
Abstract
Avian influenza viruses (AIVs) pose a significant threat to public health, and viral subtypes circulating in natural avian reservoirs can contribute to the emergence of pathogenic influenza viruses in humans. We investigated the prevalence and distribution of AIVs in 8826 migratory and resident wild birds in North America along the Pacific flyway, which is a major north–south migration pathway that overlaps with four other flyways in Alaska providing opportunities for mixing of Eurasian and American origin influenza viruses. Overall, the prevalence of AIVs was low (1%) among the wide range of avian species tested, but we detected AIVs in 69 hunter-harvested waterfowl (Anseriformes) sampled at a national wildlife refuge in California from October 2007 to January 2008. A wide range of subtypes were detected in waterfowl with H6N1, H10N7, H7N3, and H3N5 being the most common. We suspect H6N1 was introduced or remerged in 2007 at this key wintering site for waterfowl along the Pacific Flyway. Over a 3-week period, 13 H6N1 AIVs were isolated from two northern pintails (Anas acuta), three northern shovelers (Anas clypeata), three ring-necked ducks (Aythya collaris), four American widgeon (Anas americana), and one gadwall (Anas strepera). We conclude that a diverse array of AIVs was present and that cross-species transmission was occurring among waterfowl in the central valley wetlands of California.
Introduction
The objectives of this study were to characterize the prevalence and distribution of AIVs in resident and migratory wild birds in California and characterize AIV subtypes along the Pacific flyway that may contribute to the emergence of pathogenic viruses in humans. We sampled hunter-harvested waterfowl at a critical migration stopover site in California to identify AIVs circulating in these important reservoir hosts and monitor AIV subtypes that may infect waterfowl migrating south along the Pacific flyway. To characterize AIVs in less studied wild bird populations, we sampled passerines and quails during spring and fall migration seasons, raptors during fall migration, herons and egrets during nesting season, a wide range of avian species admitted to wildlife hospitals.
Materials and Methods
Sample collection
Wild birds were sampled in six California locations over a 3-year period from 2005 to 2008 (Table 1). The Sacramento National Wildlife Refuge (SNWR) complex in the central valley of California (39°26′1″N, 122°11′32″W) is an important site for AIV surveillance because 44% of waterfowl approximately 4 million birds) using the Pacific flyway over winter at the refuge or stop to feed and rest before continuing their southward migration. Hunter-harvested waterfowl (Anseriformes) were sampled five to eight times a month during hunting season (October–February) at the SNWR for all the 3 years. A variety of passerine species and quails were sampled at the Kern National Wildlife Refuge (35°22′23″N, 119°0′51″W) and surrounding areas from April to December 2006. Raptors were sampled during the 2006 fall migration (August–December) over the Marin headlands north of San Francisco (37°46′44″N, −122°25′12″W). Immature herons and egrets were sampled during the 2007 nesting season (June–August) at a rookery near Davis, CA (38°33′15″N, −121°45′44″W). Wild birds entering two wildlife hospitals that receive the maximum number of bird species in California (San Francisco Bay Oiled Wildlife Care and Education Center in Cordelia, CA [38°12′44″N, 122°7′36″W] and the Lindsay Wildlife Museum in Walnut Creek, CA [7°55′26″N 122°4′32″W]) were sampled from January 2006 to June 2007. All birds were sampled within 24 h of admission. Age and sex were recorded for all individuals whenever possible.
San Francisco Bay Oiled Wildlife Care and Education Center, Cordelia, CA.
Lindsay Wildlife Museum, Walnut Creek, CA.
Sacramento National Wildlife Refuge.
Kern county, CA.
Heron and egret rookery in Yolo county, CA.
Golden Gate Raptor Observatory, CA.
Samples were taken from wild birds using a rayon-tipped swab (MicroPur™; PurFybr, Munster, IN) that was inserted into the cloaca and then placed in viral transport media. Additional oropharyngeal samples were taken from birds at the two wildlife hospitals. Cryovial tubes were transported from the field in a cooler with ice packs to a −20°C or −70°C freezer at the collector's institution. Within a month of collection, samples were sent on dry ice to the University of California, Davis, and then transferred to a −80°C for storage.
Sample analysis
A real-time reverse-transcriptase (RT) polymerase chain reaction (rRT-PCR) procedure for detection of the influenza A virus Matrix gene (Spackman et al. 2003) was performed to identify influenza viral RNA in the samples. The primers and probe for the matrix gene and H5 were as described by Spackman et al. (2003). Approximately 50% of our samples were tested first by rRT-PCR (n = 4420), and virus isolation (VI) was attempted on positives. Because VI in embryonated chicken eggs was well established for use in waterfowl (Runstadler et al. 2007), VI (n = 4406) was conducted first on all hunter-harvested waterfowl samples in 9- to 11-day-old specific pathogen-free (SPF) chicken embryos followed by rRT-PCR on the allantoic fluid to positively identify influenza (Swayne et al. 1998, Spackman et al. 2003). Viral RNA was extracted with the MagMAX-96 Viral Isolation Kit (Ambion, Austin, TX) according to the manufacturer's instructions.
To compare AIV detection by VI and rRT-PCR in waterfowl and egrets, a subset of cloacal and oropharyngeal samples (n = 1072) were collected from birds undergoing rehabilitation at the wildlife hospital. Samples were pooled (from five or fewer individuals), immediately transported on ice to the laboratory, and unfrozen samples were tested for influenza A by both rRT-PCR and VI within 5 h of sample collection. Individual samples were archived in a −70°C freezer. These samples were excluded from estimates of overall prevalence for these species.
Subtyping method
For reverse transcription, the first-strand cDNA was made by adding 0.5 μL (1 μg/μL) Uni12 primer (5′-AGCAAAAGCAGG-3′) and 7 μL of RNA to 2.5 μL sterile distilled water. The reaction was heated to 70°C for 5 min, chilled to 4°C, and then added to 4 μL 5× first-strand buffer, 4 μL (10 mM) dNTP mix, 1 μL (10 U) RNasin RNA inhibitor, and 1 μL (200 U) M-MLV polymerase (Invitrogen, Carlsbad, CA) making a total volume of 20 μL. The reaction conditions were 10 min at 25°C, 1.5 h at 42°C, 10 min at 75°C, and 4°C for the remainder of the reaction. PCR for the hemagglutinin (HA) gene was performed using TaKaRa Ex Taq™ according to the manufacturer's protocol (Takara Bio, Otsu-Shiga, Japan) using universal HA primers (Hoffmann et al. 2001, Phipps et al. 2004). The PCR conditions were 94°C for 1 min followed by 30 cycles of 94°C for 1 min, 50°C for 1 min, 72°C for 3 min and then 72°C for 5 min, and 4°C for the remainder. The neuraminidase (NA) gene was amplified using New England Biolabs Taq DNA Polymerase (Ipswich, MA). A multiplex PCR was developed using primer sets designed to amplify all nine NA subtypes (Table 2). To design the primer sets, all published NA sequences were aligned with unpublished sequences from California. The NAs were grouped based on their sequence similarities, and primer sets were designed in conserved regions in groups of two and three subtypes (Table 2). The reaction mixture was made by aliquoting 2 μL of template, 5 μL of NEB 10 × ThermoPol Buffer, 4 μL (10 mM) Invitrogen dNTP mix, 4 μL of total (10 μM) primers, and 0.5 μL of (5000 U/μL) NEB Taq DNA Polymerase to sterile distilled water for a total volume of 50 μL. The PCR conditions were 94°C for 4 min followed by 34 cycles at 94°C for 30 s, 52°C for 30 s, 72°C for 1 min and 72°C for 10 min, and 4°C for the remainder. The PCR product was cleaned using Millipore microcon cleanup columns YM-100, #42413 (Bedford, MA) and submitted for sequencing (Davis Sequencing, Davis, CA). For sequence analysis and alignment, the Invitrogen VectorNTI program was used, and the NCBI database (
Statistical analysis
The χ 2 test of independence was used to detect differences in AIV prevalence among species, sampling locations, age classes, sex, and sampling strategy (capture of free-ranging birds vs. opportunistic sampling of birds at wildlife hospitals). Binary logistic regression was used to predict the probability of AIV shedding for specific groups of species and sampling months for the waterfowl sampled at SNWR. The McNemar's test for paired data was used to detect differences in AIV detection using VI and rRT-PCR methods.
Results
We sampled 8826 resident and migratory birds along the Pacific flyway belonging to 173 species from 19 orders and 45 families (Table 1). The overall prevalence of AIVs among wild birds sampled for this study was low, with only 84 birds (1%) testing positive by VI or rRT-PCR. Of these AIV-positive samples, most (n = 69) were from hunter-harvested waterfowl sampled at SNWR during southward winter migration (Table 3). The remaining 15 AIV-positive samples were detected by rRT-PCR among a wide range of wild bird species tested (Table 4). Subsequent VI attempts on these 15 rRT-PCR AIV-positive samples from highly divergent wildlife taxa using embyonating chicken eggs were not successful and so AIVs infecting these birds could not be subtyped. Highly pathogenic AIV (H5N1) was not detected.
Data are publically available (
Number of positive/number of tested (% positive).
H? = hemagglutinin subtype of these isolates could not be determined by sequencing.
N? = neuramidase subtype of these isolates could not be determined by sequencing.
N/A = hemagglutinin and neuramidase subtypes of these isolates could not be determined by sequencing.
Cattle egret (Bubulcus ibis), snowy egret (Egretta thula), black-crowned night heron (Nycticorax nycticorax), common murre (Uria algae), brown-headed cowbird (Molothrus ater), golden-crowned sparrow (Zonotrichia atricapilla), house finch (Carpodacus mexicanus), house finch (Passer domesticus), and California quail (Callipepla californica).
Heron and egret rookery in Yolo county, CA.
San Francisco Bay Oiled Wildlife Care and Education Center, Cordelia, CA.
Lindsay Wildlife Museum, Walnut Creek, CA.
Kern county, CA.
Prevalence differed among birds (χ 2 = 18.5, p = 0.001) and was highest among waterfowl (Anseriformes, n = 69/4618, 1.5%), followed by herons and egrets (Ciconiiformes, n = 7/763, 0.9%), shorebirds (Charadriiformes, n = 1/380, 0.3%), passerines (Passeriformes, n = 5/1309, 0.4%), and quails (Galliformes, n = 2/658, 0.3%). Even though AIVs were detected in hunter-harvested waterfowl, AIVs were not detected in any of the waterfowl sampled on admission at the two wildlife hospitals (n = 256). Similarly, free-ranging juvenile herons and egrets (Ciconiiformes) caught and sampled at a rookery had a higher prevalence of AIVs, 2% (n = 6/285), than birds of the same species and age class sampled at the wildlife hospitals, 0.2% (n = 1/421, χ 2 = 5.9341, p = 0.015). In fact, 14% (n = 2/14) nestling cattle egrets sampled at the rookery were positive for AIVs by rRT-PCR. Influenza A viruses were not detected via rRT-PCR in migratory raptors at Golden Gate Raptor Observatory (GGRO, Marin Headlands, CA) (n = 482) and wild birds sampled at the wildlife hospital in Walnut Creek, CA, which receives mostly terrestrial bird species (n = 591). Among waterfowl, the prevalence was higher in juvenile, 2.6% (n = 44/1684), than adult 0.8% (n = 17/2207) birds (χ 2 = 21.01, p < 0.001). Prevalence did not differ significantly by sex in waterfowl.
AIVs in hunter-harvested waterfowl
Among hunter-harvested species sampled at SNWR from 2005 to 2007, greater white-fronted geese (Anser albifrons) had the highest prevalence of infection followed by (1) dabbling ducks, including cinnamon teals (Anas cyanoptera), northern shovelers (Anas clypeata), American widgeons (Anas americana), American green-winged teals (Anas carolinensis), northern pintails (Anas acuta), mallards (Anas platyrhynchos), and gadwalls (Anas strepera), (2) sea ducks, that is, bufflehead ducks (Bucephala albeola), and (3) pochard ducks, that is, ring-necked ducks (Aythya collaris) (Table 3). Influenza A viruses were not detected in 18 other hunter-harvested species sampled at SNWR. The waterfowl population at SNWR was sampled repeatedly during fall migration and winter months for all 3 years of this study, and AIV prevalence differed significantly by sampling month in 2005–2006 (χ 2 = 9.0, p = 0.03) and 2007–2008 (χ 2 = 13.1, p = 0.004). During the 2005–2006 winter migration, prevalence was highest in November and February (over 3%) and lowest in December and January. Prevalence in 2007–2008 was highest on October 28 (5%) and December 2 (4%) and very low in January, but sampling efforts were discontinued by February (Fig. 1).
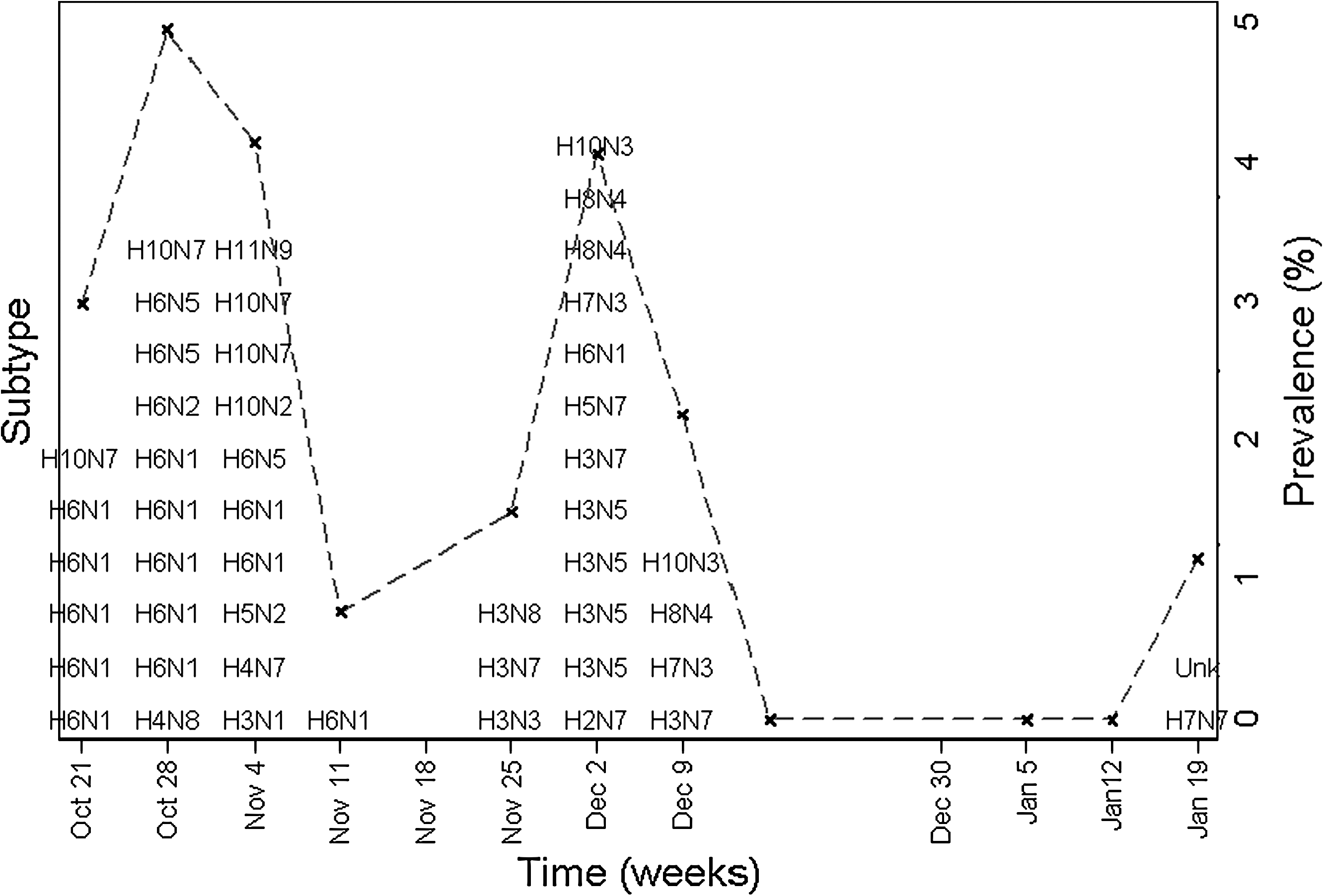
The frequency of avian influenza virus subtypes detected in hunter-harvested waterfowl from October 21, 2007 to January 20, 2008 at the Sacramento National Wildlife Refuge in California. The dashed gray line represents the weekly prevalence and the “X” marks the sampling dates.
Over the 3 years of sampling hunter-harvested birds at SNWR, annual AIV prevalence was 1.1% in 2005–2006 (n = 9/808), 0.8% in 2006–2007 (n = 12/1511), and 2.3% in 2007–2008 (n = 48/2102, χ 2 = 15.9, p < 0.001). Observed prevalence increased more than twofold in 2007–2008 over the prevalence detected in previous years. The majority of positive samples were obtained during two 3-week periods at the end of October and the beginning of December 2007, when observed weekly AIV prevalence rose above 4% (Fig. 1) Avian influenza was isolated from ten different species of waterfowl at SNWR in 2007–2008, but American widgeons and northern shovelers were five times more likely to be infected with AIVs than the other species sampled (p < 0.001).
In total, 69 isolates were characterized from hunter-harvested waterfowl sampled at SNWR. The most frequently detected subtype combinations were H6N1 (21%), H10N7 (10%), H7N3 (6%), and H3N5 (6%), but a total of 26 different subtype combinations were characterized from these 69 samples (Table 3). The frequency of each subtype was significantly different during each sampling year (χ 2 = 63.1, p = 0.009). During the first 3 weeks of sampling in fall 2007 (October 21–November 11), prevalence peaked at SNWR and almost 50% of the AIVs detected were H6N1 (Fig. 1). Waterfowl infected with H6N1 included American widgeons (n = 5), gadwall (n = 1), northern pintails (n = 2), northern shovelers (n = 3), and ring-necked ducks (n = 3). Almost 70% of H10 and 90% of H3 subtypes were detected in northern shovelers. Subtypes H9 and H12-H16 were not detected, and all H5 and H7 isolates were low pathogenic AIVs.
Discussion
Overall, the prevalence of AIVs was very low (<1%) among the wide range of wild bird species sampled throughout California during 2005–2008. The low prevalence of AIVs in waterfowl at SNWR and the higher rates of infections in juvenile waterfowl that are more susceptible than adults (Garamszegi and Moller 2007) are consistent with the previous study reporting AIV prevalence in California waterfowl (Slemons et al. 1974) and similar studies elsewhere along the Pacific flyway in Alaska (Ip et al. 2008) and Mexico (Aguirre et al. 2008). A low overall prevalence and an observed pattern of decreasing frequency in the later months of winter migration (December–January) have been reported in hunter-harvested waterfowl on the Mississippi flyway (Stallknecht et al. 1990, Ferro et al. 2008). We detected the highest AIV prevalence in hunter-harvested waterfowl in the fall at the beginning of southward migration and a second increase in prevalence in February for 2 years of sampling. Pre- and postmigration may be associated with stress and increased susceptibility to AIVs in migratory waterfowl; however, longer term field investigations are needed to confirm seasonal and interannual patterns in AIV prevalence.
A single subtype, H6N1, predominated during the fall postmigration peak in prevalence, and we suspect H6N1 was introduced or reemerged and subsequently circulated among several species of waterfowl at this important wintering ground along the Pacific flyway. This peak was followed by a second spike in prevalence 4 weeks later in early December 2007, which consisted of a variety of other subtypes (Fig. 1). The unexpected sharp increase in H6N1 infections at SNWR could have resulted from the near simultaneous influx of migrating birds that acquired their H6N1 infections elsewhere. However, the diversity of affected waterfowl species and the sustained duration of H6N1 shedding reflects transmission of H6N1 within and among different species at SNWR. The introduction of the other subtypes is most likely due to new migrants into these wintering grounds. The commingling of different species, age classes, and different AIV subtypes all likely play a role in enzootic transmission of AIVs in the central valley of California and elsewhere (Becker 1966, Krauss et al. 2004, Olsen et al. 2006). This study demonstrates that frequent, repeated sampling at the same location is needed to detect the range of circulating AIV subtypes and changes in their prevalence, which is critical for understanding transmission dynamics and AIV ecology.
The subtypes identified in waterfowl in our study areas were similar to subtypes detected in waterfowl in North America and Europe (Olsen et al. 2006), and included all HA subtypes except H9 and H12 to H16. We compared our 3 years of waterfowl surveillance to historical surveillance done in 3-year increments in Alaska (
Infection with AIVs has been detected in shorebirds (waders and gulls) in the orders Charadriiformes and Ciconiiformes (Olsen et al. 2006, Wallensten et al. 2007). Using rRT-PCR, we detected a much higher prevalence (2.7%) in Ciconiiformes than in Charadriiformes (0.2%). Although our sample size was small, nestling cattle egrets sampled at the rookery had a very high AIV prevalence, indicating that young birds, specifically cattle egrets, at this rookery may be a focus of AIV transmission. Other studies have found a range in prevalence, from 0% in nesting great blue herons (Ardea herodias) (Graves 1992) to 32% in nesting gray herons (Ardea cinerea) (Roslaia et al. 1975). We only detected AIVs in a single member of Charadriiformes, the common murre (Uria algae). Infections have been detected in murres in Europe and Alaska (Wallensten et al. 2007, Ip et al. 2008); however, ours is the first report of AIVs in this species in California. Other families (Charadridae, Scolopacidae, and Laridae) within the Charadriiformes have been reported to harbor AIVs, with 14% (Krauss et al. 2004) and 0.5% (Iverson et al. 2008) prevalence in the eastern and western United States, respectively, and less than 1% in northern Europe (Fouchier et al. 2003) and Alaska (Ip et al. 2008). We did not detect AIVs in these families, perhaps because we only sampled 105 individuals (Charadridae, n = 2; Scolopacidae, n = 13; and Laridae, n = 90), and the vast majority of birds from this order were sampled at wildlife hospitals.
Surveillance at wildlife hospitals was a convenient way to estimate AIVs prevalence among sick and injured individuals of a variety of species, but important biases in this sample population must be considered. Waterfowl, egrets, and herons were sampled by free-ranging captures and opportunistic handling at wildlife hospitals, and the prevalence detected in wildlife hospital admissions was significantly lower than their free-ranging counterparts. Compared with the free-ranging birds captured for wildlife studies, birds admitted to wildlife hospitals are biased toward very young age classes as the public often rescues birds that have fallen from the nest or separated from their parents during fledging. Hospitalized animals are also often injured or sick with multiple concurrent conditions that may have minimized their contact with the healthy free-ranging population. In this case, low pathogenic viruses may be underrepresented in the hospitalized population. Conversely, opportunistic screening at wildlife hospitals may be a more efficient way to detect AIVs that are highly pathogenic in wildlife, as sick birds involved in disease outbreaks are often brought to wildlife hospitals by the public.
Prevalence of AIVs is driven by the time of year (migration status), age class, and species present during the time of sampling. At SNWR, hunter-harvested waterfowl were sampled two to six times a month during their time at the refuge, and prevalence estimates shown here should be representative of prevalence during this time period. For passerines and raptors, sampling was discontinued because detection in these groups was very low, but years with higher prevalence could have been missed. Other studies have also found passerines and raptors to have a very low AIV prevalence (Schnebel et al. 2005, Lebarbenchon et al. 2007, Ip et al. 2008), and both may play a minimal role in the transmission of AIVs. However, raptors are extremely susceptible to highly pathogenic AIV (H5N1), indicating that the ongoing surveillance efforts for highly pathogenic AIVs may be worthwhile (Khan et al. 2009).
We detected three subtypes in waterfowl that have been detected in humans previously and caused outbreaks in poultry. The subtype H7N7, isolated from a mallard, caused conjunctivitis and influenza-like illness in poultry workers in The Netherlands (Koopmans et al. 2004) and in a woman in the United Kingdom exposed to ducks (Kurtz et al. 1996). Additionally, a large outbreak in poultry was due to H7N7 (Koopmans et al. 2004). Infection or evidence of exposure to two subtypes detected in shovelers, H7N3 and H11N9, have been detected in humans exposed to poultry and waterfowl (Gill et al. 2006, Myers et al. 2007). Recreational (hunting) and occupational exposure to these waterfowl species may similarly put people at risk for AIV exposure (Siembieda et al. 2008).
Long-term surveillance in California, similar to surveys in Europe and the eastern United States, is needed to better characterize endemic patterns of AIVs and understand the epidemic cycles of various subtypes in a range of species (Krauss et al. 2004, Munster et al. 2004, Boyce 2007, Munster et al. 2007). For Galliformes and Passeriformes, species that are more terrestrial than aquatic, we found an extremely low prevalence similar to other studies conducted in Europe (De Marco et al. 2003a, 2003b), perhaps suggesting that AIV infection in these species is incidental, representing spillover from other species. Future surveillance efforts should include heron and egret species because our results indicated that enzootic transmission occurred at colonial nesting sites. Finally, much more attention should be focused on transmission among resident and migrating waterfowl species in wintering grounds along major flyways that appear to be a focus of enzootic transmission of AIVs.
Footnotes
Acknowledgments
We gratefully acknowledge the expert technical assistance provided by Jerome Anunciacion, M. Veronica Armijos, Rachel Avilla, Michelle Bellizzi, January Bill, Brian Carroll, Robert Cummings (Orange County Mosquito and Vector Control District), Phuong Dao, Devin Dombrowski, Sandy Fender, Buzz Hall (Golden Gate Raptor Observatory), Scott Hallam, Susan Heckly, Grace Lee, Vincent Martinez, Marcia Metzler, Pam Nave, Julie Nelson, Megan Prelinger, Jarl Rasmussen, Shannon Riggs, Annie Schultz, Marie Travers, Sarah Wheeler, Stan Wright (Sacramento/Yolo Mosquito and Vector Control District), and Jean Yim. We also thank the SNWR and California Department of Fish and Game for assistance with logistics and obtaining samples at the refuge. Support for this project was provided by the Avian Influenza Coordinated Agricultural Project (U.S. Department of Agriculture/CSREES grant 2005–35605-15388), “The Prevention and Control of Avian Influenza in the United States” National Institute of Allergy and Infectious Disease, National Institutes of Health, Department of Health and Human Services (Contract no. HHSN2662007 00009C), California Department of Fish and Game's Resource Assessment Program, and the Edward E. Hills Foundation.
Disclosure Statement
No competing financial interests exist.