
Abstract
Ectoparasites, including chigger mites (genera Leptotrombidium, Schoengastia, and Blankarrtia) and one tick (genus Haemaphysalis) collected from wild-caught rodents in Thailand, were assessed for the presence of Bartonella DNA by using a polymerase chain reaction assay targeting the 16S–23S intergenic spacer region and citrate synthase gene (gltA). Of the 41 pooled samples tested, 34 were positive for Bartonella DNA. Sequence analysis demonstrated that DNA detected in 33 chigger mite pools and one tick pool was similar to Bartonella tamiae sequences previously isolated from three patients in Thailand. This is the first report of the detection of B. tamiae DNA in chigger mites; additional field and experimental investigations are required to determine the role of chigger mites as potential vectors of B. tamiae.
Introduction
Bloodsucking arthropods have been shown to be involved in the transmission of Bartonella species among their hosts (Billeter et al. 2008a). Sandflies (Lutzomyia spp.), the human body louse (Pediculus humanus humanus), and the cat flea (Ctenocephalides felis) have been implicated in the transmission of B. bacilliformis, B. quintana, and B. henselae, respectively (Chomel et al. 1996, Raoult and Roux 1999, Karem et al. 2000). Ticks were suspected to be involved in the transmissions of B. vinsonii subsp. berkhoffii and B. vinsonii subsp. arupensis (Pappalardo et al. 1997, Welch et al. 1999, Chang et al. 2001), and recent experimental study suggests that Ixodes ricinus may be involved in the transmission of B. henselae (Cotté et al. 2008). The DNA of Bartonella species has been detected in fleas collected from diverse wild rodents (Stevenson et al. 2003, Marié et al. 2006, Morway et al. 2008). In addition, Bartonella DNA was found in keds, biting flies, mites, and miscellaneous arthropods (Billeter et al. 2008a). These results demonstrated the potential role of these various insects as vectors for the transmission of Bartonella sp. among mammalian hosts.
Kosoy and colleagues (2008) recently isolated a novel Bartonella sp., B. tamiae, from three patients in Thailand. The patients showed symptoms typical for bartonellosis, such as fever, mild anemia, and ocular disorder. Although these patients had reported contact with rats, the source and/or possible vectors of the infection remains unknown. Recent detection of B. tamiae-like DNA in Amblyomma americanum ticks (Acari: Ixodidae) collected in Virginia also suggests a potential role of ticks in the transmission of B. tamiae (Billeter et al. 2008).
More than 250 species of mites are associated with health-related problems for humans and domestic animals (Mullen and Durden 2002). Probably foremost among these problems are temporary irritation of the skin, dermatitis, and allergies. There are very few human diseases caused by pathogens transmitted by mites, with two significant exceptions: rickettsialpox and scrub typhus (Mullen and Durden 2002). Several studies have implicated mites of the superfamily Dermanyssoidea in the transmission of bacterial pathogens, such as Salmonella, Pasteurella, and Borrelia (Netusil et al. 2005, Valiente et al. 2005). Orientia tsutsugamushi, the etiological agent of scrub typhus, is the only known pathogen transmitted by trombiculid mites (genus Trombicula) (Lerdthusnee et al. 2003).
The aim of this study was to evaluate mites and other ectoparasites for the presence of Bartonella-specific DNA.
Materials and Methods
Ectoparasites were collected from 41 rodents of five species: Rattus rattus (29), Rattus argentiventer (2), Bandicota indica (5), Bandicota savilei (3), and Mus cervicolor (2). These rodents were captured from five different regions of Thailand: the northern region (Nam province, n = 7), the northeastern region (Ubon Ratchathani and Ubon Thani provinces, n = 8), the eastern region (Chon Buri province, n = 1), the central region (Nonthaburi and Pha Nakon Si Ayutthaya provinces, n = 3), and the southern region (Nakhon Si Thammarat, Surat Thani, and Yala provinces, n = 22).
The ectoparasites were morphologically identified to genus. The 209 collected mites belonged to the genera Leptotrombidium (130), Schoengastia (67), and Blankarrtia (12), and eight collected ticks belonged to the genus Haemaphysalis. The mites collected from each animal were pooled for testing by mite genus. The number of mites in each pool was from 1 to 9 (mean, 5.3). The eight ticks were collected from one mouse and were also pooled together. In total, we tested 40 pools of 209 mites and one pool of eight ticks. The 40 tested mite pools belonged to the genera Leptotrombidium (24), Schoengastia (14), and Blankarrtia (2). DNA was extracted using the Qiagen Mini Kit (Qiagen, Valencia, CA) following the manufacturer's protocol for blood and body fluid, with minor modifications. Briefly, polymerase chain reaction (PCR) was performed in 25 μL of mixtures containing 12.5 μL of 2 × iQ SYBR Green Supermix (BioRad, Hercules, CA), 0.5 pmol of each primer, and 2.5 μL of template DNA. Bartonella DNA was amplified using primers for intergenic spacer (ITS) (Billeter et al. 2008b) and for gltA specifically designed for detection of B. tamiae (BtGLT5′: TTC CTG AGT TTG TAG CAA AA; BtGLT3′: GGA TCA TCT TTA ATG CCC AA). The amplification of DNA was performed on a thermal cycler (iCycler; BioRad). Briefly, for ITS, 1 cycle for 3 min at 95°C was followed by 55 cycles for 30 s at 95°C, 30 s at 57°C, and 30 s at 72°C, and a final extension cycle for 7 min at 72°C; and for gltA, 1 cycle for 4 min at 94°C, followed by 45 cycles for 30 s at 95°C, 60 s at 49°C, and 30 s at 72°C, and a final extension cycle for 7 min at 72°C. Ten microliters of each PCR product was run on a 1.5% agarose gel (Agarose; Promega, Madison, WI).
Positive PCR products were sequenced using an Applied Biosystems Model 3130 Genetic Analyzer (Applied Biosystems, Foster City, CA). The CLUSTAL-X program was used for the phylogenetic analysis of the obtained sequences. The neighbor-joining method by Kimura's two-parameter distance method and bootstrap calculation was carried out for 1000 resamplings. MEGA 4.0.1 software (The Biodesign Institute, Tempe, AZ) was used for these analyses.
Results
Of the 40 pooled mite samples, 29 (72.5%) and 9 (22.5%) were positive for B. tamiae based on ITS and gltA, respectively (Table 1). Among these, five samples were positive for both genes (12.5%). One tick sample was also positive for B. tamiae for ITS. The length of the PCR amplicons for ITS and gltA were 230–242 and 114–116 bp, respectively.
This includes a tick pool.
Sequence analysis of the ITS amplicons revealed two distinct sequence groups, one of which formed a cluster with B. tamiae strain Th239 (ITS239) and the other with strains Th307 and Th339 (ITS307/339), which were previously isolated from humans (Kosoy et al. 2008) (Fig. 1). Fourteen of the 30 ITS sequences (34.2% of tested) grouped with the ITS239 sequence, whereas the other 16 (39.0%) grouped with the ITS307/339 sequence (Table 2). These groups can be distinguished by the presence of 12-bp insertion in the amplified ITS region; this insertion was present only in ITS239 but not in ITS307/339 (data not shown). Analysis of the gltA sequences also revealed two distinct groups, and both groups were closer to strains Th239/307 than to Th339 (Fig. 2).
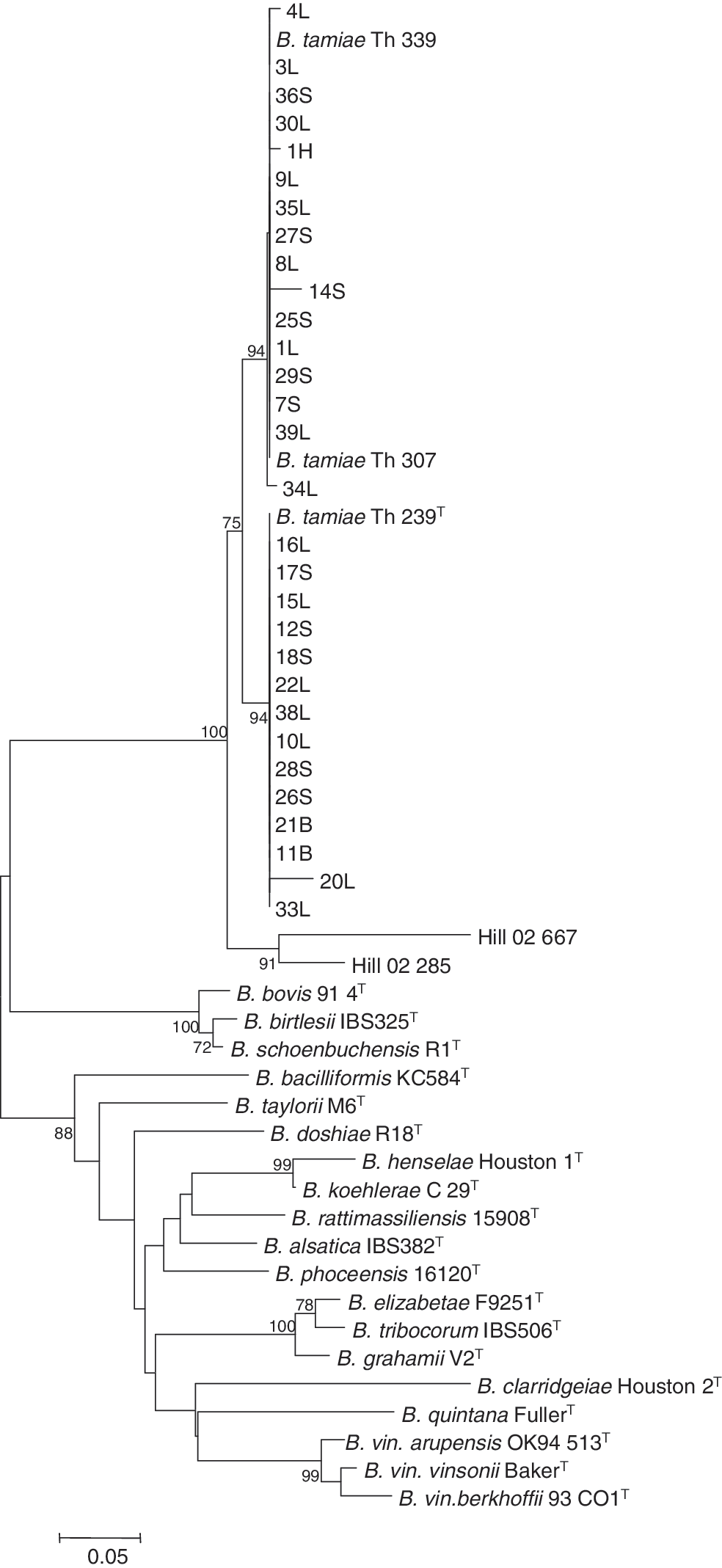
Phylogenetic analysis of the sequences of intergenic spacer (ITS) from Bartonella detected in ectoparasites collected from Thai rodents. The phylogenetic tree was constructed by the neighbor-joining method; bootstrap values were estimated with 1000 replicates. Only bootstrap replicates >70% are noted. The 30 samples that were positive for ITS were classified into two (ITS239 and ITS307/339) clusters. The sequence detected from an Amblyoma americanum tick (Hill-02-28, Hill-02-66; Billeter et al. 2008b) was also included. Sample ID represents sample no., followed by the genus of ectoparasite (L for Leptotrombidium, S for Schoengastia, B for Blankarrtia, and H for Haemaphysalis).

Phylogenetic analysis of the sequences of gltA from Bartonella detected in ectoparasites collected from Thai rodents. The phylogenetic tree was constructed by the neighbor-joining method; bootstrap values were estimated with 1000 replicates. Only bootstrap replicates >70% are noted. The nine samples that were positive for gltA were classified into the same cluster. Sample ID represents sample no., followed by the genus of ectoparasite (L for Leptotrombidium, S for Schoengastia, and B for Blankarrtia). The sequences from Brucella melitensis 16 MT and Agrobacterium tumefaciens C58T were used as out-group bacteria.
The sequence groups of ITS239 and ITS307/339 were 29.2% (7/24) and 37.5% (9/24), respectively, among mites of genus Leptotrombidium, 35.7% (5/14) and 42.9% (6/14) among mites of genus Schoengastia, and 100% (2/2) and 0% (0/2) among mites of genus Blankarrtia, respectively (Table 2). The two sequence groups were detected in 20.7% (6/29) and 44.8% (13/29) of mites collected from R. rattus, 50.0% (1/2) and 0% (0/2) of mites from R. argentiventer, 80.0% (4/5) and 20.0% (1/5) of mites from B. indica, 66.7% (2/3) and 33.3% (1/3) of mites B. savilei, and 50% (1/2) and 50% (1/2) of mites from M. cervicolor, respectively (Table 3). Of the 14 total sequence positives found in the ITS239 group, 4 were isolated from ectoparasites collected in the northern region of Thailand, whereas the other 10 came from the southern region of Thailand. Of the 16 total ITS sequence positives in the ITS307/339 group, 3 were found in the northern region of Thailand, 6 from the northeastern region, 2 from the central region, and 5 from the southern region (Table 4).
This is a tick pool.
This includes a tick pool.
Discussion
In our study, we detected DNA specific for B. tamiae in mites collected from Thai rodents; this, therefore, represents one of the few reports showing a potential role of mites in the transmission of Bartonella (Durden et al. 2004, Billeter et al. 2008a). Percentage of the sequence homologies between the detected DNA in this study and those of the known B. tamiae were 97.8–100% for ITS307/339 group and 97.7–99.6% for ITS239 group, respectively, and 98.3–100% for the gltA gene. The first study on mites in the transmission of Bartonella species was reported more than 60 years ago when Baker (1946) showed that hamsters inoculated with suspension of mites collected from Microtus voles from Canada became bacteremic with an agent that was later identified as B. vinsonii. More recently, Kim et al. (2005) tested 21 Mesostigmatid mite pool samples and found 4 (19%) positive pools by PCR targeting the 16S rRNA gene; the DNA sequence showed a high homology (99.2%) with B. doshiae. Reeves et al. (2006) demonstrated that a Steatonyssus sp., a mite removed from a bat, harbored a Bartonella sp. (96%) that was closely related to an unnamed Bartonella found in rodents. Reeves et al. (2007) detected a part of the groEL gene from one of eight pools of tropical rat mites (Ornithonyssus bacoti) collected from R. rattus in Egypt by sequence analysis and demonstrated it to be an unique sequence with 81% similarity to a Bartonella species. In addition to mites associated with rodents, evidence of Bartonella DNA has been reported from mites of bats (Chiroptera, Steatonyssus sp.) and house dust mites (Dermatophagoides farinae and D. pteronyssinus) (Valerio et al. 2005, Reeves et al. 2006).
To the best of our knowledge, this is the first report of the presence of Bartonella DNA in the chigger mite. Durden et al. (2004) reported that none of the examined chigger mites collected from gray squirrels was positive for Bartonella by PCR targeting of gltA gene. Our study is also the first report showing the detection of B. tamiae DNA in chigger mites. B. tamiae is a novel Bartonella species that was isolated from patients in Thailand (Kosoy et al. 2008). Strains Th239, Th307, and Th339 were isolated from three separate patients. Although all the patients reported exposures to rats, the role of the rats in the transmission of B. tamiae has not been proved (Kosoy et al. 2008). Interestingly, the DNA sequence of gltA which we identified in mites infesting rodents were highly similar to those of B. tamiae isolated from Thai patients. These findings suggest a possible role of the chigger mites in the transmission of B. tamiae to humans.
One tick of the genus Haemaphysalis was positive for DNA specific for the sequence group of ITS307/339. Recently, Billeter et al. (2008b) also reported the presence of B. tamiae-like sequences in two A. americanum by using the same primer pairs that were used in our study. Thus, ticks as well as chigger mites might play a role in the transmission of B. tamiae.
We identified two distinct sequence groups, ITS307/339 and ITS239, which show high sequence similarity with the isolates from the patients. The 16 sequences belonging to the sequence group ITS307/339 and 14 sequences belonging to the sequence group ITS239 demonstrated only 0-4 and 1-4 single-nucleotide polymorphisms (SNPs) among the sequences for each group, respectively. Only one SNP was found between the sequences of B. tamiae strains Th307/339 and that from the tick examined in this study. In comparison, the DNA that was detected in the A. americanum tick collected in Virginia was distinguishable from three cultures of B. tamiae isolated from Thai patients by the presence of several deletions and SNPs (Hill-02-28, Hill-02-66; Fig. 1). These results suggest the possibility that indigenous B. tamiae-like agents are distributed in geographically different areas.
To date, there have been no reports showing the isolation of B. tamiae or detection of B. tamiae-specific DNA from any animals, including rats, in Thailand (Castle et al. 2004). One of possible explanations is that chigger mites themselves serve as the principal natural reservoirs of B. tamiae and do not require involvement of the rodents for the transmission cycle. Because chigger mites generally feed only on a single host in their larval stage, they do not have an opportunity to transmit acquired pathogens during a subsequent feeding (Mullen and Durden 2002). There are three possible ways by which these mites may therefore play a role in the transmission of Bartonella. First, the mite may transmit B. tamiae transovarially and/or transstadially, thus resulting in the preservation of the organisms among the chigger mites, similar to the parasitic system of O. tsutsugamushi and Leptotrombidium mites. Second, the organisms may persist in the body of the rodents outside of their blood. Because chigger mites feed on body fluids and partially digested tissues rather than blood, some Bartonella species could potentially be acquired by chiggers from the former two sources. Third, the organisms may exist in the blood but are not cultivable on agar medium. Also, it is possible that more prevalent Bartonella species in the blood of rodents might have prevented the detection of B. tamiae. Additional investigations are needed to clarify the source of infection and the mode of transmission of B. tamiae infection in Thailand.
When comparing the PCR results of the two target regions used here for the detection of B. tamiae DNA, ITS was clearly more sensitive than gltA. Among the 34 positive samples, 25 were detected by the ITS target only and 4 were positive for gltA; only 5 were positive for both targets (ITS and gltA). The gltA primers used for the detection of B. tamiae were specifically designed for this study because preliminary results demonstrated that the application of the primers proposed by Norman et al. (1995) was not effective. Our results suggest that ITS primers are the most effective tool for screening ectoparasites for the presence of B. tamiae. Application of the same primers allowed Billeter et al. (2008b) to detect B. tamiae DNA in ticks. It would therefore be an important assignment to design a primer pair that amplifies all Bartonella species including B. tamiae more specifically.
Footnotes
Acknowledgment
The authors express their gratitude to Taweesak Monkanna, Surachai Leepitakrat, Weerayut Charoensongserkit, Warinpassorn Leepitakrat, and Sucheera Insuan from the Department of Entomology, AFRIMS, Bangkok, Thailand, for assistance in collecting and processing ectoparasite specimens.
Disclosure Statement
No competing financial interests exist.