
Abstract
To identify potential vector species of scrub typhus in the Republic of Korea (ROK), chigger mites were harvested from wild rodents captured at nine localities in October 2005. The bodies of the chigger mites were individually punctured with a fine pin, squeezed out internal contents, and examined for Orientia tsutsugamushi DNA by nested polymerase chain reaction. The exoskeleton of associated chiggers was mounted on glass slides with polyvinylalcohol (PVA) medium for identification. Among 830 individuals belonging to 4 genera and 14 species, O. tsutsugamushi was detected from 22 chiggers of six species, with an overall infection rate of 2.7%. The infection rate was highest for Leptotrombidium palpale (5.3%), followed by Neotrombicula japonica (4.3%), Leptotrombidium scutellare (3.7%), Leptotrombidium orientale (3.6%), Eushoengastia koreaensis (1.9%), and Leptotrombidium pallidum (1.5%). This study first reported O. tsutsugamushi infection from N. japonica and E. koreaensis larvae in the ROK. The population densities of L. pallidum (33.4 chiggers/rodent), historically confirmed as a primary vector of scrub typhus in the ROK, were high, whereas its infection rate was relatively low (1.5%). However, E. koreaensis was only collected from 154 individuals at seven collection sites and its infection rate was demonstrated relatively high (mean 1.9%). Additional studies are needed to determine the role of vector species in the epidemiology of scrub typhus.
Introduction
Presently, >56 species, belonging to 14 genera, have been identified as vector species of scrub typhus. Leptotrombidium deliense, Leptotrombidium fletchery, Leptotrombidium arenicola, Leptotrombidium pallidum, Leptotrombidium pavlovskyi, and Leptotrombidium scutellare are regarded as the primary vectors of scrub typhus in several countries. In Japan, >16 species, belonging to six genera, were confirmed harboring O. tsutsugamushi, with Leptotrombidium akamushi, L. pallidum, and L. scutellare being the principal vectors (Kawamura et al. 1995). In the ROK, Jackson et al. (1957) first reported the vector species of scrub typhus. Ree et al. (1991) confirmed L. pallidum as a vector of O. tsutsugamushi in the ROK and subsequently incriminated L. scutellare collected from Jeju Island as a vector (Ree et al. 1992) by applying indirect immunofluorescent antibody (IFA) test to internal tissue samples of individual engorged chigger mites. Through polymerase chain reaction (PCR) methods, Ree et al. (1997) further detected O. tsutsugamushi from L. pallidum, L. scutellare, Leptotrombidium palpale, Leptotrombidium orientale, and Leptotrombidium zetum. In the present study, we identified infection rates of O. tsutsugamushi in chigger mites, which were harvested from rodents captured from high- and low-endemic areas by nested PCR methods. An understanding of the ecology, distribution, and epidemiological role of chigger species is necessary to develop comprehensive scrub typhus control strategies.
Materials and Methods
Study areas and periods
Chigger mites were collected from rodents captured at nine collection sites during October 2005 (Fig. 1). Three site localities among them (Jangan, Gyeonggi Province; Gurye, Jeollanam Province; Hapcheon, Gyeongnam Province) were located in high endemic areas of scrub typhus, and the other six sites were located in low endemic areas.
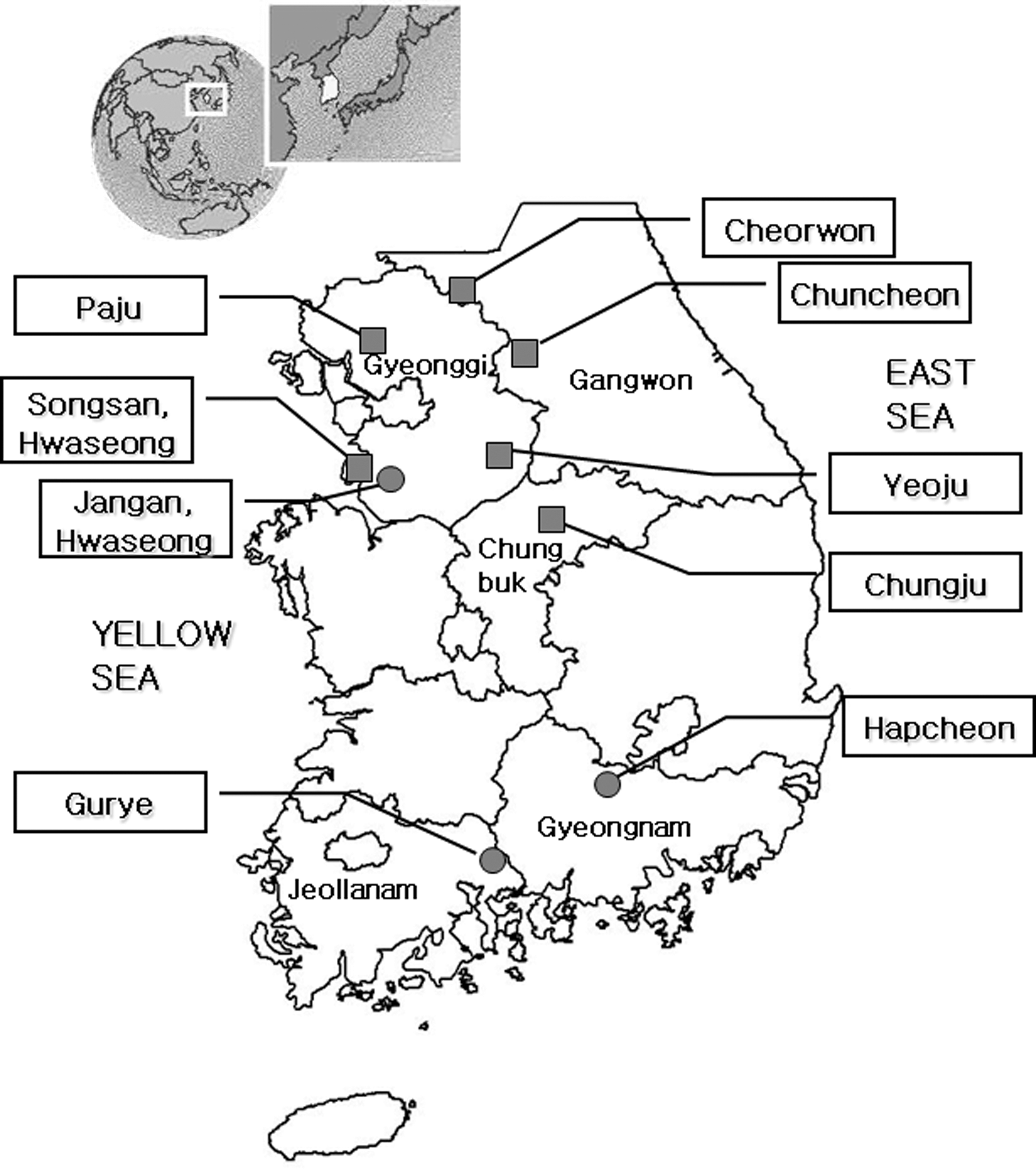
Study area. Dots (high endemic area) and squares (low endemic or no case area) indicate the collection sites of rodents and chiggers.
Rodents and chigger collection
Five to seven habitats around each collection site with similar biotic characteristics were surveyed. A total of 10–15 Sherman live traps (8 × 9 × 23 cm, H.B. Sherman Traps), baited with rolled oats and peanut butter mixture, were set at 2–3-m intervals between 17:00 and 18:00 h and collected at 24:00 h to reduce trap mortality. Rodents were euthanized under an approved animal use protocol (KCDC 2005) and hung over a glass bowl with tap water for 24 h to harvest chigger mites. The chiggers were removed from the water surface with a fine brush or loop, placed in a chigger-rearing container with charcoal–plaster mixture base, and maintained at 4°C until they were dissected for O. tsutsugamushi detection.
Preparation of chigger mites for PCR and identification
The method used by Dohany et al. (1978) and Ree et al. (1991) was applied for the detection of O. tsutsugamushi from individual chiggers. Individual chigger mites were placed on a glass slide with sterile phosphate-buffered saline (pH 7.2) under a stereomicroscope. The chigger body was punctured with a fine needle and its internal contents were removed by applying pressure to its abdomen. The internal contents of chiggers were placed individually into 1.5-mL Eppendorf tubes with phosphate-buffered saline and thoroughly mixed with an autopipette. DNA was extracted from the individual sample mixture using a DNA purification kit (Gentra). The exoskeletons of individual chiggers were transferred to glass slides, mounted with PVA medium (Bioquip), and identified to species level by using a morphological key (Ree 1990).
Nested PCR
Nested PCR assay was performed as described by Furuya et al. (1993), with modifications. The nucleotide primers encoding the 56-kDa antigen of the Gilliam strain of O. tsutsugamushi were used. Primers 34 (5′-TCA AGC TTA TTG CTA GTG CAA TGT CTGC-3′) and 55 (5′-AGG GAT CCC TGC TGC TGT GCT TGC TGC G-3′) were used for the first PCR, whereas nested PCR primers 10 (5′-GAT CAA GCT TCC TCA GCC TAC TAT AAT GCC-3′) and 11 (5′-CTA GGG ATC CCG ACA GAT GCA CTA TTA GGC-3′) were used to amplify a 483-bp fragment.
The first PCR was carried out with 5 μL of DNA, extracted from the abdominal contents of the mite, in a mixture (final volume 50 μL) containing 2 mM MgCl2, 50 mM KCl, 10 mM Tris-HCl (pH 8.3), 200 μM of deoxynucleotide triphosphates, and 200 μM of 1.25 U Taq DNA polymerase (Takara). A total of 0.1 μM of primer was used for the first PCR reaction, followed by 0.2 μM of primer for the second PCR reaction. The amplification conditions consisted of initial polymerase activation at 94°C for 5 min; 30 cycles at 94°C for 30 s, 55°C for 2 min, and 72°C for 2 min; and a final elongation step at 72°C for 7 min using a DNA Peltier thermal cycler (MJ Research). The PCR products were analyzed by electrophoresis on 1.5% agarose gel stained with ethidium bromide and examined under an ultraviolet trans-illuminator.
Analysis of the nucleotide sequence homology was performed using the Clustal W program (DNA STAR) to verify gene fragments as O. tsutsugamushi.
Results
Chigger population
A total of 6350 chiggers with a chigger index (CI) of 52.6 (chiggers/rodent) were collected from 115 Apodemus agrarius (Table 1). Cheorwon, Gangwon Province, showed the highest CI (304.6), followed by Songsan (104.2). On the other hand, the CI was low in Chuncheon (5.3), Hapcheon (6.2), and Yeoju (7.8).
Others: minor species and nonvector species.
Chigger index (CI) = total no. of chiggers/total no. of Apodemus agrarius.
L. pallidum was the most widely distributed chigger species throughout the whole study area, but was especially prevalent in the northern part of the ROK. In Gyeonggi and Gangwon Province, L. pallidum was the predominant species, accounting for >50% of the total harvested chiggers. The species composition in Chungju was variable: L. orientale and Eushoengastia koreaensis accounted for 33.6% and 27.0% of the total chiggers found in Chungju, respectively, whereas L. pallidum accounted for only 18.9% (7.3 CI). In the southern part of the ROK, L. scutellare was the predominant species, accounting for >50% of the chigger population, whereas the number of L. pallidum was very small. Songsan showed diverse species composition with 12 species belonging to three genera. In this study, we collected L. scutellare at Jangan and Songsan, Gyeonggi Province. This collection has been the northernmost collection of L. scutellare in the ROK.
Vector determination
Among 6350 harvested chiggers, we randomly selected 20 chiggers from each rodent and dissected 830 chiggers to detect O. tsutsugamushi DNA by PCR. A total of 22 chiggers (2.7%) were positive for O. tsutsugamushi (Table 2). Yeoju, Gyeonggi Province, showed the highest positive rate (13.6%) among all collection sites, followed by Hapcheon (9.9%), Cheorwon (3.7%), Chungju (2.6%), Paju (2.5%), and Songsan (0.5%). Specific DNA of O. tsutsugamushi was detected from the members of three genera and six species, including L. pallidum, L. orientale, L. palpale, L. scutellare, E. koreaensis, and Neotrombicula japonica.
Values in parentheses indicate the no. of positive chiggers.
Infection rate = (no. of positive chiggers/total no. of examined chiggers) × 100.
Discussion
Jackson et al. (1957) first reported L. pallidum as a scrub typhus vector in the ROK. They collected engorged chiggers from A. agrarius and inoculated L. pallidum into laboratory mice. Then they isolated O. tsutsugamushi from these mice and its progeny of chigger mites. Ree et al. (1991) also confirmed L. pallidum as a vector by applying IFA test to the internal contents of chigger mites. With the same method, they found O. tsutsugamushi from 6 of 1142 (0.5%) L. scutellare collected from Jeju Island. Through PCR, Ree et al. (1997) detected O. tsutsugamushi from 65 chiggers belonging to five species: L. palpale, L. orientale, Leptotrombidium zetum, L. pallidum, and L. scutellare. We also detected O. tsutsugamushi from 22 chiggers of six species. Among these species, L. pallidum, L. palpale, L. scutellare, and L. orientale were already reported as vectors in the ROK (Ree et al. 1997). Even though N. japonica (Rehacek and Tarasevich 1988, Kawamura et al. 1995, Pham et al. 2001) has been known as a vector at outside of the ROK, this is the first report of it as a vector of O. tsutsugamushi in the ROK. In addition, we were the first in the world to detect O. tsutsugamushi in E. koreaensis. Infection rates from harvested chigger mites in the present study are slightly higher than that in a previous study (Ree et al. 1997). Although the infection rates varied from 0.5% to 13.6% (mean 2.7%) over all study sites, the chigger infection rates did not represent epidemic situation of collection sites. Pham et al. (2001) also found a lower infection rate (3.3%) in endemic area than that in nonendemic area (10.2%).
L. pallidum is widely distributed throughout the ROK. In the present study, its infection rate was 1.5%, which is similar to the rates of 1.6% obtained by Ree et al. (1997). However, Ree et al. (1997) reported infection rates lower than that reported in the present study: 1.8%–5.3% for L. palpale, 1.4%–3.7% for L. scutellare, and 1.6%–3.6% for L. orientale. Such differences may be ascribed to endemicity of collection sites and other environmental factors.
Even though the IFA assay is a subjective method, it has been widely used for the identification of scrub typhus infections because of its specificity, rapidity, and sensitivity. As a result of increased sensitivity of PCR techniques, the infection rate of L. pallidum increased from 0.5% (Ree et al. 1991) to 1.64% (Ree et al. 1997) because 27% of the suspected IFA-positive chigger mites were confirmed positive through PCR. Pham et al. (2001) also adopted the nested PCR method and detected three additional species (Leptotrombidium fuji, L. himizu, and Miyatrombicular kochiensis) as potential vectors. Thus, it is suggested that the PCR assay provides greater reliability in the identification of potential vectors and relative infection rates.
Chigger indices (number of chiggers per a rodent; CI) varied greatly depending on the collecting sites, ranging from 5.3 (Chuncheon) to 304.6 (Cheorwon). Although expecting that high endemic areas would display higher chigger population, we were unable to find any correlation between the CI and relative rates of scrub typhus. Especially, Jangan, where there were 91 cases from 2003 to 2005, showed relatively low CI (14.2) compared with Songsan (104.2), where there was no case reported during the same period. This result suggested that chigger populations alone do not explain scrub typhus epidemicity. Thus, scrub typhus infection rates, proximity to human populations to collection sites, human activities, relative proportion of female mites that pass on infections to their progeny, pathogenicity of the rickettsia, etc., must be considered in health risk analysis and development of control strategies because these factors could influence human case solely or in combination (Pham et al. 2001).
We used chiggers obtained from A. agrarius. Some researchers suspected that chiggers of nonvector species would harbor rickettsia parasites for some time after they fed on an infected mouse (Toyokawa 1972, Takahashi et al. 1990, 1994). After engorgement, chiggers that acquired O. tsutsugamushi have no opportunity to transmit O. tsutsugamushi to humans because they do not feed again. In addition, O. tsutsugamushi acquired from these chiggers was transtadially transmitted to the adult stage, but rarely or not transmitted vertically to their progeny (Takahashi et al. 1994). Because of those opinions, some researchers used unfed chigger to determine the true reservoir of O. tsutsugamushi (Pham et al. 2001, Misumi et al. 2002, Takahashi et al. 2002). To determine vector species more clearly, we need to collect unfed chiggers by soil collection or black clothes/plates methods and test the infection status of each chigger.
Footnotes
Acknowledgment
This study was financially supported by the Korea Centers for Disease Control and Prevention under the budget for health promotion (no. 4800-4851-300).
Disclosure Statement
No competing financial interests exist.