
Abstract
Hantavirus HTN·007 was originally isolated from a small-eared pygmy rice rat (Oligoryzomys microtis) captured in northeastern Peru. The results of analyses of nucleotide and amino acid sequence data in this study indicated that HTN·007 is a strain of Rio Mamoré virus (RIOMV) which is enzootic in small-eared pygmy rice rat populations in Bolivia. As such, the results of this study extend our knowledge of the geographical range of RIOMV and support the notion that the small-eared pygmy rice rat is the principal host of RIOMV.
Introduction
Specific rodents are the principal hosts of the hantaviruses for which natural host relationships have been well characterized. For example, the hispid cotton rat (Sigmodon hispidus) in southern Florida is the principal host of Black Creek Canal virus (BCCV) (Ravkov et al. 1995, Rollin et al. 1995, Glass et al. 1998), the marsh rice rat (Oryzomys palustris) in the southeastern United States is the principal host of Bayou virus (BAYV) (Ksiazek et al. 1997), and the long-tailed pygmy rice rat (Oligoryzomys longicaudatus) in Argentina and Chile is the principal host of ANDV (Levis et al. 1998). Other natural hosts of hantaviruses include moles and shrews (order Soricomorpha) (Carey et al. 1971, Arai et al. 2008).
Some hantaviruses cause severe diseases in humans. For example, HTNV is an agent of hemorrhagic fever with renal syndrome in Asia (Lee et al. 1978), SNV is an agent of hantavirus pulmonary syndrome (HPS) in North America (Nichol et al. 1993), and Laguna Negra virus (LANV) and ANDV are agents of HPS in South America (López et al. 1996, Johnson et al. 1997).
The genomes of hantaviruses consist of three single-stranded, negative-sense RNA segments, designated small (S, ∼1.9 kb), medium (M, ∼3.7 kb), and large (L, ∼6.6 kb). These segments encode the nucleocapsid (N) protein, glycoprotein precursor (GPC) for the envelope glycoproteins (Gn and Gc), and RNA-dependent RNA polymerase (RdRp), respectively. Our most comprehensive knowledge of phylogenetic relationships within the genus Hantavirus heretofore was based on analyses of complete N protein gene sequences and complete GPC gene sequences (Ramsden et al. 2009).
RIOMV is naturally associated with the small-eared pygmy rice rat (Oligoryzomys microtis) in Bolivia (Bharadwaj et al. 1997, Carroll et al. 2005). Previously, our knowledge of RIOMV was limited to virus-specific RNA from a small-eared pygmy rice rat (BYU 19014) captured near the town of Mineros in the Department of Santa Cruz (Carroll et al. 2005) and virus-specific RNA from a small-eared pygmy rice rat (OM 604) captured near Rio Madidi in the Department of La Paz, a small-eared pygmy rice rat (OM 556) captured near Rio Beni in the Department of La Paz, and three small-eared pygmy rice rats (OM 137, OM 142, and OM 143) captured near Rio Tijamuchi in the Department of Beni (Bharadwaj et al. 1997) (Fig. 1). We note that RIOMV has never been isolated from a small-eared pygmy rice rat or any other animal captured in Bolivia.
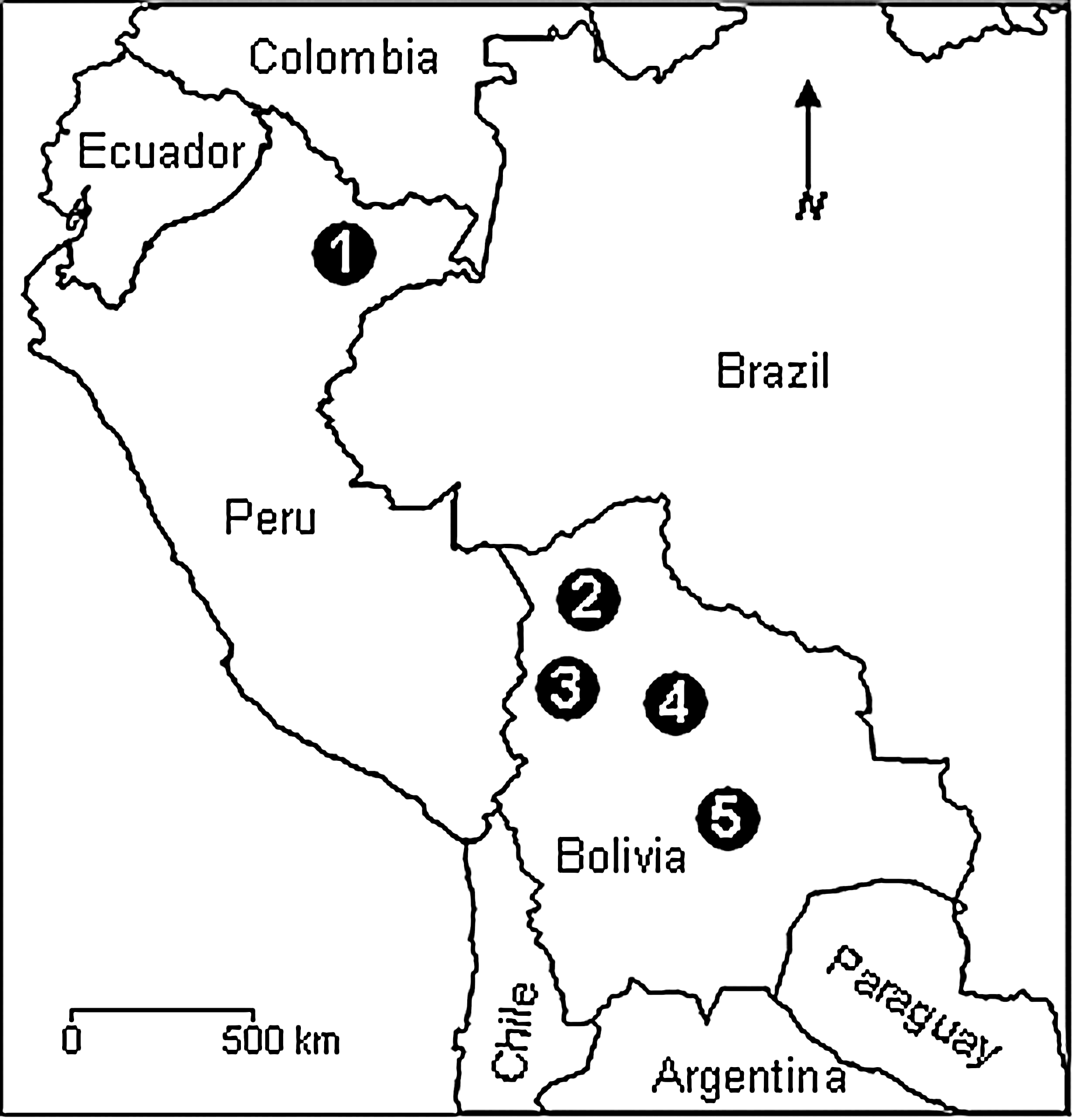
Map showing the five localities at which RIOMV-infected small-eared pygmy rice rats (Oligoryzomys microtis) were captured in previous studies: (1) Iquitos (3°45′S, 73°11′W), (2) Rio Madidi (12°34′S, 67°00′W), (3) Rio Beni (14°16′S, 67°32′W), (4) Rio Tijamuchi (14°56′S, 65°09′W), and (5) Mineros (17°5′S, 63°40′W). RIOMV strains HTN·007 and OM 556 were isolated from small-eared pygmy rice rats captured near Iquitos and Rio Beni, respectively. RIOMV, Rio Mamoré virus.
Hantavirus HTN·007 was originally isolated from a small-eared pygmy rice rat captured near the town of Iquitos in northeastern Peru (Powers et al. 1999) (Fig. 1). The results of an analysis of S segment sequence data in previous studies indicated that HTN·007 is phylogenetically closely related to RIOMV strain OM 556 (Powers et al. 1999, Rosa et al. 2005). The objective of this study was to improve our knowledge of the relationship of HTN·007 to RIOMV strain OM 556 and other members of the genus Hantavirus.
The nucleotide sequences of a 1926-nt fragment of the S segment, 3694-nt fragment of the M segment, and 6554-nt fragment of the L segment of HTN·007 and the nucleotide sequence of a 6561-nt fragment of the L segment of MAPV strain HV 97021050 were determined in this study. These sequences included the complete N protein gene, complete GPC gene, and complete RdRp gene of HTN·007 and the complete RdRp gene of MAPV strain HV 97021050, respectively. The sequence of the L segment of MAPV strain HV 97021050 was determined to improve the representation of the South American hantaviruses in the phylogenetic analysis of RdRp gene sequence data.
Materials and Methods
Genetic characterization of HTN·007 and the L segment of MAPV strain HV 97021050
Total RNA was isolated from monolayers of Vero E6 cells infected with either HTN·007 or HV 97021050, using TRI REAGENT™ (Sigma, St. Louis, MO). First-strand cDNA was generated from the S, M, and L segments of HTN·007 and the L segment of HV 97021050 by using SuperScript II RNAse H- reverse transcriptase (Invitrogen Life Technologies, Carlsbad, CA) in conjunction with oligonucleotides HTS90 (5′-TAGTAGTAGACTCCTTGAGAAGCTA-3′), HTM91 (5′-TAGTAGTAGACTCCGCAAGAAGAAGC-3′), HTL59 (5′-TAGTAGTAGACTCCGGGATAGAAAAGATCAGAATAATGG-3′), and HTL1 (5′-GTAGTAGACTCCGGGATAGAAA-3′), respectively. HTS90 was expected to anneal to the 25-nt fragment at the extreme 3′ end of the S segment of HTN·007, HTM91 to the 26-nt fragment at the extreme 3′ end of the M segment of HTN·007, HTL59 to the 39-nt fragment at the extreme 3′ end of the L segment of HTN·007, and HTL1 to a 22-nt fragment near the 3′ end of the L segment of HV 97021050. Amplicons were generated from the first-strand cDNA by using the Triple Master® PCR System (Eppendorf North America, Westbury, NY) in conjunction with HTN·007 S segment (HTS92 [5′-TAGTAGTATGCTCCTTGAAAAGCATTC-3′] and HTS90), HTN·007 M segment (HTM101 [5′-TAGTAGTATGCTCCGCAGGAACAAAAGCCTCGGTAAAAGGATATAG-3′] and HTM91), HTN·007 L segment (HTL60 [5′-TAGTAGTATGCTCCGGGAAAAGAACAC-3′] and HTL61 [5′-ATGTATGTRAGTGCWGATGCCAC-3′], and HTL91 [5′-ACATACCTAATCACCATCTCTACG-3′] and HTL59), and HV 97021050 L segment (HTL19 [5′-TAGTAGTATGCTCCGRGAAMAGA-3′] and HTL8 [5′-GAACTGTATCCTGAGCTGGAATG-3′], HTL3 [5′-CTCCATAATAGACATTGAGCC-3′] and HTL4 [5′-TATGTKAGTGCTGATGCCAC-3′], and HTL9 [5′-CCTGGAAGGTCAGACAGTGAACC-3′] and HTL1). Both strands of each gel-purified amplicon were sequenced directly, using the Big Dye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA). The nucleotide sequences of the S, M, and L segments of HTN·007 and the nucleotide sequence of the L segment of HV 97021050 were deposited into the GenBank nucleotide sequence database under accession nos. FJ532244, FJ608550, FJ809772, and EU788002, respectively.
Genetic characterization of rodent TTU76249
HTN·007 was originally isolated from pygmy rice rat TTU76249 (Museum of Texas Tech University). The nucleotide sequence of the cytochrome-b (Cytb) gene of TTU76249 was determined in this study to affirm the species identity of this rodent and define the phylogenetic relationship between the pygmy rice rat associated with HTN·007 in Peru and small-eared pygmy rice rat BYU 19014 (GenBank accession no. AY439000), which was captured in Bolivia and naturally infected with RIOMV. Genomic DNA was isolated from 0.1 g of liver, using a phenol extraction protocol (Longmire et al. 1997). The complete Cytb gene (1143-bp) was amplified using Go-Taq ® DNA Polymerase (Promega, Madison, WI) in conjunction with oligonucleotides L14115 and H15288 (Martin et al. 2000). Both strands of the amplicon were sequenced directly, using the Big Dye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems), and the nucleotide sequence of the Cytb gene of TTU76249 was deposited into the GenBank nucleotide sequence database under accession no. FJ374766.
Data analysis
The analyses of the N protein, GPC, and RdRp gene sequences included 25, 22, and 11 viruses, respectively (Table 1). The predicted amino acid sequences were aligned using the computer program CLUSTALW (2.09) (Thompson et al. 1994), nonidentities between amino acid sequences were calculated using the computer program MEGA, version 4.0 (Tamura et al. 2007), and the nucleotide sequences of the N protein genes, GPC genes, and RdRp genes were aligned manually, guided by the computer-generated amino acid sequence alignments.
Apodemus agrarius, striped field mouse; Apodemus flavicollis, yellow-necked mouse; Bandicota indica, greater bandicoot rat; Calomys laucha, small vesper mouse; Lemmus sibiricus, Siberian brown lemming; Microtus arvalis, common vole; Microtus californicus, California vole; Microtus fortis, reed vole; Microtus pennsylvanicus, meadow vole; Myodes glareolus, bank vole; Oligoryzomys fulvescens, fulvous pygmy rice rat; Oligoryzomys longicaudatus, long-tailed pygmy rice rat; Oligoryzomys microtis, small-eared pygmy rice rat; Oryzomys couesi, Coues' rice rat; Oryzomys palustris, marsh rice rat; Peromyscus leucopus, white-footed mouse; Peromyscus maniculatus, deer mouse; Rattus norvegicus, brown rat; Reithrodontomys megalotis, western harvest mouse; Reithrodontomys mexicanus, Mexican harvest mouse; Sigmodon alstoni, Alston's cotton rat; Sigmodon hispidus, hispid cotton rat; Suncus murinus, Asian house shrew.
Russia (KK), Khabarovsk Krai; Russia (SI), Siberia; USA (CA), California; USA (FL), Florida; USA (LA), Louisiana; USA (MD), Maryland; USA (NM), New Mexico; USA (NY), New York; USA (TX), Texas.
RdRp, RNA-dependent RNA polymerase; GPC, glycoprotein precursor; N protein, nucleocapsid protein; na, nucleotide sequence oí the complete RdRp, GPC, or N protein gene not available from the GenBank nucleotide sequence database (June 1, 2009).
The marsh rice rat (Oryzomys palustris) is the principal host of BAYV; however, the nucleotide sequences of BAYV strain Hu_La are from a human (specifically, a fatal case of hantavirus pulmonary syndrome).
Catacamas virus, Choclo virus, and Maporal virus are provisional species in the genus Hantavirus.
Myodes glareolus formerly was Clethrionomys glareolus.
ANDV, Andes virus; BAYV, Bayou virus; BCCV, Black Creek Canal virus; CADV, Caño Delgadito virus; CATV, Catacamas virus; CHOV, Choclo virus; DOBV, Dobrava-Belgrade virus; ELMCV, El Moro Canyon virus; HTNV, Hantaan virus; ISLAV, Isla Vista virus; KHAV, Khabarovsk virus; LANV, Laguna Negra virus; MAPV, Maporal virus; MULV, Muleshoe virus; NYV, New York virus; PHV, Prospect Hill virus; PUUV, Puumala virus; RIOMV, Rio Mamoré virus; RIOSV, Rio Segundo virus; SEOV, Seoul virus; SNV, Sin Nombre virus; THAIV, Thailand virus; TPMV, Thottapalayam virus; TOPV, Topografov virus; TULV, Tula virus.
The analyses of Cytb gene sequences included small-eared pygmy rice rat (O. microtis) BYU 19014 (Department of Santa Cruz in Bolivia; GenBank accession no. AY439000), small-eared pygmy rice rats MVZ193785 and MVZ193858 (western Brazil, U58381 and EU258549, respectively), Andean pygmy rice rat (Oligoryzomys andinus) NK11547 (western Bolivia, AY452200), Chacoan pygmy rice rat (Oligoryzomys chacoensis) TTU104514 (northwestern Paraguay, EU258453), destructive pygmy rice rat (Oligoryzomys destructor) ACUNHC898 (Ecuador, EU258544), yellow pygmy rice rats (Oligoryzomys flavescens) MNHNP3357 (southwestern Paraguay, EU258542), GD010 (southwestern Paraguay, AY452199), and CRB1405 (eastern Brazil, EU258545), fulvous pygmy rice rats (Oligoryzomys fulvescens) TTU84609 (Honduras, EU258547) and TTU104513 (southern Mexico, EU258548), long-tailed pygmy rice rat (O. longicaudatus) MVZ155842 (southern Argentina, U03535), Moojen's pygmy rice rat (Oligoryzomys moojeni) MN36357 (northeastern Brazil, DQ826017), black-footed pygmy rice rats (Oligoryzomys nigripes) TTU104515 (southeastern Paraguay, EU258550), TK61708 (southwestern Paraguay, EU258551), and NK22527 (northeastern Paraguay, AY041196), sprightly pygmy rice rats (Oligoryzomys vegetus) ROM113156 (Costa Rica, EU258541) and ROM112193 (Nicaragua, EU258538), pygmy rice rats (Oligoryzomys species) FHV·4083 (western Venezuela, DQ227457) and ACUNHC275 (southwestern Venezuela, EU258537), minute spiny mouse (Neacomys minutus) AMNH272867 (Peru, EU258536), forest small rice rat (Microryzomys minutus) MVZ166666 (Peru, EU258535), and Peruvian rice rat (Oreoryzomys balneator) ACUNHC1204 (Ecuador, EU258534). Kimura 2-parameter mean pairwise distances between the nucleotide sequences were calculated using MEGA, version 4.0 (Tamura et al. 2007).
Phylogenetic relationships among the viruses in each data set and among the rodents were estimated using a Bayesian Markov Chain Monte Carlo (MCMC) method implemented in the computer program MrBayes 3.1.2 (Huelsenbeck and Ronquist 2001, Ronquist and Huelsenbeck 2003). Each analysis utilized the general time reversible + I + G model of nucleotide substitution, as determined by MrModeltest, version 2 (Nylander 2004). Two independent runs of four Markov chains (10 million generations each run) were performed for each data set, with sampling every 1000th generation. The first 1000 trees were discarded after review of the likelihood scores, the consensus tree (50% majority rule) was constructed from the remaining trees, and clades with probability values of >0.95 were considered well supported by the data. The Thottapalayam virus sequences were the designated out group taxa in the analyses of the N protein, GPC, and RdRp gene sequence data. Similarly, the minute spiny mouse AMNH272867, forest small rice rat MVZ166666, and Peruvian rice rat ACUNHC1204 were the designated out group taxa in the analyses of Cytb data.
Results
The alignment of complete N protein gene sequences (n = 25) was 1335 characters in length. The Bayesian analysis of the N protein gene sequences (Fig. 2A) placed HTN·007 in a close sister relationship to RIOMV strain OM 556 and placed HTN·007 and RIOMV strain OM 556 in a sister relationship to LANV. Further, the Bayesian analysis of the N protein gene sequences grouped HTN·007, RIOMV strain OM 556, LANV, ANDV, CHOV, and MAPV with Caño Delgadito virus (CADV) and grouped BAYV and CATV with BCCV and Muleshoe virus (MULV).
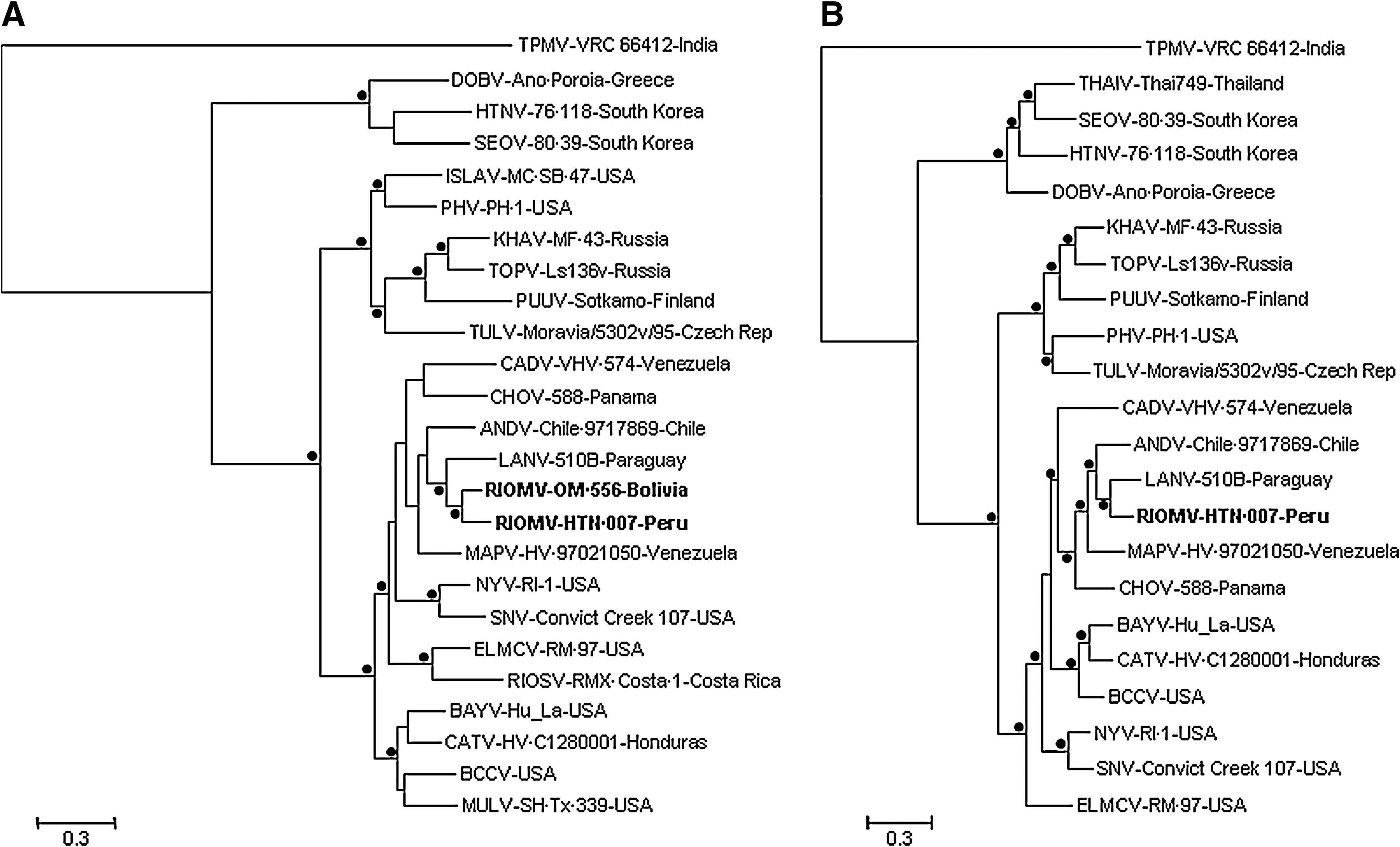
Phylogenetic relationships among hantaviruses based on Bayesian analyses of (
The alignment of complete GPC gene sequences (n = 22) was 3468 characters in length. (RIOMV strain OM 556, Rio Segundo virus, and MULV were not included in the alignment because the nucleotide sequences of the complete GPC genes of these viruses have not been determined.) The Bayesian analysis of the GPC gene sequences (Fig. 2B) placed HTN·007 in a sister relationship to LANV, grouped HTN·007, LANV, ANDV, CHOV, and MAPV with CADV, and grouped BAYV and CATV with BCCV.
The alignment of complete RdRp gene sequences (n = 11) was 6480 characters in length. The Bayesian analysis of the RdRp gene sequences (Bayesian tree not shown) indicated that ANDV, MAPV, and HTN·007 are monophyletic and separated the ANDV–MAPV–HTN·007 lineage from SNV, the only North American virus included in the analysis of RdRp gene sequences.
Nonidentity between the complete sequences of the N proteins of HTN·007 and RIOMV strain OM 556 was 2.1%, nonidentities between the complete sequences of the N proteins of HTN·007 and OM 556 and the complete sequences of the N proteins of LANV strain 510B and ANDV strain Chile-9717869 ranged from 6.8% to 8.9%, and nonidentity between the complete sequences of the N proteins of 510B and Chile-9717869 was 9.6% (Table 2).
Sequence nonidentities between the glycoprotein precursors and between the nucleocapsid proteins are listed above and below the diagonal, respectively.
Strain OM 556 was not included in the pairwise comparisons of complete GPC sequences because the primary structure of the complete GPC of this virus has not been determined; however, nonidentity between the sequence of a 164-aa fragment of the GPC of strain OM 556 and the sequence of the homologous region of the GPC of HTN·007 was 1.2% (see Results section).
Our knowledge of the amino acid sequence of the GPC of RIOMV in Bolivia presently is limited to a 164-aa fragment of the GPC of strains OM 556 and OM 137 (GenBank accession nos. U73688 and U73687, respectively). Nonidentities between these sequences and the sequence of the homologous region of the GPC of HTN·007 ranged from 1.2% to 2.4%. Similarly, nonidentities between the GPC sequences of strains OM 556 and OM 137 and the sequence of the homologous region of the GPC of LANV strain 510B ranged from 4.9% to 6.1%. In pairwise comparisons of complete GPC sequences, nonidentity between the GPC of HTN·007 and 510B was 8.8% and nonidentity between the GPC of 510B and ANDV strain Chile-9717869 was 13.5% (Table 2).
The Bayesian analysis of the Cytb gene sequences (Fig. 3) indicated that TTU76249 (Peru) and small-eared pygmy rice rats MVZ193858 (western Brazil), MVZ193785 (western Brazil), and BYU 19014 (Bolivia) are monophyletic and phylogenetically distinct from the other members of the genus Oligoryzomys included in the analysis. In pairwise comparisons of Cytb gene sequences, genetic distances between TTU76249, MVZ193785, MVZ193858, and BYU 19014 ranged from 1.5% to 3.3%, genetic distances between phylogenetically closely related species (e.g., O. fulvescens and O. vegetus) were greater than 8.1%, and genetic distances between TTU76249, MVZ193785, MVZ193858, and BYU 19014 and the 17 other pygmy rice rats included in the analysis ranged from 11.3% to 14.4%. These results confirm that TTU76249 is a small-eared pygmy rice rat and indicate that TTU76249 is phylogenetically closely related to the small-eared pygmy rice rat associated with RIOMV in Bolivia.
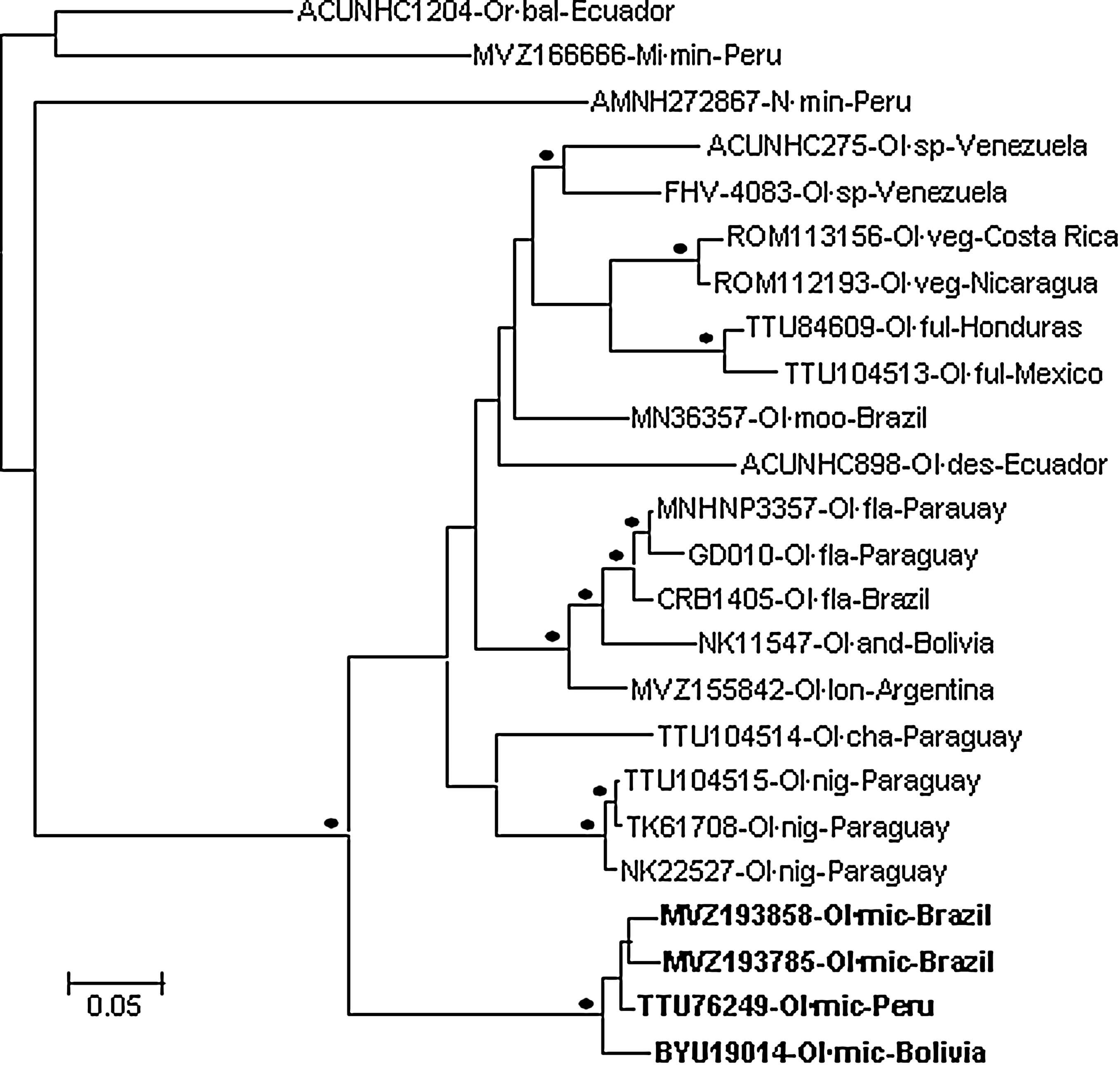
Phylogenetic relationships among rodents based on Bayesian analysis of cytochrome b gene sequences. The scale bar indicates the expected number of substitutions per site. The black dots (•) at the nodes indicate clade probability values of >95.0%. The branch labels include (in the following order) identification number of the voucher specimen, species, and country. Mi·min, Microryzomys minutus (forest small rice rat); N·min, Neacomys minutus (Jurua bristly mouse); Ol·and, Oligoryzomys andinus (Andean pygmy rice rat); Ol·cha, Oligoryzomys chacoensis (Chacoan pygmy rice rat); Ol·des, Oligoryzomys destructor (destructive pygmy rice rat); Ol·fla, Oligoryzomys flavescens (yellow pygmy rice rat); Ol·ful, Oligoryzomys fulvescens (fulvous pygmy rice rat); Ol·lon, Oligoryzomys longicaudatus (long-tailed pygmy rice rat); Ol·mic, O. microtis (small-eared pygmy rice rat); Ol·moo, Oligoryzomys moojeni (Moojen's pygmy rice rat); Ol·nig, Oligoryzomys nigripes (black-footed pygmy rice rat); Ol·sp, Oligoryzomys species (pygmy rice rats not identified to species level); Ol·veg, Oligoryzomys vegetus (sprightly pygmy rice rat); Or·bal, Oreoryzomys balneator (Peruvian rice rat).
Discussion
The Eighth Report of the International Committee on Taxonomy of Viruses (Nichol et al. 2005) set forth four criteria for demarcation of a hantaviral species. The virus must (1) exhibit at least a 7% difference in amino acid sequence identity from strains of other hantaviral species in comparisons of complete N protein sequences and comparisons of complete GPC sequences, (2) occupy a unique ecological niche, (3) represent a unique serotype (as defined by two-way cross-neutralization tests), and (4) not be a product of genetic reassortment between strains of different hantaviral species.
Collectively, the monophyly of HTN·007 and RIOMV strain OM 556 in the Bayesian tree generated from N protein gene sequence data, the high level of identity between the complete sequences of the N proteins of HTN·007 and RIOMV strain OM 556, and the high level of identity between the sequence of the GPC of HTN·007 and sequences of the GPC of RIOMV strains OM 556 and OM 137 indicate that HTN·007 is a strain of RIOMV. As such, the isolation of HTN·007 from rodent TTU76249 is evidence that the geographical range of RIOMV includes northern Peru and supports the notion that the small-eared pygmy rice rat is the principal host of RIOMV.
The results of the Bayesian analyses of N protein and GPC gene sequences indicate that HTN·007 is phylogenetically more closely related to LANV strain 510B than to any of the other viruses included in this study. We note that the International Committee on Taxonomy of Viruses previously determined that RIOMV and LANV are different species in the genus Hantavirus (Nichol et al. 2005). We also note that LANV is naturally associated with the small vesper mouse (Calomys laucha) in western Paraguay (Johnson et al. 1997) and large vesper mouse (Calomys callosus) in northwestern Argentina (Levis et al. 2004) and that the serological relationship between RIOMV and LANV in two-way cross-neutralization tests has not been determined. (We were unable to obtain an isolate of LANV strain 510B for LANV × RIOMV cross-neutralization tests in this study.) Regardless, the 7.2% nonidentity between the complete sequences of the N proteins of HTN·007 and 510B, 8.8% nonidentity between the complete sequences of the GPC of HTN·007 and 510B, and association of RIOMV with the small-eared pygmy rice rat in Peru support the treatment of RIOMV as a species distinct from LANV.
Cotton rats (Sigmodon spp.), rice rats (Oryzomys spp.), and pygmy rice rats (Oligoryzomys spp.) are members of the family Cricetidae, subfamily Sigmodontinae (Musser and Carleton 2005). Alston's cotton rat (S. hispidus) in western Venezuela is the principal host of CADV (Fulhorst et al. 1997), the hispid cotton rat in northern Texas is the putative principal host of MULV (Rawlings et al. 1996), Coues' rice rat (Oryzomys couesi) in Honduras is the principal host of CATV (Milazzo et al. 2006), the fulvous pygmy rice rat (O. fulvescens) in Panama is the principal host of CHOV (Vincent et al. 2000), a pygmy rice rat (Oligoryzomys spp.) in western Venezuela is the principal host of MAPV (Fulhorst et al. 2004), and as indicated previously, the hispid cotton rat in southern Florida, marsh rice rat in the southeastern United States, and long-tailed pygmy rice rat in Argentina and Chile are the principal hosts of BCCV, BAYV, and ANDV, respectively. The present-day association of phylogenetically closely related hantaviruses with congeneric, allopatric, sigmodontine rodents (e.g., BAYV with the marsh rice rat in the southeastern United States and CATV with Coues' rice rat in Honduras) suggests that the principal host relationships of some hantaviruses are a consequence of an ancient relationship between the genus Hantavirus and subfamily Sigmodontinae.
The geographical range of the small-eared pygmy rice rat includes the Amazon Basin of Brazil and contiguous lowlands of Peru, Bolivia, and Paraguay (Musser and Carleton 2005). Assuming that the present-day association of RIOMV with the small-eared pygmy rice rat represents an ancient relationship, the geographical range of RIOMV may include Brazil and Paraguay as well as Peru and Bolivia.
The results of the Bayesian analyses of N protein gene and GPC gene sequences in this study indicate that the hantaviruses principally associated with rice rats are phylogenetically more closely related to BCCV than to the hantaviruses principally associated with pygmy rice rats and that the hantaviruses principally associated with pygmy rice rats are phylogenetically more closely related to CADV than to the hantaviruses principally associated with rice rats. We note that cotton rats are the principal hosts of BCCV and CADV. Hypothetically, BAYV, RIOMV, and the other hantaviruses associated with oryzomyine rodents (Oryzomys spp. and Oligoryzomys spp.) descended from viruses that were originally associated with cotton rats and the association between the genus Hantavirus and oryzomyine rodents was established after the last common ancestor of BCCV and CADV.
As stated previously, the results of the Bayesian analyses in this study indicate that RIOMV is phylogenetically more closely related to LANV than ANDV. The association of LANV with the small vesper mouse in Paraguay and large vesper mouse in Argentina may be a consequence of interspecific (pygmy rice rat-to-vesper mouse) virus transmission after the divergence of RIOMV from ANDV rather than an example of the long-term shared evolutionary history of hantaviruses and rodents in the subfamily Sigmodontinae.
The published literature on autochthonous HPS in Peru and Bolivia is limited to a report of a nonfatal case and fatal case in the Department of Santa Cruz in Bolivia (Carroll et al. 2005). The etiological agent in the nonfatal case was LANV; the identity of the etiological agent in the fatal case was not determined. The present lack of evidence for HPS caused by RIOMV should be interpreted with caution because fatal HPS in eastern Brazil has been linked to a hantavirus that is phylogenetically more closely related to RIOMV than LANV and naturally associated with Fornes' pygmy rice rat (Oligoryzomys fornesi) (Rosa et al. 2005). In pairwise comparisons, the amino acid sequence of a 301-aa fragment of the N protein of the hantavirus associated with Fornes' pygmy rice rat (GenBank accession no. DQ451829) was 95.7% and 96.7% identical to the homologous sequences of the N proteins of RIOMV strains OM 556 and HTN·007, respectively.
Footnotes
Acknowledgments
John D. Hanson and Martin H. Richter contributed equally to this study. National Institutes of Health grant AI-067947 (Viral determinants of hantavirus pulmonary disease in the hamster) provided the financial support for this study.
Disclosure Statement
No competing financial interests exist.