
Abstract
Dengue has affected the north provinces of Argentina, mainly Salta province. The 2009 outbreak, with 5 deaths and >27,000 infected, was the most important, and the first to extend into the central area of the country. This article includes research on seasonal Aedes aegypti abundance variation in Orán City (Salta province), and determination of the date of mosquito population increase and an estimation of the date of maximum rate of increase as well as the intrinsic rate of natural increase (r), to detect the optimal time to apply vector control measures. Between September 2005 and March 2007, ovitraps were randomly distributed in the city to collect Ae. aegypti eggs. The variation observed in the number of collected eggs was described by fitting a third-degree polynomial by the least square method, allowing to determine the time when population increase began (week 1), after the temperate and dry season. Eggs were collected throughout the year, with the highest variation in abundance during the warm and rainy season, and the maximum value registered in February 2007. The rate of increase of the number of eggs laid per week peaked between weeks 9 and 10 after the beginning of the population increase (week 1). Week 1 depends on temperature, it occurs after getting over the thermal threshold and the needed accumulation of 160 degree-day is reached. Consequently, week 1 changes depending on temperature. Peak abundance of eggs during 2005–2006 was recorded on week 15 (after week 1); during 2006–2007, the peak was observed on week 22. Estimation of the intrinsic rate of natural increase (r) of Ae. aegypti is useful not only to determine optimal time to apply vector control measures with better cost–benefit, but also to add an insecticide control strategy against the vector to diminish the possibility of resistance.
Introduction
Ae. aegypti was considered eradicated from Argentina in 1963; nevertheless, in 1986 the Argentina Ministry of Health reported a reinfestation of the northern provinces of Formosa and Misiones (Avilés et al. 1999, PAHO 2001). The first dengue cases registered in the northwestern province of Salta in 1997 were due to serotype DEN-2 (Avilés et al. 1999). Since then, dengue has become endemic in the Argentine northwest, especially in Salta. The second outbreak, due to DEN-1, took place in 2000. It mainly affected the provinces of Formosa and Misiones (Northeast Argentina), in relation with neighboring countries where outbreaks were occurring at the same time (Avilés et al. 2003). While 214 cases of DEN-1 were detected in Salta in 2002, DEN-3 was registered for the first time in the province of Misiones (Ministerio de Salud de la Nación 2004). By 2003, DEN-1, −2, and −3 were detected, so the three virus serotypes were co-circulating in the north of Argentina (PAHO 2004).
Studies carried out in the provinces of Salta (Avilés et al. 1999), Córdoba (Almirón and Asis 2003), and Buenos Aires (Vezanni et al. 2004) show the risk of dengue outbreaks in Argentina. Between January and June 2009, the dengue outbreak peaked, being the most important in the history of the country. On June 9, 2009, there were 5 confirmed deaths and 27,752 infected, among autochthonous (25,828 in 13 provinces) and imported cases (886). About 92% of cases occurred in the provinces of Chaco (46%), Catamarca (36%), and Salta (10%) (Ministerio de Salud de la Nación 2009). The 2009 dengue outbreak was the first one to extend into central Argentina, where autochthonous cases were recorded in the provinces of Córdoba, Santa Fe, Buenos Aires, Santiago del Estero, and Catamarca, even affecting important cities such as Buenos Aires and Córdoba (Ministerio de Salud de Córdoba 2009).
According to Pan American Health Organization, the main factors of dengue resurgence in the region are the global climate change and deficient infrastructure (especially if household water storage is common, and solid waste disposal and health services are inadequate) in cities were population increased dramatically (Ministerio de Salud de la Nación 2009). In the Americas, control vector effectiveness has decreased due to a variety of factors (PAHO 1989), such as nonsustained vector control actions over time, which are based mainly on chemical control, and community participation as well as health education, but only during epidemics and outbreaks (San Martín and Brathwaite-Dick 2007). The World Health Organization (WHO) classifies dengue as a major international public health problem because of the geographical expansion of both the mosquito vector and the dengue virus, the increased frequency of epidemics, the co-circulation of multiple virus serotypes, and the re-emergence of dengue hemorrhagic fever in new areas (Gibbons and Vaughn 2002, Gubler 2002).
In Argentina, the actual vector distribution encompasses an area that begins in the northern provinces and extends to the central province of La Pampa (Ministerio de Salud de la Nación 2009), which surpasses the historical distribution limit of the vector; therefore, the number of people who could be in contact with the vector increasing virus transmission probability increases (WHO 2009).
Since there is no vaccine to protect against dengue, the only method for controlling or preventing dengue virus transmission is to combat the vector mosquitoes (Guzmán and Kouri 2002, Guzmán et al. 2004, WHO 2009). Setting ovitraps serves as a method of vector detection (Kitron et al. 1989, Chadee and Corbet 1990, Chadee et al. 1995, Reiter et al. 1995). Ovitraps data have been reported to be more sensitive than the traditional Stegomyia indices in detecting low population densities of the vector (Focks 2003). Moreover, ovitraps have provided useful data on spatial and temporal distributions of Ae. aegypti and other container-inhabiting mosquito species, allowing to obtain a better knowledge of the vector activity (Ritchie 1984).
The aim of this work was to generate a method to forecast dates of vector maximum population increase (the intrinsic rate of natural increase “r”), that is, the rate of population increase per individual and unit time (Begon et al. 1988), by using ovitraps. The intrinsic rate of natural increase (r) is useful for forecasting insect population increase under certain environmental conditions (Ricklefs and Millar 2000, Southwood and Henderson 2000, Andrewartha and Birch 2005). Biological parameters and principal vital statistics of an insect plague population are basic tools to elaborate control strategies (Southwood and Henderson 2000). Knowing estimations such as the intrinsic rate of natural increase (r) of the species is useful not only to determine the optimal time to apply control measures with the most cost–benefit, but also to maximize chemical control strategies against vector population that could be resistant (WHO 1970).
Thereby, our objectives were to study the seasonal variation of Ae. aegypti abundance in the city of Orán, to determine the likely dates of the beginning of the seasonal activity and to estimate the intrinsic rate of natural increase (r) and (dN/dT) as an alternative to detect the optimal time to apply control strategies.
Materials and Methods
Study site
Field studies were conducted at the city of San Ramón de la Nueva Orán, hereafter Orán City (23°08′ S, 64°20′ W, elevation 337 m.a.s.l.), head of Orán department with 3562 km2 (Salta province) and 270 km from the city of Salta, the capital of the province of Salta (Fig. 1). Orán's urban area covers ∼20 km2 with 72,712 inhabitants (INDEC 2001). The city is located in the phyto-geographic area of Yungas, which has a subtropical climate, with a dry season of about 6 months (June–November) in which the precipitations are below 50 mm per month, and a rainy period between November and April. The city has mean annual rainfall of 1000 mm (Brown et al. 2001), mean temperature of 21°C, and a mean relative humidity of 70%. The temperature varies considerably both during the day and along the year. The year could be divided into three periods: spring (September–December) is hot and dry, summer (January–April) is warm and rainy, and autumn–winter (May–August) is cold and dry (Brown 1995).
Map of Argentina showing Salta Province with Orán City (23°08′ S, 64°20′ W).
The city is surrounded by native forest and crop areas, mainly sugarcane, grains, citrus (grapefruit, orange, lemon, and tangerine), scoop horticulture (tomato, corn, zucchini, and eggplant), tropical fruit (banana, avocado, and mango), and forest trees like Eucalyptus spp. (INTA 2004).
Ae. aegypti sampling
Sampling was performed by using ovitraps, consisting of 350 mL plastic cups, containing 250 mL of hay infusion prepared a week before use. According to the WHO, hay infusion results an effective attractant to gravid Ae. aegypti female mosquitoes (Reiter et al. 1991, Reiter and Gubler 1997, Reiter and Nathan 2001). Ninety houses were randomly selected in the urban area of the city of Orán. One ovitrap was placed in a shaded site at ground level in the backyard of each house (Reiter and Nathan 2001) and weekly replaced from September 2005 to March 2007. Each week, after replacing ovitraps, at the laboratory the egg-papers were removed and eggs counted. The eggs are visible to the naked eye but, for accuracy, are best counted with the aid of a magnifying glass at ×10 magnification (Reiter and Nathan 2001). Then, the number of eggs per paper was recorded on a field form. On northwestern Argentina Ae. aegypti is the only Aedes species recorded at the moment. Besides that, egg-papers from several ovitraps were randomly selected and kept moist for 2–3 days as recommend by WHO (Reiter and Nathan 2001) and then the eggs were hatched to third or fourth instar and examined to corroborate that belonged to Ae. aegypti. No other species but Ae. aegypti was recorded.
These ovitraps provided useful data regarding temporal variation of vector population density (Loop 1957, Reiter and Gubler 1997).
Meteorological data
Daily maximum and minimum temperature (°C), humidity (%), and rainfall (mm) were provided by Orán City National Meteorological Station (Servicio Meteorológico Nacional).
Data analysis
Ae. aegypti population size was weekly estimated through the sum of egg-catches of the 90 sampled houses of the city. Since the main aim of this work was to forecast the time of the year when vector density increase is maximum (r), data were analyzed separately in two periods that included the maximum population peaks observed, that is, September 2005 to March 2006 (spring–summer) and September 2006 to March 2007. Description of the observed variation in each spring–summer period was determined by fitting a third-degree polynomial by the least square method of the egg-catches number from ovitraps that respond to the following equation:
where N(t) is the weekly egg-catch number, t is time (weeks), and a 0, a 1, a 2, and a 3 are constants. The third-degree polynomials have the useful property of having both a maximum and a minimum point, in this case N(t), that help to describe the seasonality of Ae. aegypti egg-catches.
The fitting is necessary to determine the week in which oviposition activity begins to increase (week 1) for each spring–summer period studied. When the rainy season begins, following the temperate-cold and dry period, eggs begin to hatch, followed by larval and pupal development and finally adult emergence. As a result, the vector activity increases, which is reflected in an increasing egg-catch number. It was possible to predict such increase (week 1) starting to count from the date on which average minimum temperatures for last 7 days exceeded the development thermal threshold of 12.8°C, estimated for Ae. aegypti in Córdoba (temperate zone on the central part of the country) by Domínguez et al. (2000), while minimum temperature did not decrease significantly (at least during the next 3 weeks). As can be inferred from the results reported by Domínguez et al. (2000), that time is considered “week 1,” since 160 degree-days must be accumulated, being the physiological time required for newborn larvae to become adults. A rise in the number of eggs in ovitraps can be observed from this week on.
The rate of increase of the egg-catch number is determined from the derivative of N(t) as a function of time (derivative of Equation 1), that is
where dN/dT is the instant rate of increase (derivative) from N (eggs number/week) as function of time t (weeks), and a
0, a
1, a
2, and a
3 are the respective constants from Equation 1. According to Equation 2, the rate of increase changes over time showing a maximum rate value, when the derivative is null (second derivative of the abundance), provided that the third derivative is negative:
which, using the least square values obtained from Equation 1, will determine population size and therefore the per capita rate of increase. The latter is obtained by dividing the population rate of increase by the population size at every time [(1/N)(dN/dT)], and whose maximum value is the parameter r (intrinsic rate of natural increase).
Points of maximum population rates of increase tend to come mid-way between seasonal minima and maxima. However, they do not represent points of maximum per capita rate of increase (which occur just after the seasonal minima).
Results
Ae. aegypti population showed wide egg abundance variation with maximum values in February 2006 (7967 eggs) and February 2007 (14,099 eggs) (Fig. 2). During May and September 2006 (temperate-cold and dry period), the egg numbers were low (Fig. 2). Ae. aegypti population in the city of Orán remains active during these periods because residents maintain breeding sites by continuous irrigation of cultivated plants (house garden and greenhouses) and because temperature is not very unfavorable (∼10°C mean minimum). These conditions allowed some emerged females to survive in late autumn, and to lay eggs in the ovitraps during that period. It is also possible that at least some immature individuals were able to complete their development and that emerged adults could find the necessary resources to feed and breed near houses and domestic areas. Egg abundance began to rise at increasing rate at the end of that season, when daily minimum temperature exceeded the growth thermal threshold, allowing for a greater number of individuals and an increasingly rapid development. That was evident when the required 160 degree-days were accumulated. Such conditions were observed when there was increased activity (week 1) corresponding to dates from October 13–20, 2005, and September 27 to October 4, 2006.
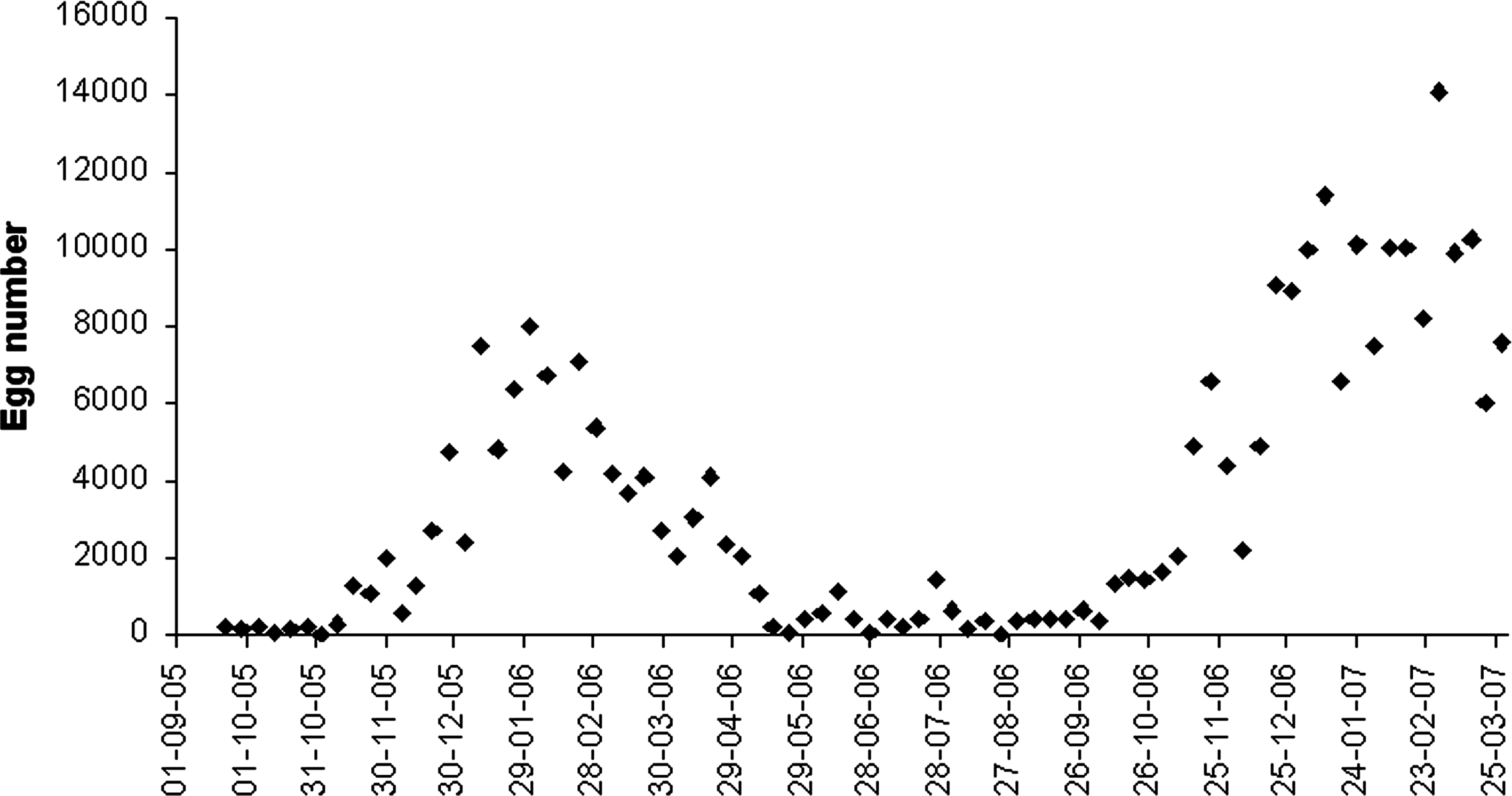
Temporal fluctuation of Aedes aegypti eggs from September 2005 to March 2007.
Third-degree polynomials fitted to data of each study period gave the following equations:

The vertical line C marks the week on which the population rate of increase was calculated to be at a maximum for the period 2005–2006.
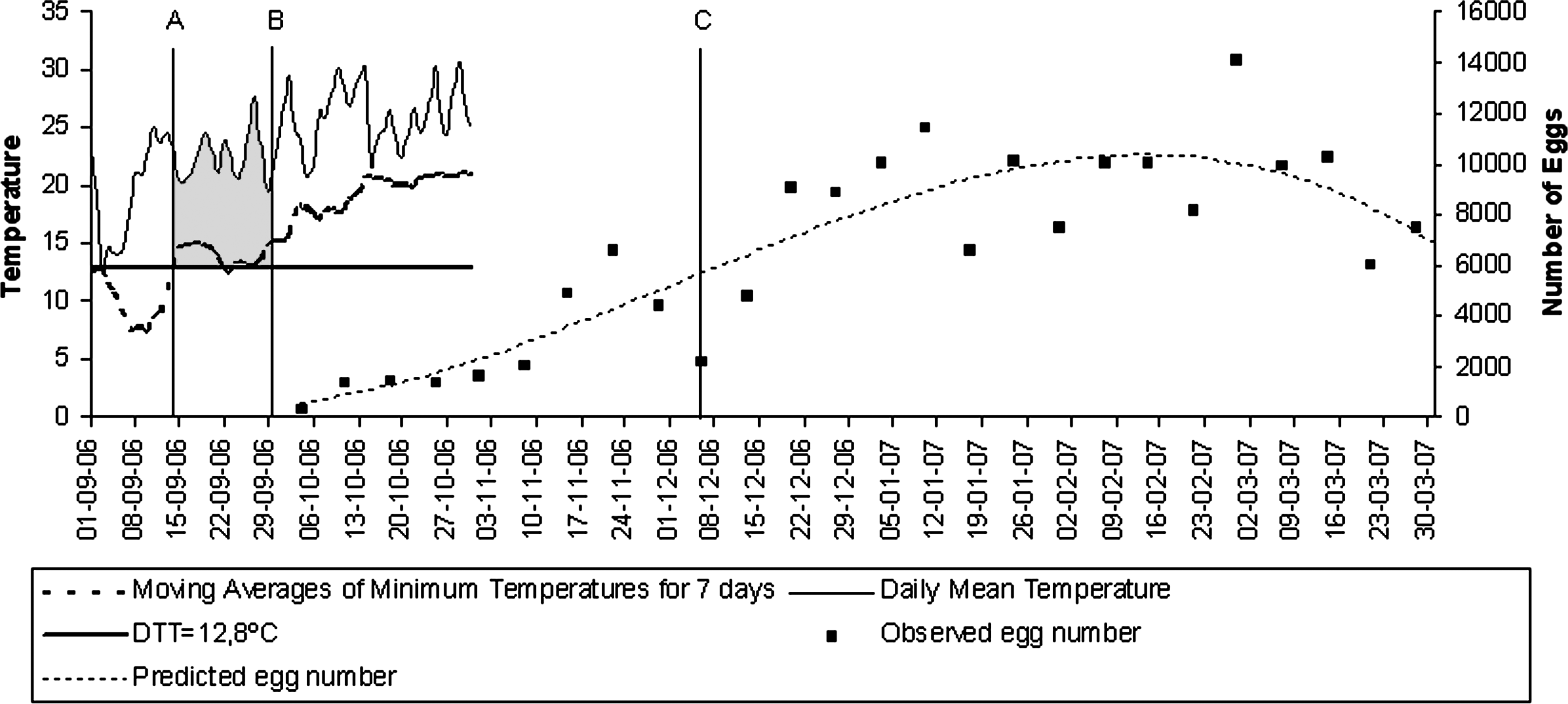
The vertical line C marks the week on which the population rate of increase was calculated to be at a maximum for the period 2006–2007.
For 2005–2006 period
For 2006–2007 period:
The maximum egg abundance at the first study period was recorded on week 15, from week 1, according to the thermal threshold, and to the accumulation of 160 degree-day with 7967 eggs (vertical line C on Fig. 3). For the second period, corresponding to week 22, with 14,099 eggs (vertical line C on Fig. 4), the per capita rate values were 0.2012 and 0.1387, respectively. The maximum egg abundance registered in the second year was 77% higher than that in the first year. The maximum observed egg abundance value in each period could be directly associated to rainfall occurred in the previous winter and spring. Accordingly, the amount of precipitation recorded in winter and spring of the second year (328.9 mm) was 72% higher than in the first year (190.9 mm).
Discussion
In several regions of the world, endemic diseases show significant seasonal fluctuations. Dengue fever shows important seasonal patterns whereby transmission is highest in the months of heavy rainfall and humidity (Vezzani et al. 2004, Hopp and Foley 2001). The most significant climatic factors for vector-borne diseases seasonality include temperature and rainfall. Increased precipitation may increase vector abundance by expanding the size of existent larval habitat and creating new breeding sites (Patz et al. 2003), and consequently increase the potential for dengue transmission (Hales et al. 2003). Even though the mosquito population was always active during the dry season in the city of Orán, in this research the highest Ae. aegypti abundance was found to be associated with high amounts of rainfall. This could be due to Ae. aegypti breeding habitats generally are small water-holding containers, such as plant pots and discarded bottles and tires (Hales et al. 2003), which thus maintain the active vector population during winter.
Temperature is also a key factor affecting mosquito populations. Domínguez et al. (2000) determined the developmental thermal threshold at 12.8°C for Ae. aegypti at semi natural conditions in Córdoba City (Argentina). In Taiwan, Chen and Huang (1988) calculated a 13.4°C development thermal threshold, quite close to the estimation obtained in Córdoba. Similar values were obtained at other laboratory studies (Bar-Zeev 1958, Lee 1994, Tun-Lin et al. 2000). Therefore, the one calculated by Domínguez et al. (2000) was used in this work to determine week 1 (the Ae. aegypti oviposition activity increase after dry season) for the city of Orán. Adult activity is also influenced by temperature. According to Chen and Huang (1988), in Taiwan the low winter temperatures limit Ae. aegypti dispersal. During temperate-winter and dry period in the city of Orán, the vector activity was considerably diminished compared to the warm and rainy period, but it was always active, and therefore precipitations could be the limiting factor more than temperature in our study area. In the city of Córdoba (Argentina), oviposition activity was not observed below 17°C (Domínguez et al. 2000), in agreement with data reported by Campos and Maciá (1996) for the temperate climate of the city of Quilmes (Buenos Aires province). However, oviposition was not recorded below 20°C in the city of Buenos Aires (Schweigmann et al. 1996). In the city of Orán, the lowest mean weekly temperature recorded during 2006 was 13.7°C in the last week of July; nevertheless, oviposition activity was registered during July.
Mosquito abundance peaks may be conditioned by climatic variability (De Garín et al. 2000) and the effect of temperature over changes in the growth rate could also affect the feeding rate as well as the vector–host contact rate (Patz et al. 2003). A rise in the minimum temperature could have the most significant effect over vector abundance allowing for the invasion of new areas and permanence throughout the year (De Garín et al. 2000).
Therefore, in the city of Oran we took minimum temperature as one of the most important parameters to calculate the initial week of oviposition activity increment (week 1). According to our results, week 9 after activity increase starts is when the maximum growth population rate occurred, the appropriate moment to combat Ae. aegypti from an ecological point of view. Thus, the mosquito population could be regulated by preventing maximum vector abundance, which, according to our research, corresponds to the month of December for both periods of study. During the 2009 dengue outbreak in the city of Orán, cases began to be reported in January (Ministerio de Salud de la Nación 2009), just after the maximum vector population growth occurred. However, from a sanitary point of view, it would be ideal to apply integrated management measures, by working on appropriate environmental sanitation each year during the months of August and September, considering that these are the months of maximum per capita rate of increase and that oviposition activity increases at this time of the year. Future outbreaks could be avoided this way.
The contribution of this work consists of providing a simple method to predict the time of highest activity of Ae. aegypti, that is, to determine when week 1 occurs, and help to adopt vector control measures. To do that it is necessary to count on a daily temperature register for the study city.
The proposed method could be adopted to implement control strategies to prevent future outbreaks in the city of Orán. It could also be applied to other regions in the argentine northwest, as well as provinces such as Córdoba, where Ae. aegypti activity was registered between October and May (Domínguez et al. 2000), and even Buenos Aires (Campos y Maciá 1996), with a similar vector activity pattern to those in the city of Orán, though mosquitoes were active all year long in the city of Orán, while there was no recorded mosquito activity in central Argentina during the winter (Campos y Maciá 1996, Domínguez et al. 2000).
Footnotes
Acknowledgments
This work was partially funded by Red Latinoamericana de Control de Vectores (RELCOV), VIGI+A of the Ministerio de Salud de la Nación (National Health Ministry), and the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), SECyT–National University of Córdoba. WRA is a member of the Scientific Career of CONICET, Argentina.
Disclosure Statement
No competing financial interests exist.