
Abstract
Dog rabies has commonly been associated with the eastern and southern border areas in Mpumalanga province, and the Nkomazi district in the east has been most affected. In other parts of the province, canid rabies has been under control for many years; however, in 2008, dog rabies spread to other parts of the province and resulted in a widespread outbreak. The objective of this study was to genetically characterize rabies viruses in an attempt to determine the source of this recent outbreak. Fifty-five rabies viruses were recovered from domestic dogs between 2000 and 2008 from Mpumalanga province and bordering areas. The viruses were characterized through nucleotide sequencing of the cytoplasmic domain of the glycoprotein gene and the G-L intergenic region. Phylogenetic analysis of these viruses and those previously characterized from Mpumalanga province and neighboring countries and provinces clearly supported the placement of the viruses from the current outbreak and those from Nkomazi district in one lineage. This demonstrated that the recent emergence of rabies in Mpumalanga province resulted from the spread of rabies from Nkomazi district. A comparative analysis demonstrated close genetic relationships among rabies viruses from Mpumalanga and KwaZulu-Natal provinces, Swaziland, and Mozambique. Findings from this investigation have shown that rabies continues to pose a definite public health threat in South Africa, a situation similar to other African countries.
Introduction
Before 1950, the distribution of rabies in South Africa was limited to the greater part of the interior plateau, west of the Drakensberg mountains, coinciding with part of the distribution of the yellow mongoose (Cynictis penicillata), but this changed markedly following introduction of the canid strain in the 1950s (Swanepoel 2004). Dogs became reservoirs and main vector species of rabies; subsequently, the infection spilled over into wild canids including the black-backed jackal species (Canis mesomelas) and bat-eared foxes (Otocyon megalotis) (Bishop et al. 2002). These two wild carnivore species are capable of sustaining rabies infection cycles in the absence of domestic dogs in the Limpopo province and Cape regions of South Africa (Sabeta et al. 2007, Zulu et al. 2009).
Mpumalanga occupies 6% of the country's total surface area and approximately 7% of the country's population lives in this province. The population is unevenly distributed with approximately half of the population living in communal lands. The distribution of the dog population correlates with that of human settlements, hence large dog populations are found in densely populated communal lands. The initial emergence of canid rabies in this province resulted from an epizootic that originated in Angola in 1947 and spread through Namibia, Botswana, and northern South Africa (the former Transvaal province) in 1950 (Swanepoel 2004). Canid rabies has been endemic in Nkomazi district, which is adjacent to Swaziland and Mozambique, since then (Kloeck 1997, Bishop et al. 2002). Rabies in this area has been self-limiting, largely owing to the geographical landscape and physical barriers (Kloeck 1997). Recently in Mpumalanga province, dog rabies appeared in districts where it had never been reported or had been under control in the past two decades (Onderstepoort Veterinary Institute Records 2008).
Despite the frequent occurrence of black-backed jackal rabies in the Limpopo province, bordering Mpumalanga province in the north (Brückner and Hurter 1978, Barnard 1979, Swanepoel 2004, Zulu et al. 2009), jackals have not played a notable role in the transmission of rabies to the latter (Kloeck 1997). This wild canid is often exterminated in the urban, tribal, forest, and farming areas of this province. The adjacent Kruger National Park (KNP) is reportedly free of rabies (Kloeck 1997), although rabid stray dogs are occasionally found in the park (Skukuza State Veterinary 1977–2006). The rapid spread of the disease in this part of the province became a public and veterinary health concern. The aim of this study, therefore, is to establish the origin of this outbreak by using molecular characterization of rabies virus specimens from the affected area.
Materials and Methods
Viruses
Fifty-five original animal brain specimens recovered from domestic dogs in Mpumalanga, Limpopo, and KwaZulu-Natal provinces, Mozambique, and Swaziland between 2000 and 2008 were included for analysis. Brain samples from rabies-suspect animals were submitted to the Rabies Laboratory at Onderstepoort Veterinary Institute in Pretoria for routine rabies diagnosis. The samples were tested for the presence of lyssavirus antigen using the fluorescent antibody test (Dean et al. 1996), and all those that were positive for RABV antigen were immediately stored at −20°C before molecular characterization. The year of sampling and the locality of origin of specimens are shown in Table 1.
The viruses that were sequenced previously (Coetzee and Nel 2007, Cohen et al. 2007, Zulu et al. 2009).
KNP, Kruger National Park.
Total RNA extraction, reverse transcription (RT)-polymerase chain reaction, and cycle sequencing
Total viral RNA was extracted from the original brain samples, using Trizol reagent according to the manufacturer's instructions (Sigma-Aldrich). The cytoplasmic domain of the glycoprotein gene and the G-L intergenic region of the rabies genome were reverse transcribed and amplified, using the G/L primers as previously described (Sacramento et al. 1991). The polymerase chain reaction amplicons were purified using the Wizard SV Gel Clean-Up System, according to the manufacturer's guidelines (Promega). Purified polymerase chain reaction products were cycle sequenced in both directions with the G/L primer set, using the BigDye Sequencing Kit (ABI PRISM BigDye Terminator v3.1; Applied Biosystems).
Phylogenetic analysis
Nucleotide sequences were edited using Mega 4.1 software and consensus sequences assembled after comparison of the forward and reverse sequences. For phylogenetic analysis, the consensus sequences were trimmed to 592 bp to include the cytoplasmic domain of the glycoprotein and the G-L intergenic region. Nucleotide sequences were then aligned using Clustal X (Higgins and Sharp 1989, Higgins 2003), and a phylogenetic tree was constructed using the neighbor-joining method according to Saitou and Nei (1987). Bootstrap statistical support values were determined using a 1000 replicates to evaluate the significance of the branching pattern, and values of ≥ 70% were regarded as providing evidence for a phylogenetic grouping (Hills and Bull 1993).
Results
Rabies virus specimens recovered from dogs in Limpopo and KwaZulu-Natal provinces, Mozambique, and Swaziland were included to determine the phylogenetic relationships among rabies viruses from these regions with those from Mpumalanga province. Each specimen included in the study was shown to contain RABV antigen by fluorescent antibody test and successfully amplified to yield the expected product of approximately 850 bp in size. Through phylogenetic analysis, viruses characterized in this study segregated into two lineages, A and B (Fig. 1). Lineage A comprised rabies viruses obtained from domestic dogs in Mpumalanga and KwaZulu-Natal provinces, Swaziland, and Mozambique. An average nucleotide sequence homology of 99.7% was obtained from this group of viruses, thus showing that these viruses share a common origin. Lineage B comprised one cluster (LP) consisting of viruses from Limpopo province. These viruses, with a sequence homology of 99.8%, formed a compact cluster that was clearly distinct from that composed of viruses obtained from Mpumalanga and KwaZulu Natal provinces and the neighboring countries. The branch supporting these viruses had a bootstrap support value of 99%.
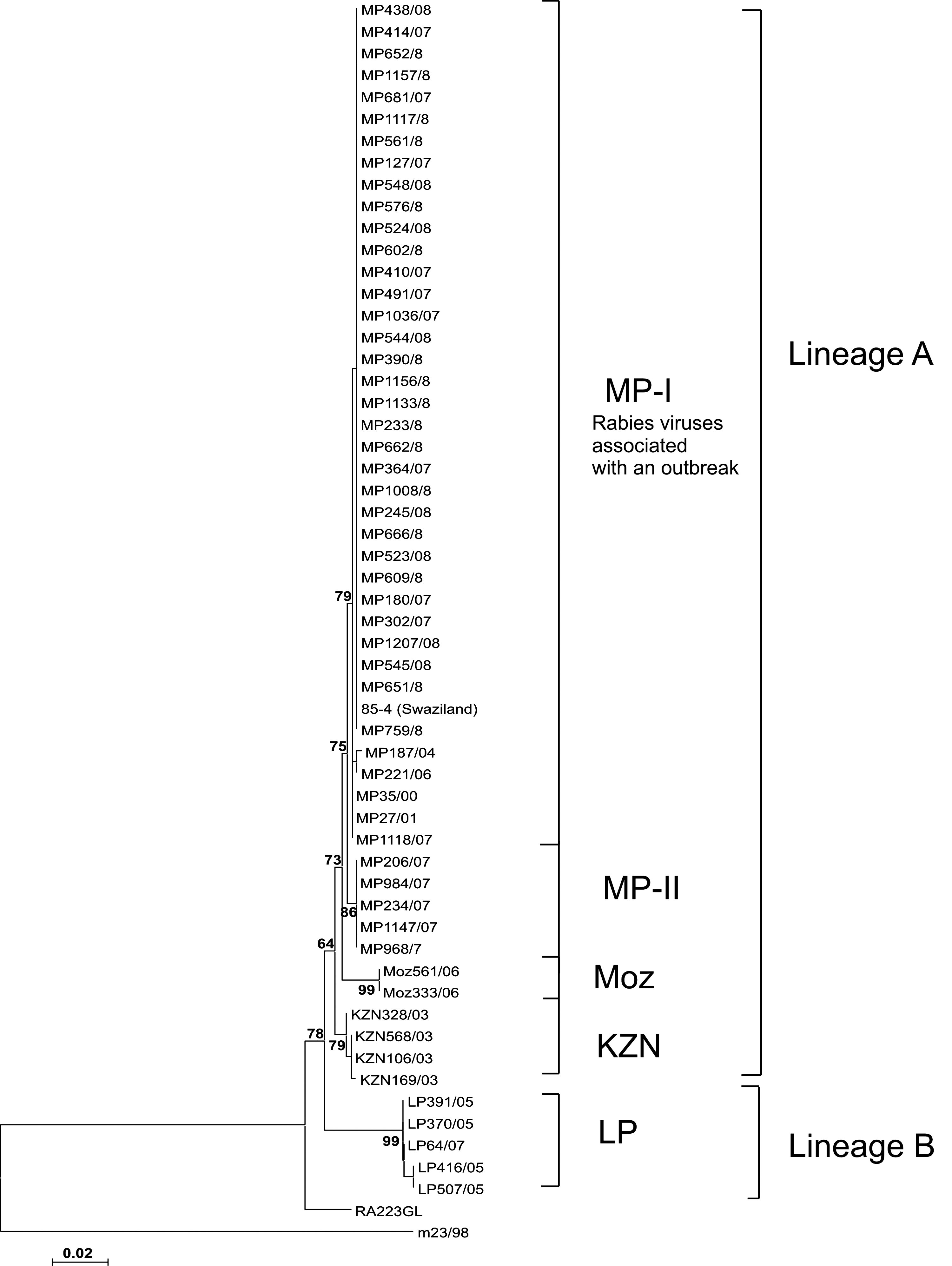
Phylogenetic tree based on an alignment of a 592 bp region of the G-L intergenic region for 55 rabies viruses obtained from domestic dogs from Mpumalanga Province (MP), KwaZulu-Natal Province (KZN), (Lineage A) and Limpopo Province (LP) (Lineage B), Mozambique (Moz), and Swaziland (Swaz). The sequences are preceded by the prefixes LP, MP, KZN, Moz, or Swaz to denote the province or the country of origin. The mongoose rabies virus m23/98 and canid rabies virus RA223GL were used to root the tree.
The branching of the neighbor-joining tree demonstrated that within lineage A four clusters—MP-I, MP-II, Moz, and KZN—could be distinguished (Fig. 1). Although the bootstrap support value of 64% for the branch that gave rise to these clusters is not significant, the internal branches supporting clusters within this lineage have high bootstrap values. Viruses in this lineage are closely related and are likely to have resulted from the spread of a single rabies strain that has a wide distribution in eastern South Africa, Swaziland, and Mozambique. Clustering of viruses was generally determined by their geographic origins demonstrating localized rabies cycles. Two viral clusters—MP-I and MP-II—were identified in Mpumalanga province. All viruses associated with the outbreak grouped together in cluster MP-I. One virus obtained from a dog in Swaziland (85/04) was also found in this cluster. Three viruses (1008/08, 1156/08, and 1157/08) within cluster MP-I were recovered from stray dogs in close proximity to the KNP. Spatial distribution of viruses in cluster MP-II spanned the east and south-eastern border areas of the province. Viruses from Mozambique and KwaZulu-Natal Province were found in separate clusters, Moz and KZN, respectively, according to their origins.
Surveillance data show that in the past decade canid rabies was endemic in Nkomazi district, and frequently cases were also reported from the east and the south-eastern districts (Fig. 2a). In contrast, mongoose rabies had a wide distribution in central and western parts of the province. In early 2008, dog rabies spread from Nkomazi district westward to adjacent Mbombela district and subsequently northward to Bushbuckridge district, including areas surrounding KNP (Fig. 2b).
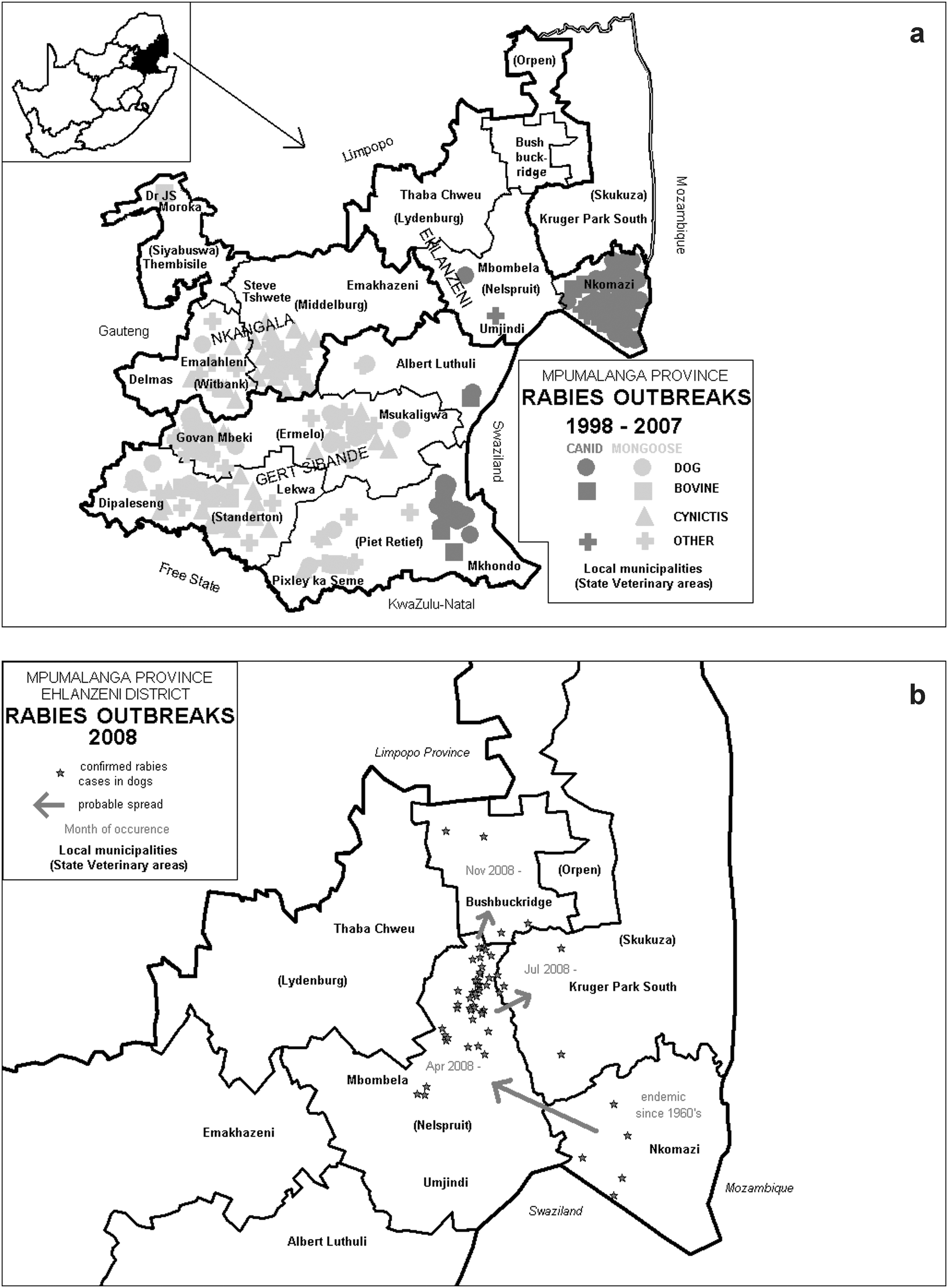
(
Discussion
Genetic characterization of rabies viruses continues to expand our knowledge of global RABV diversity and assist in identifying newly emerging virus-host associations. According to the data presented here, there are currently two rabies cycles maintained by the domestic dog in Mpumalanga province. Overall, these viruses belong to one viral lineage widely distributed in Mpumalanga province, KwaZulu Natal province, Mozambique, and Swaziland. Multiple introduction of the rabies virus into the province could explain the presence of these two cycles. The viruses comprising cluster MP-I were confined to eastern-most Nkomazi district before 2008, and those comprising cluster MP-II were limited to the east and south-eastern border areas. Similar observations were reported from a previous epidemiological study of rabies in northern South Africa (Zulu 2007, Zulu et al. 2009). The areas in Mpumalanga where rabies remains uncontrolled are bordered by Mozambique and Swaziland along the east and KwaZulu Natal province in the south, and all these are also rabies endemic (Dlamini 2001, Dias and Rodrigues 2003, Coetzee and Nel 2007). Given the porous nature of African borders, the exchange of the RABV between vector species from Mpumalanga province and those in the neighboring countries is expected to be fairly frequent. The identification of a virus from Swaziland (d85/4) that was genetically similar to the viruses associated with an outbreak (cluster MP-I) is an example of such transborder transmission of rabies. Globally, transborder transmission is a common feature that facilitates the spread of the disease. The recent spread of rabies from southern Zimbabwe to dogs in Limpopo province in northern South Africa that resulted in a severe human rabies outbreak is one such example (Cohen et al. 2007).
The rabies variant associated with the recent outbreak appears to have been responsible for endemic dog rabies in eastern Mpumalanga. Considering the spatial distribution of the virus, it is possible that human facilitated vector movements resulted in the introduction of rabies into adjacent districts. It is obvious that the affected districts had populations of susceptible dogs that favored the rapid spread of the virus, thereby resulting in the outbreak. The spread of rabies from eastern districts to other parts of the province has demonstrated the inadequacy of the control of the disease through dog vaccination. Likewise, in other rabies endemic provinces in South Africa, the reasons for failure to control rabies include low vaccination coverage and lack of sustainability of vaccination campaigns. Though rabies has never been confirmed in the KNP, the identification of rabies cases in close proximity to the park poses a threat to wildlife. Dog populations in neighboring localities to the park appear as possible sources of rabies infection, a situation similar to that in the Serengeti National Park in Tanzania. In these studies, it was found that the domestic dog populations in communities adjacent to the park were the sources of rabies infection to bat-eared foxes and other wildlife species in the park (Cleaveland and Dye 1995).
The data presented in this report have demonstrated that the dog rabies outbreak resulted from the spread of rabies from Nkomazi district. Emergence of rabies in new geographical zones, in areas where it has been under control for many years, and in new host species has become common in South Africa (Cohen et al. 2007, Sabeta et al. 2007, Ngoepe et al. 2009, Zulu et al. 2009). This development highlights the importance of sustainable and continuous parenteral vaccination of domestic dogs. In addition, strict vigilance and early response to primary outbreaks have to be maintained in endemic areas and buffer zones should be created and maintained in areas bordering such regions. The study further underscores the importance of molecular epidemiology as a tool for identifying sources of infection and tracing routes of spread of infectious agents such as RABV.
Footnotes
Acknowledgments
This work was funded by Department of Agriculture, South Africa (grant # OVI4/16/c171). The authors would like to acknowledge Rosa Costa of the Central Veterinary laboratory in Mozambique for providing rabies samples from Mozambique. We would also like to thank Truuske Gerdes from Onderstepoort Veterinary Institute for critically reviewing this article.
Disclosure Statement
No competing financial interests exist.