
Abstract
The study area, Cuddalore, is one of the endemic districts for Japanese encephalitis (JE) in southern India and there is a strong seasonality in JE case incidence, as well as JE virus (JEV) infection in the principal vector Culex (Culex) tritaeniorhynchus Giles. In a longitudinal 3-year study (July 2003 to June 2006), we determined the susceptibility of wild-caught female Cx. tritaeniorhynchus for JEV infection over several seasons from several villages. The susceptibility varied in all four seasons with the lowest value (4.82 geometric mean [GM]) in hot and wet seasons and highest (13.22 GM) in cool and wet seasons. Infection rate was significant between seasons (7.08–11.85 GM) and years (4.82–13.22 GM). Although the vector was abundant throughout the year, with an average per man-hour density ranging from 58 to 652, the JEV infection rates showed no correlation with vector abundance during different seasons in the index villages. The temporal and spatial changes in the competency of the vector appeared to influence the JEV infection rate in vector, which may at least partially explain the seasonality in JEV human cases in the study area.
Introduction
Materials and Methods
Study area
Our study area, the Cuddalore district of Tamil Nadu, is endemic for JEV (latitude 11.75 N and longitude 79.75 E). Most of the villages of this district are encircled mainly with rice and sugarcane fields and inhabited by farmers and agricultural laborers. In addition to cattle, the villagers also rear pigs and fowl. A total of eight villages in four blocks (villages/block), namely Nallur Block—S.S. Puram and Kodikkalam, Mangalore Block—Avinangudi and Pothiramangalam, Vridhachalam Block—Alichikudi and Sathukudal, and Kammapuram Block—Ko. Adhanur and V. Kumaramangalam, were selected for this study. Meteorological data showed that the maximum and minimum temperatures ranged from 32°C to 39°C and 20°C to 28°C, respectively. The annual rainfall ranged from 0 to 254 mm and relative humidity (RH) ranged between 85% and 92% during the study period. Many of the villages receive irrigation water through the high-level channel from a reservoir, Wellington, where generally farmers raise two rice crops in a year, beginning in June. The villages that are irrigated through the low-level channel lie in the single cropping zone, where a single long-term rice crop is grown, beginning in September/October.
Viral assay
The virus stock used throughout the study was from a single stock. Twenty percentage of mouse brain JEV suspension in 10−3 dilution of the Vellore strain (1958) of India (P20778) passage level P14 in mice was used throughout the study. This 20% JEV mouse brain suspension was serially diluted 10-fold (10−3) with fowl blood and used for oral feeding to Cx. tritaeniorhynchus. The titer of the 20% mouse brain suspension was 106 tissue culture infection dose per milliliter. Chickens used in this study were maintained in a well-protected area free from natural infection. They were periodically tested to find out the infection level.
Mosquito collection
For the susceptibility test in the laboratory, adult female mosquitoes resting nearby cattle sheds and pigsties were collected during dusk hours from the study villages. These collections were repeated during different seasons, namely, hot and wet season (July–September), cool and wet season (October–December), cool and dry season (January–March), and hot and dry season (April–June). All these mosquitoes were identified and sorted to species level and only Cx. tritaeniorhynchus females were used for the infection experiment. Vector density was recorded as females per man hour (PMH). For each season, 800 female mosquitoes (100 specimens from the eight index villages) were infected and tested.
Feeding experiments
Adults of Cx. tritaeniorhynchus females were held in Barraud cages at room temperature of 29°C ± 1°C and 80% ± 5% RH throughout. Water-soaked raisins and cotton pledgets soaked in 1% glucose solution were provided as food. These female mosquitoes were starved for 16–18 h before feeding on JEV blood mixture sweetened with 1% sucrose from cotton pledgets. Female fed on normal blood (uninfected fowl) were kept as control. After feeding, we segregated the fully fed Cx. tritaeniorhynchus mosquitoes. Viremic blood-fed mosquitoes were carefully maintained in the laboratory for 12–14 days at 29°C ± 1°C and 80% ± 5% RH throughout and the water-soaked raisins were provided as food. Special precautions were taken for holding infected mosquitoes in the specially designed mosquito proof cages.
Screening of mosquitoes
Each female specimen was assayed individually for virus content using enzyme-linked immunosorbant assay (ELISA). All blood-fed female specimens were kept individually in microcentrifuge tubes and were triturated in chilled 0.75% bovine albumin phosphate saline (pH 7.4) containing antibiotics (1000 units of penicillin and 2 mg of streptomycin per milliliter). Each mosquito suspension was subjected to two cycles of freezing and thawing and centrifuged at 10,000 rpm for 1 h, and the supernatant was used for screening by ELISA on the same day. Monoclonal antibody, 6B4A-10 (reactive against all the viruses in the JE/WN/SLE/MVE complex), was used as capture antibody and the captured antigen from the test sample was detected by detector monoclonal antibody peroxidase conjugate, SLE MAB 6B6C-1 (Division of Vector-Borne Infectious Diseases, CDC, Fort Collins, CO), reactive against all flaviviruses. A mosquito sample is considered positive for flavivirus antigen if its optical density is equal to or greater than mean + 4 standard deviations of optical density of uninfected laboratory mosquitoes (Gajanana et al. 1995, 1997).
Statistical analysis
The infection rate (IR) is expressed as the percentage of Cx. tritaeniorhynchus females under examination in the feeding series found to contain JEV (Philip Samuel et al. 1998).
The experiments were conducted in a similar fashion in the same place without any detectable variations. These experiments were conducted in different seasons in the same place, using same strains present in those areas, during same period, and using the same stock of the viral strain. Both chi-square test and analysis of variance were carried out using SPSS-15.0 version (Tewari et al. 2008).
Results
The susceptibility of Cx. tritaeniorhynchus to JEV, based on IRs, was lower in the hot and wet season during July–September (2%–8%) and increased subsequently to 10.3%–21% in the following cool and wet season during October–December.
Again the IR in the next season, cool and dry during January–March, decreased to 4.8%–14% and showed an increase during the hot and dry (April–June) season (10.5%–16.8%; Table 1). Susceptibility of Cx. tritaeniorhynchus to JEV from different geographical locations of the study areas (eight index villages) in almost all seasons in 3 years (2003–2006) showed significant difference (p < 0.05), except for one season, cool–dry 2006 (p > 0.05). Infection rate between seasons within different villages and years also showed significant variation (p < 0.05; Table 2).
GM, geometric mean.
Comparison of infection rate between seasons (within a village).
Comparison of infection rate between villages (within a season).
Not significant.
The vector density of Cx. tritaeniorhynchus in PMH was recorded seasonwise during dusk collections from July–September 2003 to April–June 2006 (Fig. 1). The IRs obtained during the different seasons showed no correlation with the abundance of the vector in the index villages, which ranged from 58.61 to 652.2 PMH.
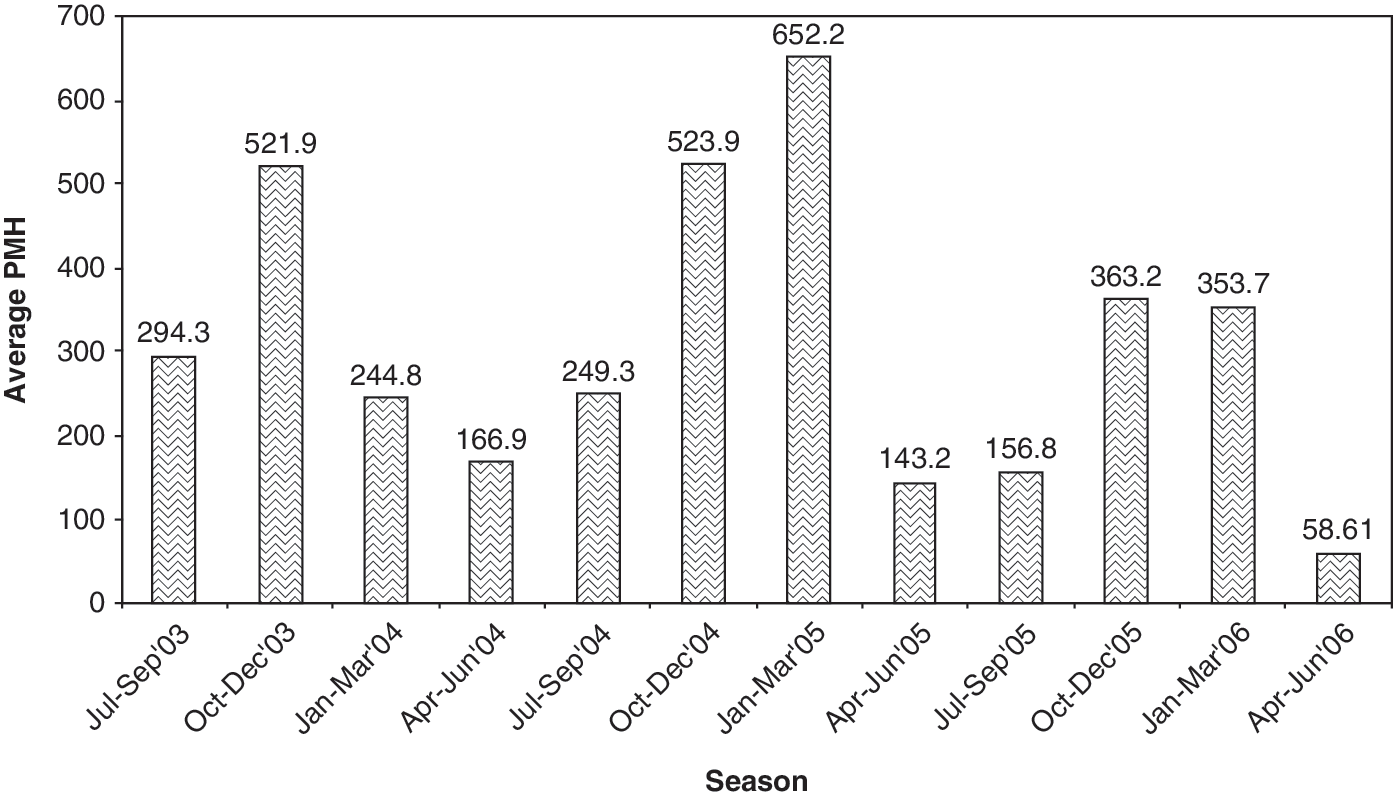
Abundance of Culex tritaeniorhynchus in different seasons in index villages. PMH, per man hour.
Pearson correlation coefficient was evaluated to assess the relationship between the independent (maximum temperature, minimum temperature, and humidity) and dependent variables (PMH density and IR). We have also checked for normality of the variables using one-sample Kolmogorov–Smirnov test and found that the data were normally distributed. As the data follow normal distribution, there is no need for transformation. Multiple linear regressions between IR and all the other meteorological data yielded a multiple correlation coefficient (r 2 = 0.20) showing poor relationship. When the dependent variable was PMH density, a negative correlation exists with maximum temperature (r = −0.634, p = 0.027) and minimum temperature (r = −0.728, p = 0.007), but we did not find any correlation when the dependent variable was IR. The IR showed no correlation with PMH density (r = 0.012, p < 0.970), rainfall (r = 0.147, p < 0.648), humidity (r = −0.065, p < 0.842), maximum temperature (r = 0.073, p < 0.822), and minimum temperature (r = −0.25, p < 0.433) (Table 3).
Correlation is significant at the 0.05 level (two-tailed).
Correlation is significant at the 0.01 level (two-tailed).
PMH, per man hour.
Discussion
In this study, we demonstrated that JEV can infect wild-caught Cx. tritaeniorhynchus females, but the rates vary temporally and geographically. These findings on vector competence help elucidate the transmission pattern of JEV in India, which was similar to other studies using different vectors/arboviruses (Hardy et al. 1979, Hayes et al. 1984, Reisen et al. 1996). In previous vector competence studies in India, most with JEV were limited to only one season and a relatively small sample size with vectors, ranging from 2 to 7 females (Banerjee et al. 1978, 1984). In contrast, our investigation was over four consecutive seasons, included a much larger sample size (100–800) of the principal JE vector, and evaluated relatively small-scale spatial variations from different locations within a JE endemic district. The sample size used in this study was comparable with similar studies conducted in other countries (Freier and Grimstad 1983, Reisen et al. 1996).
Vector susceptibility in different seasons showed a higher IR of 13.22 geometric mean during the cool and wet season (October–December), which coincided with peak transmission season of JE, and a lower IR of 4.82 geometric mean during nontransmission periods (January–March). This suggests that variations in infection during different seasons are mainly responsible for transmission of JEV to humans (Table 1). Thus, temporal changes might greatly impact the efficiency of arboviral transmission in nature, which would have significant epidemiological importance as suggested by Hardy et al. (1979) and Hayes et al. (1984).
In our study, IRs with respect to JEV were similar. Banerjee et al. (1978) have reported similar results with lower IRs in JE vectors. This may be due to the low virus titer and quantity in the blood meal (Rosen et al. 1985) or due to the presence of several anatomical or physiological barriers (Hardy et al. 1983, Weaver et al. 1984). However, it is suggested that the variations in the susceptibility may be due to age distribution of the vector mosquitoes.
Climate variability has a direct influence on the epidemiology of vector-borne diseases (Hardy et al. 1990, Githeko et al. 2000). However, IRs recorded in our study did not show any correlation with the rainfall, temperature, and humidity during different seasons.
Cx. tritaeniorhynchus populations, despite close proximity to each other, varied in their susceptibility to their JEV in different seasons and years. Variations in susceptibility to different arthropod viruses among geographic strains of different populations, including laboratory populations of mosquitoes, had been already recorded in several parts of the world (Gubler and Rosen 1976, Gubler et al. 1979, Banerjee et al. 1983, 1988, Rosen et al. 1985, Boromisa et al. 1987, Vazeille et al. 2001, Mousson et al. 2002, Paupy et al. 2003). We have not examined variation in the population/strain of Cx. tritaeniorhynchus in the study area. However, we observed significant variation in the oral susceptibility in the different study villages. This may not be due to geographical variations as all the index villages are located in one district. This variation in this survey may be due to the spatial variation in larval habitat quality or density of vectors, which will affect the competency of the adults from each index village.
Virus infection in vector mosquitoes also shows a seasonal pattern. The improved understanding of this seasonality will enhance the understanding of host–pathogen interactions and will improve the accuracy of public health surveillance and forecasting systems for the control of communicable diseases and enhancement of preventive health interventions (Fisman 2007). There is a future need to find out the pathways of virus infection and also to quantitatively evaluate the viral particles present in different organs of the vector mosquitoes. The temporal and spatial changes influenced the JEV infection in vector mosquitoes and this would have played a role in the occurrence of seasonality in JEV cases in the endemic study area.
Footnotes
Acknowledgments
The authors are grateful to the Director General, Indian Council of Medical Research, for his support and encouragement in conducting this study. The authors express thanks to the staff members of the Centre for Research in Medical Entomology, Field Station, Vriddachalam, for their excellent technical assistance in laboratory and field work; to S.C. Tewari, for reviewing the manuscript; and to K. Venkatsubramani and A. Venkatesh, for DTP work. The authors are thankful to the Department of Science and Technology, Ministry of Science and Technology, Government of India, New Delhi, for funding this project (SP/SO/C-47/2001).
Disclosure Statement
No competing financial interests exist.