89.5%) collected in Tomsk suburbs were infected with the Siberian subtype. The distribution of the subtypes in ticks collected within the Tomsk city limits was found to be more homogeneous, 53% Siberian subtype and 47% Far Eastern subtype.
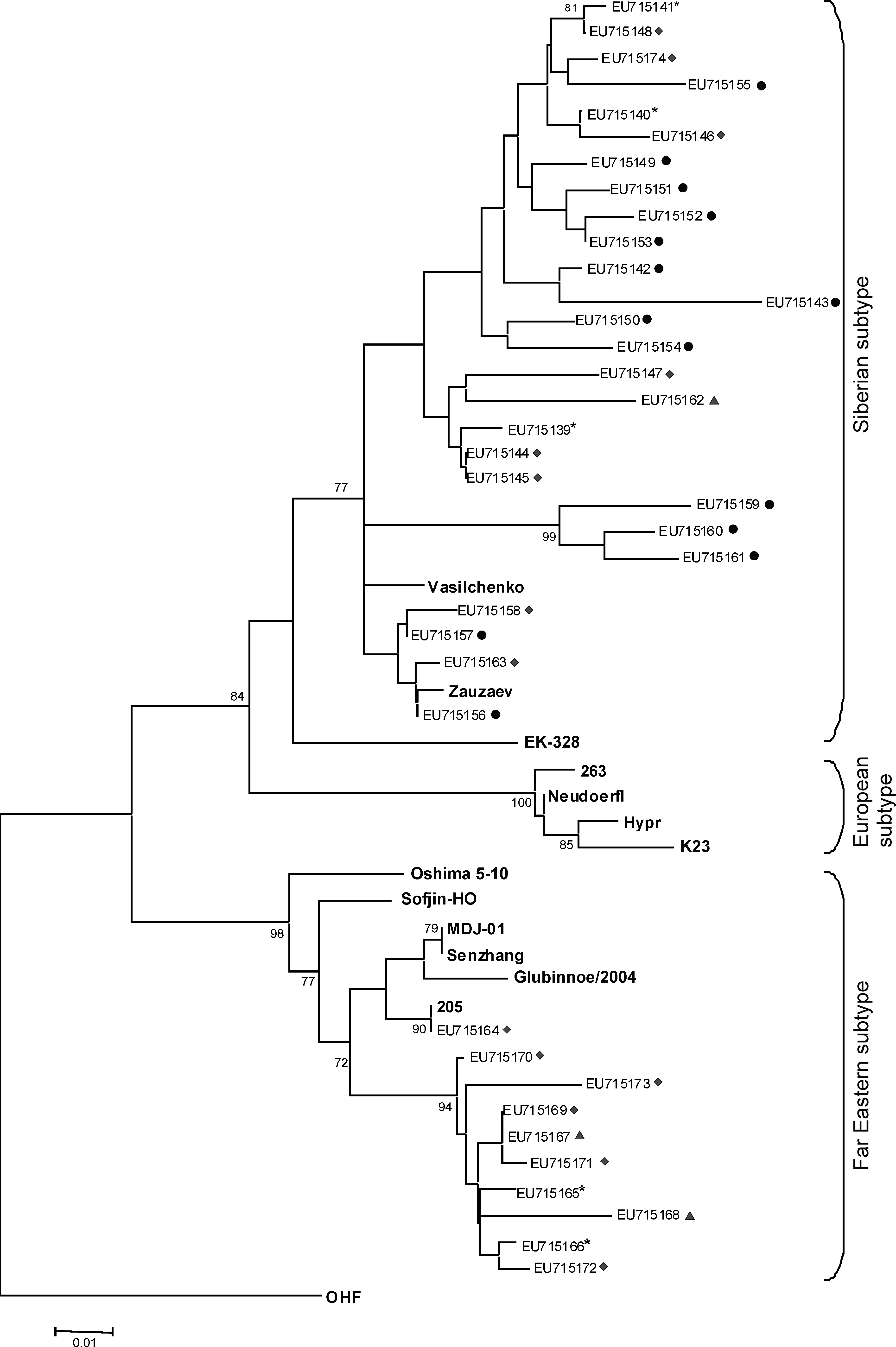
The phylogenetic tree (neighbor-joining, Kimura two-parameter distances) of 36 TBEV sequences from individual ticks and 14 prototype strains of TBEV (bold). Bootstrap is shown when >60%. Omsk hemorrhagic fever (OHF) (AY193805) was used as an outgroup. *, Tomsk oil refinery plant (TORP) suburb site; •, Kolarovo suburb site; ♦, Old necropolis (ON) city site; ▴, Tomsk State University (TSU) city site.
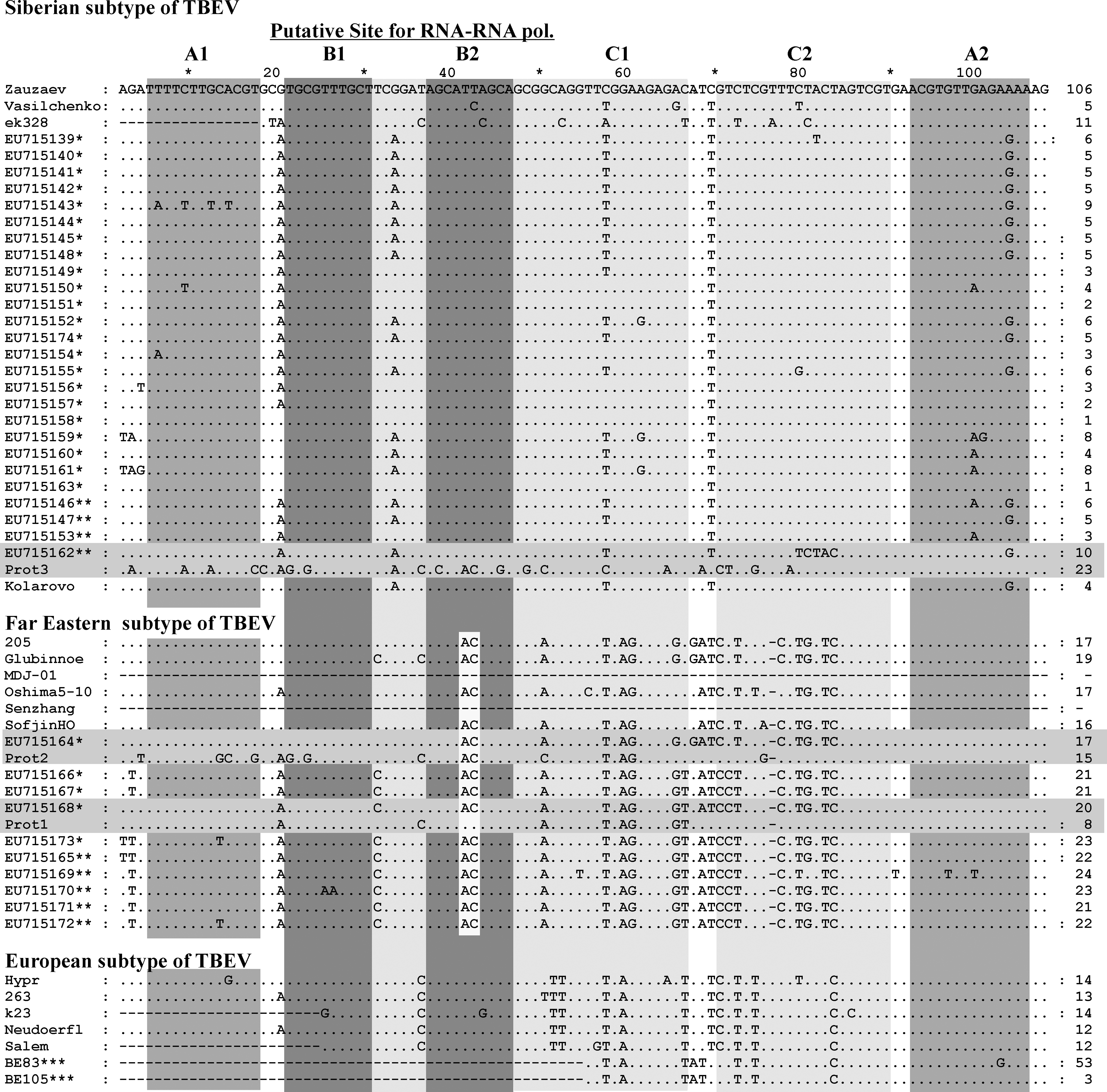
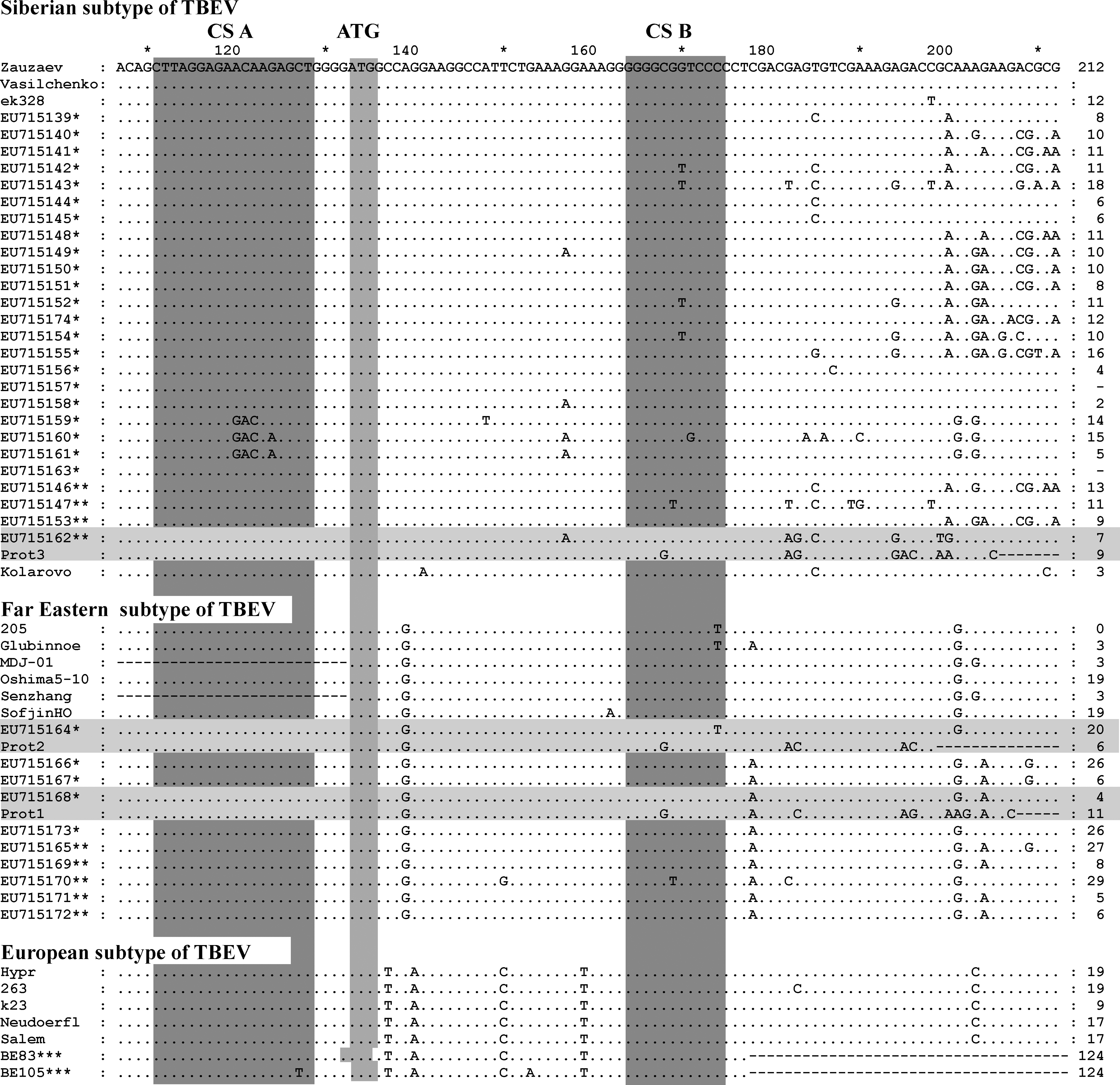
The alignment of 5′-noncoding region and N-end of protein C of TBEV sequences directly derived from ticks collected in the field- and laboratory-isolated strains of TBEV. The elements of the Y-shaped structure, conserved sequence (CS) A, CS B, and start codon are marked by white-gray columns. Far Eastern subtype-specific substitution AC at position 40–41 is marked by white column into very conserved B2 element (marked white–gray column). *, viral RNA isolated from Ixodes persulcatus; **, from Ixodes pavlovskyi; ***, from Ixodes ricinus ticks. The sequences from ticks (parent, not adapted) and after three passages in pig kidney embryo cell: parent (EU715168) for Prot1 (EU715174), parent (EU715164) for Prot2 (EU715175), and parent (EU715162) for Prot3 (EU715176) are marked with white-gray lines. The sequences of from Ix. ricinus ticks BE83 (AY945365) and BE105 (AY945327) were described by Casati et al. (2006), and the sequence Salem (FJ572210) isolated from the brain of a naturally exposed monkey (Macaca sylvanus) in Germany. The sequence Zausaev (AF527415), Vasilchenko (L40361), ek328 (DQ486861), 205 (DQ989336), Glubinnoe/2004 (DQ862460), MDJ-01 (AY217093), Oshima 5–10 (AB062063), Senzhang (AY182009), Sofjin HO (AB062064), Hypr (TEU39292), 263 (TEU27491), k23 (AM600965), Neudoerfl (TEU27495), and Kolarovo/2008 (FJ669215) of laboratory-isolated strains of TBEV were used. The GenBank numbers for other TBEV sequences directly derived from ticks are presented in the figure.
We have uncovered a wide genetic diversity in the primary sequences of the Siberian subtype viruses. Only four primary sequences grouped together with the previously described strain Zausaev, whereas all the other sequences did not cluster with the reference sequences (Fig. 2). Field isolates of the Far Eastern subtype of TBEV were less divergent. One TBEV sequence was homologous to Far Eastern strain 205, isolated in Khabarovsk region of Russia. The remaining nine Far Eastern sequences formed a separate subclade.
Sequence analysis of the 5′-NCR region
The alignment of 5′-NCR sequences revealed multiple mutations in the Y-shaped structure as well as the CS A and CS B elements, but no changes in the start codon (Fig. 3). The average level of homology within 5′-NCR of TBEV from individual ticks was 95% for Far Eastern subtype and 89% for Siberian subtype. The substitution G32 → A in the left shoulder loop sequence of the Y-shaped structure was observed in the majority of sequences of Siberian subtype, whereas T30 → C substitution was observed in the majority of Far Eastern subtype sequences. Table 1 illustrates differential variability of the main 5′-NCR elements. For example, no substitutions were identified in the B2 element and the start codon, whereas the C1 and A2 elements and the putative site for RNA polymerase were a highly variable (up to 80.6%). We also found that substitutions in A2 were more common for Siberian subtype, whereas substitutions in C2 were typical for Far Eastern subtype of TBEV. We did not identify tick species–specific mutations in the viral RNA isolated from Ix. persulcatus and Ix. pavlovskyi.
Number of substitutions was calculated by comparing with strain Zausaev for Siberian subtype and with strain 205 for Far Eastern subtype of TBEV.
TBEV, tick-borne encephalitis virus; 5′-NCR, 5′ noncoding region; CS, conserved sequence.
Subtype-specific nucleotide changes in 5′-NCR of TBEV have some effect on the Y-shaped structure and their predicted secondary conformation (Fig. 4). The prototype and field-isolated strains of Far Eastern subtype have nucleotides AC instead of UU at position 40–41 nt (Fig. 3) and thus form a smoother hairpin in the left shoulder of the Y-shaped structure in comparison with European and Siberian subtypes. Difference of secondary structure of the right shoulder of the Y-shaped structure for Far Eastern subtype TBEV has been induced by multiple subtype-specific nucleotide changes in Far Eastern sequences. However, it is not clear whether or not these conformational changes affect virus virulent characteristics for different subtypes of TBEV.
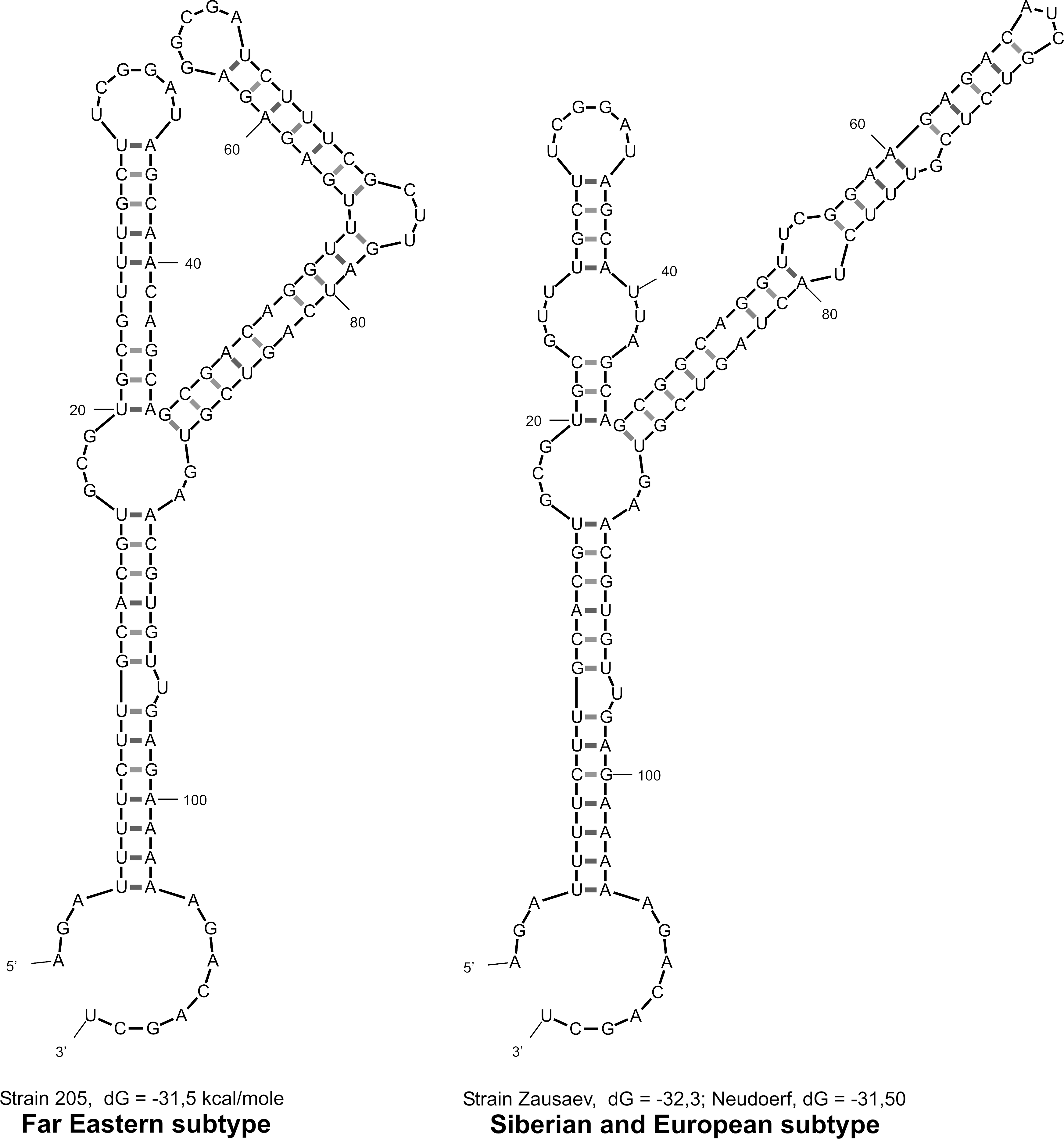
The Y-shaped structures of 5′-noncoding region for the different subtypes of TBEV.
Diversity of the 5′-NCR in TBEV from individual ticks passaged on PKE cells
PKE cells were infected with the 21 individual tick homogenates, which tested positive for TBEV in reverse transcription (RT) PCR. The successful infection, as judged by the detection of the TBEV RNA in PKE cells after three serial passages, was obtained only in four cases named Prot1, Prot2, Prot3, and Kolarovo. The isolates Prot1, Prot2, and Prot3 exhibited low level of replication in PKE cells. The viral RNA and protein E were detected in PKE cells on passages 1 and 3 by RT-PCR and monoclonal modified ELISA, respectively. Both tests were negative on passage 5. The isolate Kolarovo replicated with high efficiency similarly to strain 205 of TBEV as we described previously (Ternovoi et al. 2007). We compared a variability of 5′-NCR sequences of the passaged viral RNAs from Prot1, Prot 2, Prot 3, and Kolarovo with the 5′-NCR sequences of the initial individual tick homogenates used as an inoculum (Table 2 and Fig. 3). We found 14, 22, and 28 substitutions in 5′-NCR in comparison with the original viral RNA from ticks Prot1, Prot 2, and Prot 3 and only 7 substitutions for Kolarovo. Substitutions were found in all 5′-NCR elements with exception of CS A and the start codon.
Isolates were sequenced after three passages on pig kidney embryo cells. Sequencing of the Kolarovo isolate from the polymerase chain reaction–positive individual tick failed because of a low concentration of the viral RNA in the homogenate.
The secondary structure of the Prot1 5′-NCR was similar to the initial viral RNA. The crucial changes were found in the secondary structure of viral 5′-NCRs Prot2 and Prot3 (Fig. 5). We were unable to recover efficiently replicating TBEV from the infected PKE cells after five passages and concluded that observed multiple substitutions in 5′-NCR did not allow these variants to adapt to PKE cells.
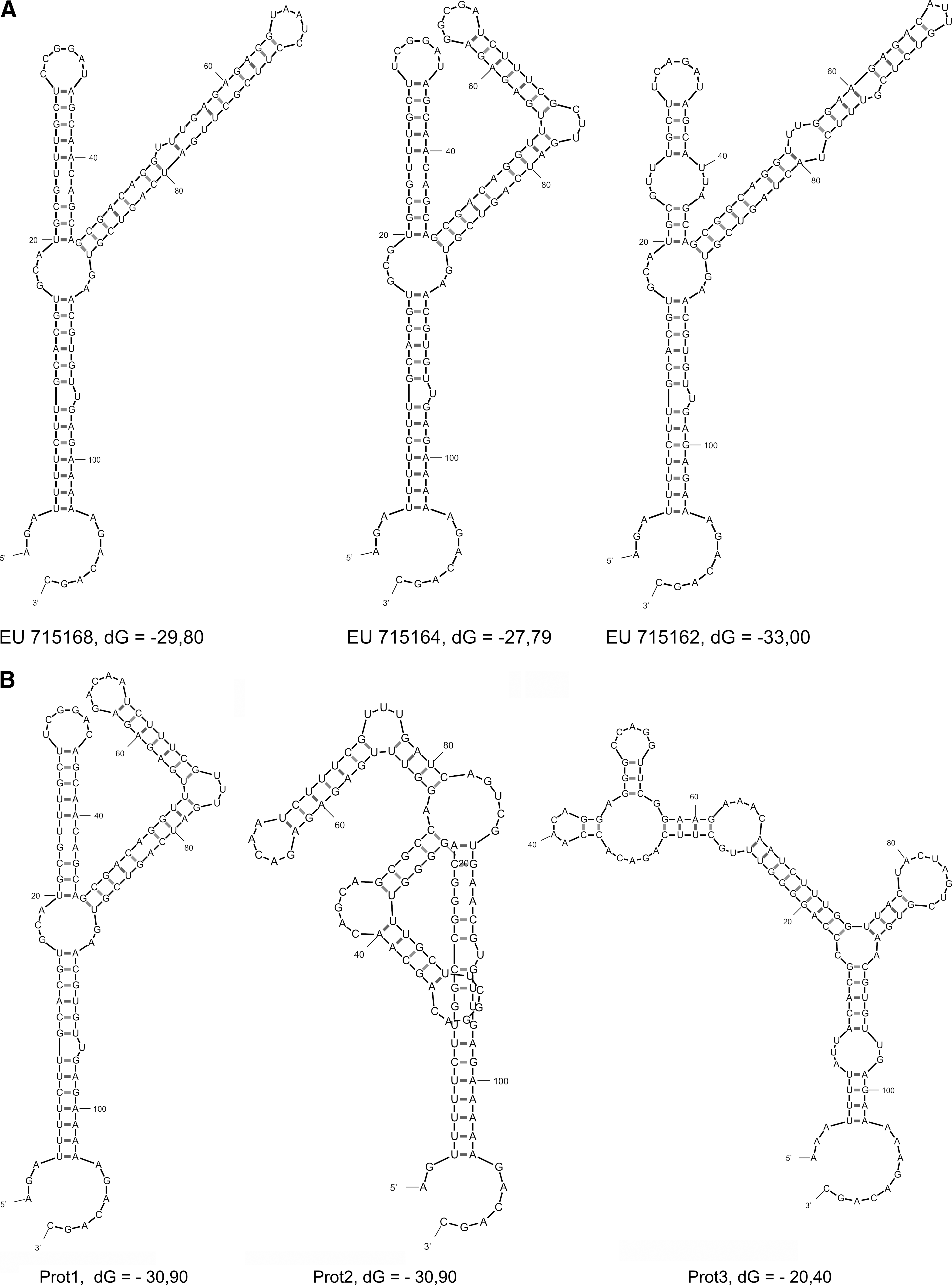
The Y-shaped structures of the field-isolated (
Discussion
Tomsk region is considered to be a highly endemic region for Siberian subtype of TBEV with 10-fold higher morbidity than the average in Russia (Zlobin et al. 2007). The infected ticks collected in the city foci were predominantly Ix. Pavlovskyi, whereas infected ticks from suburban foci were mostly Ix. persulcatus. The viral RNA was found more frequently in nymphs (up to 12.3%) than in imago or larvae. Our genotyping data demonstrate that ticks in Tomsk suburbs are infected with the Siberian subtype of TBEV more frequently (89.5%) than ticks within the city (53%). The wide diversity of the Siberian subtype in Ix. persulcatus and Ix. pavlovskyi suggests that this subtype is endemic for Tomsk region and has a long history of natural evolution. Only four variants from urban and rural sites of Tomsk region were similar to previously known Zausaev strain of TBEV. This strain was isolated in 1985 from a patient with a lethal progressive chronic TBE infection in Moscow (Gritsun et al. 2003b). The patient had suffered a tick bite in Tomsk in 1973. Our result confirms that Zausaev-like strains presently circulate in Tomsk region and may potentially cause chronic TBE infectious in humans.
The relatively high prevalence of Far Eastern subtype of TBEV in urban areas compared to suburbs (47% vs. 10.5%) suggests that the distribution of this subtype is related to human activity. We identified two subclades within Far Eastern subtype. One TBEV sequence was identical to the prototype strain 205, which was isolated in Khabarovsk region in 1957 and now is being used for TBEV vaccine production in Tomsk (Safronov et al. 1991, Leonova et al. 2007). The second subclade of Far Eastern variants is more divergent but clusters together and forms a branch separate from 205-like variant. The level of divergence of among these variants is similar to the divergence of the Siberian subtype. This implies the existence and independent prolonged microevolution for this Far Eastern subclade in Tomsk. Our data suggest several independent incidents of recent and old transmission of the Far Eastern variants from Far Eastern region of Russia into Tomsk urban area.
The mutations in 5′-NCR of the viral RNA of West Nile virus have the potential to compromise replication, translation, and packaging of the flaviviral genome (Yu et al. 2008). Recently, high variability in 5′-NCR (up to 55.5%) was reported for the Western European TBEV subtype derived from individual Ix. ricinus ticks (Casati et al. 2006). In our study the variability of 5′-NCR was found in significantly large percentage of the infected individual ticks. Substitutions in the C1 element were found in 80.6% samples and the putative site for viral RNA polymerase in 72.2%. At the same time the B1 and B2 elements were highly conserved. All these mutations are located in Y-shaped structure of 5′-NCR. However, the predicted secondary structures of 5′-NCR were generally unaffected by these nucleotide mutations (data not shown). In the study mentioned above, high variability of 5′-NCR was found in CS A and CS B, which form the folding-stem structure containing the start codon in between (Casati et al. 2006). We found that the levels of variability for CS A and CS B were lower than in the elements forming Y-shared structure.
The comparison of the 5′-NCR viral RNA sequences isolated from individual ticks with sequences of the viral RNAs from infected PKE cells after three serial passages (Prot1, Prot2, and Prot3 variants) revealed substantial variability in the sequence of the Y-shaped structure. These changes lead to the dramatic alterations in the predicted secondary structure of the 5′-NCR of viral RNAs, and these variants could not adapt to cell culture after five serial passages. Only variant Kolarovo genotyped as a Siberian subtype easily adapted to PKE cells after three passages. The number of mutations in the 5′-NCR of Kolarovo was low in comparison with Prot1, Prot2, and Prot3 variants. These data correlate with high efficiency of replication of Kolarovo in PKE cells. We conclude that the variability in the 5′-NCR may reflect the viral strategy to select the fittest RNA molecule for viral infection of the different host cell types.
The genetic diversity in 5′-NCR of the Siberian and Far Eastern TBEV in two species of ticks suggests relatively long local microevolution of TBEV in Tomsk city and suburban natural foci. We believe that such biodiversity and hypervariability of 5′-NCR of TBEV in natural tick host is the main mechanism for adaptation of TBEV to different hosts, in different environmental conditions, and seasonal climate changes (winter–summer seasons) in Western Siberia.
Footnotes
Acknowledgments
This work was supported in part by the Grant RNP.2.1.1.7515 from State Program for Science Development in High School and Grant 387.2008.4 from the President's Program for Outstanding Scientific School.
Disclosure Statement
The authors have no commercial associations that might create a conflict of interest in connection with this article. All authors are working in nonprofit federal organizations such as State Research Center of Virology and Biotechnology “Vector,” Koltsovo, Novosibirsk Region, and Tomsk State University, Tomsk, Russia. No competing financial interests exist.