
Abstract
Chikungunya virus (CHIKV) is a globally emerging arbovirus responsible for unprecedented outbreaks in the western Indian Ocean, the Indian subcontinent and Italy. To assess the receptivity of Australia to CHIKV, we exposed 10 Australian mosquito species to a 2006 strain of CHIKV isolated from a viremic traveler from Mauritius. In susceptibility trials, the infectious dose required to infect 50% of the mosquitoes was 100.6 cell culture infectious dose (CCID)50/mosquito for Aedes procax, 101.7 CCID50/mosquito for Aedes albopictus, 102.1 CCID50/mosquito for Aedes vigilax, and 102.6 CCID50/mosquito for Aedes aegypti and Aedes notoscriptus. When exposed to blood meals containing between 103.5 and 104.1 CCID50/mosquito of CHIKV, infection rates in these five species, plus Coquillettidia linealis, were ≥81%. Subsequent transmission rates ranged between 20% for Ae. notoscriptus and 76% for Ae. vigilax. In contrast, Culex spp. were poor laboratory vectors, with infection and dissemination rates ≤20% and ≤12%, respectively. Although Australia has efficient laboratory vectors, the role a mosquito species plays in potential CHIKV transmission cycles will also depend on its geographical and temporal abundance, longevity, and association with humans.
Introduction
Due to the potential for explosive outbreaks of CHIKV, it is important to estimate the risk of establishment beyond its current known geographical range. The risk of CHIKV spread to Australia is high due to a geographical proximity to endemic areas, such as Indonesia (Laras et al. 2005) and Malaysia (AbuBakar et al. 2007), and the potential for importation via international traffic. Indeed, between 2006 and 2008, there were 43 travelers diagnosed with CHIKV infection entering Australia (Liu et al. 2008, C. Liu and C. Taylor, unpublished data).
The sylvan vectors circulating CHIKV between primates in Africa are tree-hole inhabiting Aedes spp., of which Aedes furcifer, Aedes taylori, Aedes africanus, and Aedes luteocephalus are the most important (Jupp and McIntosh 1988). The primary urban vectors are Aedes aegypti and Aedes albopictus. Of the species implicated abroad, only Ae. aegypti and Ae. albopictus occur in Australia. However, the distribution of both of these species in Australia is limited, with Ae. aegypti occurring in northern Queensland (Sinclair 1992) and Ae. albopictus in the Torres Strait islands, between the tip of Cape York Peninsula and Papua New Guinea (Ritchie et al. 2006). The vector competence of Australian strains of these two species need to be assessed, as geographical variation in susceptibility to CHIKV transmission has been observed previously (Tesh et al. 1976).
Little is known on the vector status of other Australian mosquitoes for CHIKV. In terms of other potential vectors, species that are capable of transmitting the endemic alphaviruses, Ross River virus (RRV) and Barmah Forest virus (BFV), need to be examined for their vector competence. Consequently, in the current paper, we describe vector competence experiments undertaken to determine the ability of Australian mosquito species to become infected with and transmit a Reunion Island strain of CHIKV. This CHIKV strain contains an alanine to valine mutation in the membrane fusion glycoprotein E1 gene (E1-A226V) that has been linked to the high number of cases and severity of disease during the recent outbreaks, and increased infectivity in Ae. albopictus (Schuffenecker et al. 2006, Tsetsarkin et al. 2007, Vazeille et al. 2007). We also describe the development of a rapid, specific, and sensitive real-time TaqMan reverse transcriptase–polymerase chain reaction (RT-PCR) assay for detecting CHIKV in saliva expectorates collected during transmission attempts.
Materials and Methods
Virus strain
A strain of CHIKV isolated from a patient from Mauritius who visited Melbourne in March 2006 was used for all vector competence experiments (kindly supplied by J.D. Druce, Victorian Infectious Diseases Laboratory, Melbourne, Australia) (Druce et al. 2007). This isolate shared 100% homology with other viral sequences from Reunion Island (Druce et al. 2007) and contained the E1-A226V mutation (A.T. Pyke, unpublished data). The stock virus had been passaged three times in African green monkey kidney (Vero) cells and had a final titer of 108.0 cell culture infectious dose (CCID)50/mL.
Mosquitoes
Mosquitoes were collected as eggs, larvae, or adults from several locations in Queensland (Table 1). Adult mosquitoes were collected using Centers for Disease Control (CDC) light traps (Model 512; John Hock Co., Gainesville, FL) baited with CO2 (1 kg dry ice). Adult mosquitoes were transported to the laboratory and exposed to the virus within 12 h of collection.
Ae. aegypti were in the F1 generation.
Ae. albopictus were in the F7 generation.
Larvae of Aedes notoscriptus and Culex quinquefasciatus were collected from discarded containers at a construction site in the suburb of Coopers Plains, Brisbane. The Ae. albopictus used were F7 generation from a colony established from eggs collected from Masig Island in the Torres Strait in April 2005. Eggs and larvae were reared at 27°C, 70–75% relative humidity (RH), and 12:12 light:dark (L:D), and fed Hikari® Cichlid Staple pellets (Kyorin Co. Ltd., Himeji, Japan). Adults were held for 3–5 days on 10% sucrose and were starved for 24 h before virus exposure.
Exposure of mosquitoes to CHIKV
As CHIKV is exotic to Australia, all vector competence experiments were undertaken in the physical containment (PC3) insectary at Queensland Health Forensic and Scientific Services, Brisbane, Australia. To determine susceptibility to infection, mosquitoes were exposed for 2–4 h to serial 10-fold dilutions of CHIKV using a membrane feeding apparatus and pig intestine as the membrane (Rutledge et al. 1964). Dilutions were prepared by adding stock virus to washed defibrinated sheep blood (Institute of Medical and Veterinary Science, Adelaide, Australia) and 1% sucrose. Due to insufficient numbers and variable feeding rates, Coquillettidia linealis and Cx. quinquefasciatus were exposed only to titers of 103.7 CCID50/mosquito and 104.1 CCID50/mosquito, respectively, while Verrallina funerea was exposed to 103.0 CCID50/mosquito. To determine the virus titers during feeding, pre- and postfeeding samples of the blood/virus mixture were diluted 1:10 in growth media (GM; Opti-MEM [Gibco–Invitrogen Corporation, Grand Island, NY] containing 3% fetal bovine serum (FBS), and antibiotics and antimycotics), and stored at −80°C to await titration.
Following feeding, mosquitoes were briefly anesthetized with CO2, and blood engorged mosquitoes were sorted by species, before being placed in 1 L containers within an environmental growth cabinet (Sanyo Electric, Gunma, Japan). Mosquitoes were maintained at 28°C, 75% RH, and 12:12 L:D, and provided 10% sucrose.
Processing of mosquitoes for virus susceptibility and transmission
After an incubation period of 14–15 days, mosquitoes fed the three lowest virus titers were placed individually in 2 mL tubes containing 1 mL of GM +3% FBS and three sterile glass beads. To demonstrate virus transmission, mosquitoes exposed to the highest dose were anesthetized with CO2 and the legs and wings removed, before the proboscis was inserted into a microcapillary tube containing GM +20% FBS (Aitken 1977). Mosquitoes were allowed to expectorate for approximately 30 min, after which the contents of the capillary tube were expelled into a 2 mL tube containing 600 μL of GM +3% FBS. The bodies and legs + wings were placed separately in 2 mL tubes containing 1 mL of GM +3% FBS and three sterile glass beads. All whole mosquitoes, legs + wings, body remnants, and expectorates were stored at −80°C to await analysis.
Virus assay
All pre- and postblood/virus mixture samples were titrated as 10-fold dilutions in a 96-well microtiter plate containing confluent Vero cell monolayers. Plates were incubated at 37°C with 5% CO2 for 7 days, after which they were examined for cytopathic effect. Plates were then fixed with PBS/acetone and stored at −20°C.
Whole mosquitoes, body remnants, and legs + wings were homogenized for 2–3 min using a SPEX 8000 mixer/mill (Spex Industries, Edison, NJ), before being filtered through a 0.2 μm Supor® membrane filter (Pall Corporation, Ann Arbor, MI). The filtrate was inoculated in duplicate onto Vero cell monolayers within a 96-well microtiter plate. Plates were incubated and fixed as described above.
A fixed cell culture enzyme immunoassay (CCEIA) was used to confirm the cytopathic effect caused by virus infection in all blood/virus mixtures and mosquito samples (Broom et al. 1998). As specific CHIKV monoclonal antibodies were not available, an alphavirus cross-reactive monoclonal antibody (B10) was used in the CCEIA (kindly provided by R.A. Hall, University of Queensland, Brisbane, Australia).
CHIKV RNA was extracted from the saliva expectorates using the Qiagen BioRobot Universal System and QIAamp Virus BioRobot MDx Kit (Qiagen, Clifton Hill, Australia).
Real-time TaqMan RT-PCR assay for detection of CHIKV RNA in saliva expectorates
To detect viral RNA in the saliva expectorates collected during transmission attempts, a real-time TaqMan RT-PCR assay specific to the Reunion Island E1-A226V strain of CHIKV was developed. Probe and primer pairs targeting the CHIKV structural envelope glycoprotein E1 were designed as previously described (Pyke et al. 2004). Primers and the dual labeled probe were derived using the sequence of CHIKV Reunion isolate 05-061 (GenBank accession number
In addition, separate synthetic primer and probe positive controls were developed using the method of Smith et al. (2006) with minor modifications. These provided specific, nonviral reagents that eliminated potential contamination by viral RNA controls. Briefly, two high-performance liquid chromatography–purified oligonucleotides with complementary forward and reverse sequences were designed for each primer and probe control. The primer control (forward sequence: 5′-CCCGGTAAGAGCGGTGAA
The primer and probe synthetic controls were prepared individually to produce double-stranded DNA that could be used directly in the TaqMan RT-PCR. Each forward primer and its complementary reverse strand were diluted to 200 μM, combined, and then annealed following a process of denaturation at 94°C for 1 min and hybridization at room temperature for 10 min. Ten-fold dilutions of the synthetic controls were then prepared and titrated individually for use in the TaqMan assay. Briefly, 2.5 μL of each dilution was combined with an equal volume of a synthetic control base mix. For the primer control, the base mix consisted of 1.5 μM of rGAPDH probe (sequence as above) in a final volume of 2.5 μL. Similarly, the probe base mix contained 3 μM of both the rGAPDH forward and reverse primers (sequences as above) in a final volume of 2.5 μL. Once combined with the appropriate base mix, each dilution (total volume 5 μL) was assayed in the TaqMan reaction described below. An optimal working dilution was then determined from these results (data not shown), and both the primer and probe controls were used at a dilution of 10−9.
The TaqMan RT-PCR was performed using the ABI 7500 Fast Real-Time PCR System (PE Applied Biosystems, Foster City, CA). Detection of CHIKV RNA and amplification of the 62 bp product were carried out using a single-tube, one-step RT-PCR format in a final reaction volume of 20 μL. The reaction mix was prepared using the Superscript III Platinum one-step qRT-PCR system (Invitrogen, Carlsbad, CA) and contained 0.4 μL Superscript™ III RT/Platinum® Taq mix, 9.5 μL of 2 × reaction mix, 370 nM primers, 180 nM dual-labeled probe, 47 nM ROX Reference Dye, and 5 μL of extracted viral RNA or diluted synthetic control. The cycling conditions were as recommended by the manufacturer (Fast Mode, PE Applied Biosystems, Foster City, CA) and consisted of one cycle at 50°C for 5 min, one cycle at 95°C for 10 min, and 40 cycles at 95°C for 3 s and 60°C for 30 s. The threshold cycle number (C t) was determined for each sample and a negative result indicating no RNA detection corresponded to any C t value that was ≥40 cycles.
Analysis
The titers of the blood/virus mixtures were estimated using the 50% endpoint method of Reed and Meunch (1938). The virus dose per mosquito was calculated using the assumption that 2 μL of blood/virus solution was ingested by each mosquito (Watson and Kay 1999). The susceptibility of each species to CHIKV infection was calculated by Probit analysis (SPSS 2007). Probit models were assessed using the Pearson chi-square goodness-of-fit statistic, and susceptibility to infection was expressed as ID50 and defined as the virus dose per mosquito at which 50% of the mosquitoes tested positive for CHIKV infection in the CCEIA.
Results
Sensitivity of the CHIKV TaqMan assay
The sensitivity of the CHIKV TaqMan assay was assessed using 10-fold dilutions of RNA extracted from pelleted stock virus, which had an initial titer of 108 CCID50/mL. Results of the titration indicated that at least 0.0112 CCID50/mL could be detected, producing a C t value of 34.63.
Susceptibility to CHIKV infection
To test susceptibility to infection, seven mosquito species were fed CHIKV doses ranging from 100.6 to 104.1 CCID50/mosquito (Fig. 1). All species were susceptible to infection, although Aedes spp. were the most susceptible, with ID50s ranging from 100.6 to 102.6 CCID50/mosquito, for Aedes procax, and Ae. notoscriptus and Ae. aegypti, respectively (Table 2). As infection rates for Culex annulirostris and Culex sitiens exposed to the highest dose were <50%, ID50s were not calculated for these species.
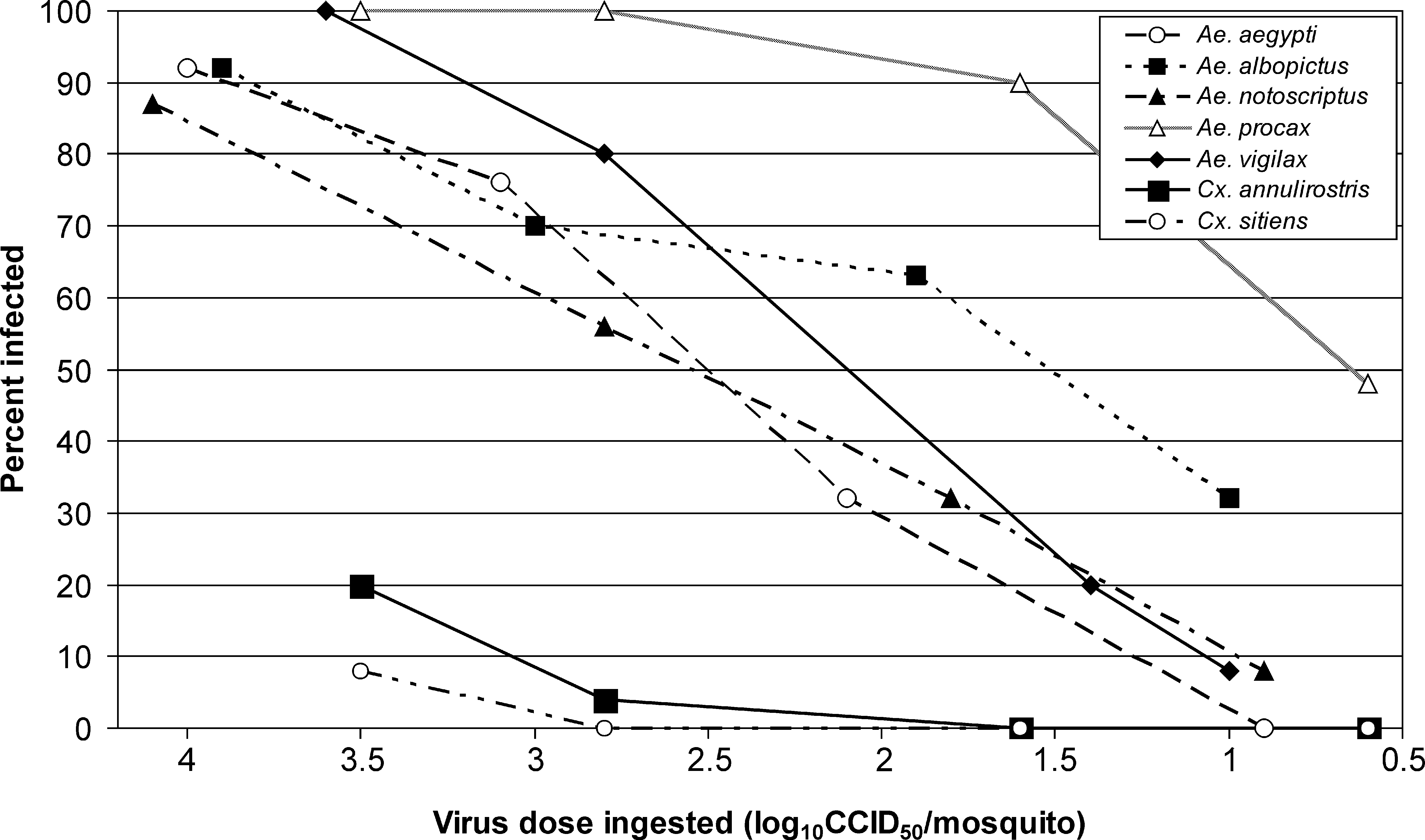
Percent infection rate in seven Australian mosquitoes species exposed to 100.6 to 104.1 cell culture infectious dose (CCID)50/mosquito of chikungunya virus tested at 14–15 days postexposure.
The log10 cell culture infectious dose required to infect 50% of the mosquitoes.
CI, confidence interval.
Infection, dissemination, and transmission
After exposure to titers of 103.0–104.1 CCID50/mosquito of CHIKV, 9 out of the 10 species developed a disseminated infection and transmitted the virus (Table 3). Infection rates in the five Aedes spp. were ≥87%. If the virus infected the midgut, it disseminated through the hemocoel in 100% of Ae. aegypti, Ae. albopictus, Aedes. vigilax, and 96% of Ae. procax. Subsequent transmission rates for these species ranged from 32% for Ae. albopictus to 76% for Ae. vigilax.
Titer (log10CCID50/mosquito) of the infectious blood meal to which mosquitoes were exposed.
Percentage of mosquitoes containing virus in their bodies (number positive/number tested).
Percentage of mosquitoes containing virus in their legs and wings (number positive/number tested).
Percentage of mosquito expectorates in which virus was detected (number of positive expectorates/number tested).
Percentage of mosquitoes with a disseminated infection in which virus was detected in the expectorate (number of positive expectorates/number disseminated).
Of the other species tested, dissemination and transmission rates were <30% and ≤20%, respectively. Only 23% of Ae. notoscriptus developed a disseminated infection, although six out of the seven mosquitoes with a disseminated infection transmitted the virus. Despite an infection rate of 72%, only 28% of Ve. funerea developed a disseminated infection, and 12% transmitted the virus. Infection, dissemination, and transmission rates in Cx. annulirostris and Cx. sitiens were ≤20%, while Cx. quinquefasciatus was refractory to infection.
Discussion
During the emergence of epidemic CHIKV around the Indian Ocean rim, Ae. albopictus and Ae. aegypti have been implicated as the epidemic vectors (Tsetsarkin et al. 2007, Vazeille et al. 2007, Kaur et al. 2008). We have now confirmed that Australian populations of Ae. aegypti and Ae. albopictus are also highly susceptible to infection and readily transmit the virus. The anthropophilic feeding behavior of Ae. aegypti throughout its distribution, including Australia (C.C. Jansen, Australian Biosecurity Cooperative Research Centre for Emerging Infectious Disease, A.F. van den Hurk and S.A. Ritchie, Queensland Health Tropical Public Health Unit, unpublished data), further enhances its status as a major potential vector. Further, Ae. aegypti is a common domestic species, whose high population densities in urban areas in north Queensland have previously facilitated explosive outbreaks of dengue viruses (Hanna et al. 1998, Ritchie et al. 2004). However, as Ae. aegypti currently occurs only in north Queensland, the range through which Ae. aegypti would be considered a major potential CHIKV vector is limited.
Despite a Torres Strait strain of Ae. albopictus being highly susceptible to infection and readily developing a disseminated infection, only 32% of the females transmitted the virus. This was surprising, as other geographical populations of Ae. albopictus have exhibited high vector competence for the E1-A226V strain of CHIKV, and it was this virus–vector combination that led to the explosive nature of the Indian Ocean outbreak in 2006–2007 (Tsetsarkin et al. 2007, Vazeille et al. 2007). Preliminary analysis of Ae. albopictus populations collected from the Torres Strait suggests that there is a considerable genetic diversity in this region (N.W. Beebe, L.A. Hill, University of Queensland and A.F. van den Hurk, unpublished data). While a wild-type population was used to establish the colony, it is plausible that colonizing effects and rapid genetic drift of this small colony population could have reduced the extent of genetic diversity. The resulting colony phenotype that has emerged may display a salivary gland infection and/or escape barrier that was observed in our experiments. Further experiments are required to determine if the different Torres Strait populations of Ae. albopictus differ in their vector competence for CHIKV.
Ae. albopictus is even more restricted in its distribution in Australia than Ae. aegypti, being found only on the Torres Strait islands, situated off northern Queensland. It is feared that if Ae. albopictus becomes established on the mainland, it could rapidly colonize much of Australia, including temperate regions. A number of urban/periurban areas could then become receptive to arboviruses, such as CHIKV and dengue viruses, where currently there is little threat to the human population (Russell et al. 2005). Analysis of the host-feeding patterns of North and South American populations demonstrated that Ae. albopictus is primarily mammalophilic, with human feeding rates ranging between 24% and 50% (Gomes et al. 2003, Richards et al. 2006). However, in both of these studies, when adjusted for geographical and temporal host availability, Ae. albopictus exhibited a predilection for feeding on humans. In Australia, Ae. albopictus was readily sampled using human bait in the Torres Strait, and landing counts on some islands were >50 mosquitoes per 5 min collecting period (Ritchie et al. 2006). Therefore, while only being a moderately efficient vector of CHIKV in our experiments, the ability for Ae. albopictus to rapidly colonize urban/periurban habitats, coupled with its vicious human feeding behavior, suggests that it could play a role in CHIKV transmission should it become established on mainland Australia.
The results of these studies indicate that Australia has several other mosquito species potentially capable of facilitating an outbreak of CHIKV should it be introduced. Aedes spp. that have been implicated previously as endemic RRV and BFV vectors were highly competent laboratory vectors of CHIKV, particularly Ae. vigilax, Ae. procax, and Cq. linealis (Kay 1982, Boyd and Kay 1999, Ryan and Kay 1999, Ryan et al. 2000, Jeffery et al. 2002). This was not unexpected, as RRV, BFV, and CHIKV are antigenically and genetically related viruses in the Alphavirus genus, and are all causative agents of polyarticular disease in humans (Powers et al. 2001). Conversely, the low vector competence of Australian Culex spp. is in agreement with overseas studies on this genus (McIntosh and Jupp 1970, Jupp et al. 1981).
While highly efficient laboratory vectors, Ae. vigilax, Ae. procax, and Cq. linealis possess ecological traits that could influence their status as potential CHIKV vectors. When compared with Ae. aegypti or Ae. albopictus, none of these species utilizes larval habitats closely associated with human habitation. A saltmarsh species, Ae. vigilax is distributed throughout much of coastal Australia, with king tides or above average rainfall triggering large population increases (Lee et al. 1984). Both Ae. procax and Cq. linealis are common freshwater species in southeastern Australia, utilizing transient floodwater pools, and semipermanent water with emergent vegetation as larval habitats, respectively. All three species are opportunistic in their host-feeding habits, with humans generally accounting for <20% of the blood meals identified (Kay et al. 2007, C.C. Jansen, A.F. van den Hurk and S.A. Ritchie, unpublished data). Therefore, the potential role that Ae. procax, Ae. vigilax, and Cq. linealis could play in CHIKV transmission cycles may be offset by their limited contact with humans, especially when compared to Aedes (Stegomyia) spp. Nonetheless, increased urbanization in coastal areas of eastern Australia will further encroach on larval habitats, thus potentially exposing residents to populations of these species, particularly those of Ae. vigilax.
Our results suggest that the Brisbane strain of Ae. notoscriptus expresses a midgut escape barrier to infection, as only 27% of the individuals with an infected midgut developed a disseminated infection. This barrier to transmission may be a trait common with this species and alphaviruses, as it has been previously demonstrated that despite RRV infection rates of >85% in southeast Queensland populations of Ae. notoscriptus, transmission rates are <13% (Watson and Kay 1998, Ryan et al. 2000). However, the lower transmission efficiency of both CHIKV and RRV by Ae. notoscriptus may be countered by the fact that it is the dominant container-inhabiting Aedes spp. associated with human habitation throughout much of Australia (Lee et al. 1982) and it did yield 13 isolates of RRV during an outbreak in 1994 (Ritchie et al. 1997). Indeed, Ae. notoscriptus was the second most abundant species collected using CDC light traps in the western suburbs of Brisbane (Kay et al. 2008). Residents in these suburbs are regularly exposed to bites from this species, as Ae. notoscriptus host seeking coincides with times when residents are often active.
Although evidence of infection in the field is essential for confirming potential vectors, we now have an indication of the species that could be targeted in control programs, should an outbreak of CHIKV occurs in Australia. Additional experiments, employing other mosquito populations, may provide an even more comprehensive assessment of the risk of CHIKV to Australia, including the examination of potential intraspecific variation in vector competence for CHIKV, as has been previously demonstrated for populations of Ae. aegypti and Ae. albopictus abroad (Tesh et al. 1976, Reiskind et al. 2008). Further, the vector competence of Australian mosquito species for other CHIKV strains, particularly those that do not possess the E1-A226V mutation, needs to be assessed. In terms of control strategies, procedures outlined in the Dengue Fever Management Plan for North Queensland aimed at controlling Ae. aegypti could be adapted for the control of other container-inhabiting species, including Ae. albopictus and Ae. notoscriptus (Ritchie et al. 2002). However, as the larval habitats of Ae. vigilax, Ae. procax, and Cq. linealis are more widespread, encompassing temporary or semipermanent ground water sites, alternative strategies may be necessary to suppress transmission.
Footnotes
Acknowledgments
The authors wish to acknowledge Judith Northill and Doris Genge for assisting with virus assays, and Ina Smith and Russell Simmons for useful discussion on sample analysis. We thank Julian Druce for providing the CHIKV isolate and Roy Hall for the provision of the monoclonal antibody. We also thank Petrina Johnson and Luke Rapley for providing the Ae. aegypti eggs and Jay Nicholson for maintaining the Ae. albopictus colony. Nigel Beebe provided valuable insight for the discussion, Jason Jeffery assisted with the Probit analysis, and Cassie Jansen and Jay Nicholson commented on the manuscript. This study was funded by the Australian Biosecurity Cooperative Research Centre for Emerging Infectious Disease (Project number 2.065R).
Disclosure Statement
The authors are not aware of any commercial associations or biases that might be perceived as affecting the objectivity of this research paper.