
Abstract
This is the first report on the presence of Hepatozoon canis in Vulpes vulpes in Italy. During the years 2005 and 2006, a total of 119 foxes were collected and their spleen tissues were screened by microscopy, polymerase chain reaction, and sequencing. In the same area, 290 ticks were picked off from dogs or collected from the environment. Microscopy detected inclusion bodies regarded as belonging to the genus Hepatozoon in four samples, whereas molecular diagnostics evidenced 16 foxes (13.4%) and 6 ticks (2.1%) positive to H. canis. The H. canis isolates we found in foxes, compared with the strains we previously detected in dogs from the same area and with the strains found in foxes from other European countries, show a certain genetic heterogeneity. In fact, seven isolates cluster with the Italian dog strain and nine isolates cluster with the fox strain found in Spain and Slovakia; moreover, the dog's strain is closely related to one tick's isolate, and the strain found in three Rhipicephalus sanguineus and in one Ixodes ricinus collected from the environment cluster with the aforementioned Spanish and Slovak fox strains. Our findings confirm the importance of R. sanguineus as final host and suggest that I. ricinus might also be implicated in parasite transmission, explaining in that way the occurrence of hepatozoonosis in areas considered R. sanguineus-free. The peridomestic habits of V. vulpes and the increasing global temperature are expected to amplify the impact of this vector-borne disease and to enforce the transmission of Hepatozoon to domestic animals.
Introduction
Forty-six species belonging to the genus Hepatozoon have been described in mammals, which become infected by ingesting the arthropod containing infective sporozoites, whereas no transfer during bloodsucking has ever been demonstrated. The parasite develops in mammalian leucocytes. The brown dog tick, Rhipicephalus sanguineus, was identified by Christophers in 1907 as invertebrate host of H. canis, although other tick species, such as Haemaphysalis longicornis and Haemaphysalis flava in Japan (Murata et al. 1995) and Amblyomma ovale in Brazil (Forlano et al. 2005), have been identified as possible vectors.
The first report on H. canis came from India and was observed in blood smears of dogs (James 1905), but today the known area of its geographic distribution includes southern Europe, Asia, Africa, South America, and the southeastern United States (McCully et al. 1975, Rajamanickam et al. 1985, Kontos and Koutinas 1990, O'Dwyer et al. 2001). Prevalence data on dogs are still restricted to small areas scattered all over the world and range from 0.9% in France (Criado-Fornelio et al. 2009) to 71% in Croatia (Vojta et al. 2009). The pathological features in the dog range from asymptomatic to severely symptomatic (Marchetti et al. 2009), mainly depending on the level of parasitemia. The animals may have anemia, leucocytosis, intermittent fever, cachexia, purulent conjunctivitis, rhinitis, and less frequently, diarrhea, paraparesis, and necroses in the liver, the spleen, and the bone marrow. The therapy with sulphonamides or dipropionate may be successful. A more severe symptomatology in dogs is usually related to a different species, Hepatozoon americanum, with musculoskeletal involvement (Vincent-Johnson et al. 1997), and is restricted to the United States. The arthropod implicated in the transmission of this species is Ambylomma maculatum, the Gulf Coast tick (Mathew et al. 1998).
From Italy there are some detailed case reports of symptomatic dogs (Arru et al. 1982, Rosati 1986, Bettini 1988, Eleni et al. 1994, Gavazza et al. 2003, Marchetti et al. 2009, Sasanelli et al. 2009), but very few data on the prevalence of H. canis in dog populations (Cassini et al. 2009) and no data on other intermediate hosts. In the framework of a survey on fox parasites (Calderini et al. 2009) and on piroplasmosis in domestic and wild vertebrate hosts (Tampieri et al. 2008, Cassini et al. 2009), we detected H. canis in the animals living in central Italy. The aims of this article are to report on the presence of H. canis in Vulpes vulpes and in ticks, to characterize the parasite by the genetic point of view, and to study its genetic relationship with the strains found in other European countries.
Materials and Methods
The study was performed on 119 foxes shot by legal hunting during population control governmental programs in the years 2005 and 2006. The animals were brought to the “Istituto Zooprofilattico del Lazio e della Toscana” for pathological and parasitological examinations. Spleen tissues were partly preserved in 70% ethanol and partly used to make impression smears. All animals were collected in three areas of the Central Apennines located at 0–2180 m above sea level, within the latitudes 42°16′–43°20′N and the longitudes 11°20′–13°7′E. In the same area, 118 and 172 ticks were, respectively, picked off from dogs and collected from the environment by the dragging woolen blanket technique (Mannelli et al. 1999). The arthropods were identified using the common identification keys (Iori et al. 2005) and then stored in glass tubes containing 70% ethanol.
The spleens of all 119 foxes were screened for H. canis by microscopy: Giemsa-stained touch preparations of three different parts of each spleen were roughly examined by microscopy for a total of 357 slides and part of them (the slides prepared from 56 animals) were discarded because of tissue decomposition. So, only 63 spleens were suitable for morphological examination.
Ticks and spleen tissues were screened by molecular diagnostics (polymerase chain reaction [PCR] and sequencing) for the presence of Hepatozoon. Genomic DNA from spleen samples was extracted using a kit produced by Promega (Wizard SV Genomic DNA Purification®), whereas from tick specimens by means of the phenol–chloroform method applied to each single adult specimen or to pools of nymphs (5 specimens each) or larvae (10 specimens each) (Favia et al. 2001).
To evidence the presence of Hepatozoon DNA, the PCR was performed using the primers CRYPTO F (5′-AACCTGGTTGATCCTGCCAGT-3′) (Herwaldt et al. 2003) and RLB-R2 (5′-CTAAGAATTTCACCTCTGACAGT-3′) (Centeno-Lima et al. 2003) that amplify a fragment of approximately 800 bp of the 18S piroplasm rRNA. The protocol applied is the same we used to evidence piroplasm species (Tampieri et al. 2008). DNA extracted from the blood of a dog found positive to H. canis (Cassini et al. 2009) and distilled water were used, respectively, as templates for positive and negative controls. Amplification products were electrophoresed in a 1.5% agarose gel with 5 μg/mL ethidium bromide. Amplicons were purified (Sure Clean Kit; Bioline) and then sequenced (MWG Biotech). In addition, positive samples were also tested with HEP-1 (5′-CGCGAAATTACCCAATTCTA-3′) and HEP-4 (5′-TAAGGTGCTGAAGGAGTCGTTTAT-3′), which amplify a 670 bp portion of the 18S rRNA gene of Hepatozoon spp. (Criado-Fornelio et al. 2006).
All sequences for both foxes and ticks were assembled and corrected by visual analysis of the electropherogram using Bioedit v.7.0.2 (Hall 1999). Using the BLAST program, they were compared with those of foxes from other European countries (Spain DQ439541, Spain AY150067, Slovakia DQ869309) available in GenBank and with the sequence previously reported by us in Italian dogs (FJ608736). The 18S rRNA sequence for Babesia microti (FJ480420) was included as a outgroup for the Hepatozoon tree. To ensure open-reading frames, and to exclude the presence of any pseudogene, each nucleotide sequence was conceptually translated into amino acid sequences and aligned using MEGA 4.0 (Tamura et al. 2007) and a neighbor-joining tree was constructed. The robustness of the tree was evaluated by the bootstrapping method with 1000 replicates using MEGA 4.0.
Statistical analysis
The results concerning the three areas were compared by means of the chi-squared and the Fisher's exact tests.
Results
The spleen slides showed intracellular bodies, faint blue in color and elliptical in shape (8–12 μm × 3–6 μm). Many of the bodies had a nucleus (round or semilunar), deep blue-stained and usually located at one end of the elliptical body (Fig. 1). The classical “wheel spoke” structure of the meronts producing micromerozoites was also observed. These bodies were regarded as belonging to the genus Hepatozoon. The microscopic examination of the spleens showed 4 H. canis-positive foxes out of 63 (6.3%) animals screened. The results of these four spleens were later confirmed by molecular diagnostics, which evidenced 16/119 (13.4%) foxes positive to H. canis. Table 1 summarizes the molecular results by area, showing that the lowest infection rate was observed in the vicinity of Viterbo (11.5%) and the highest in that of Siena (16.7%), whereas the area of Rieti was very close to the mean value (13.5%).
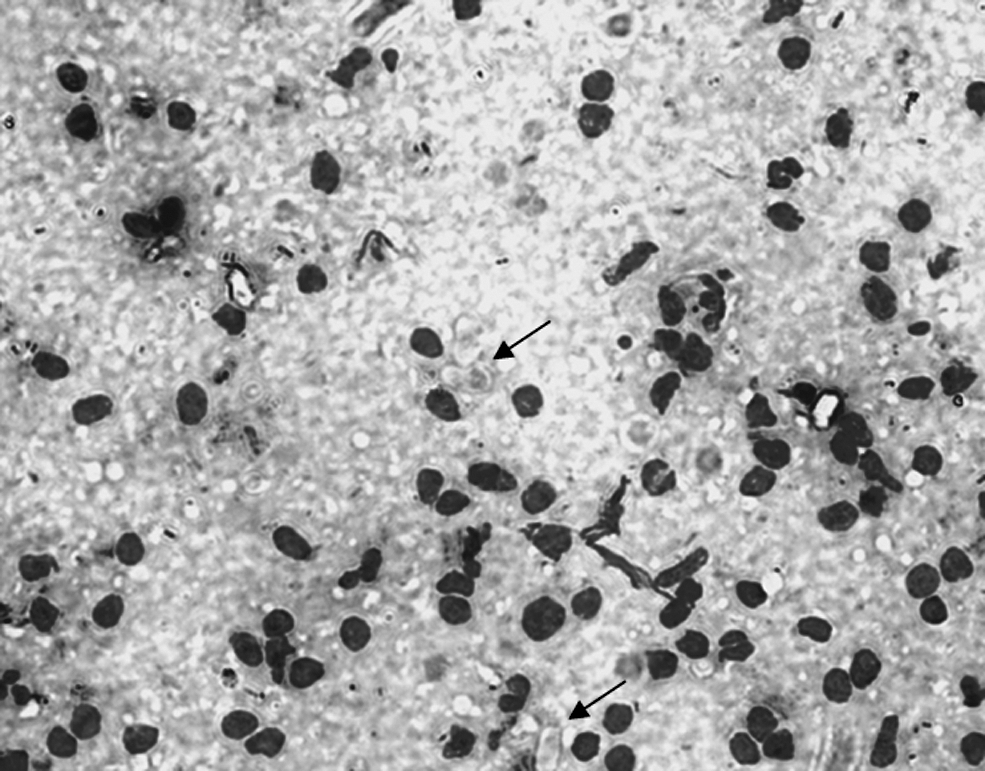
Gametocytes of Hepatozoon canis (arrows) on a spleen impression smear (× 400).
A total of 290 ticks were collected and identified as follows: Dermacentor marginatus (n = 6), Haemaphysalis punctata (n = 6), Hyalomma marginatum (n = 11), Ixodes ricinus (n = 143), Rhipicephalus bursa (n = 9), R. sanguineus (n = 68), and Rhipicephalus turanicus (n = 47). Table 2 summarizes these results and shows that six adult females were PCR positive: two R. sanguineus picked off from dogs, and three R. sanguineus and one I. ricinus collected from the environment.
Sequence analysis indicated that seven isolates (4546, 4530, 1509, 4548, 4552, 6735, and 4241) cluster with the strain we detected in Italian dogs (FJ608736) (99.5–99.7% of identity), whereas other nine isolates (1441, 4156, 339, 4434, 1448, 1442, 1458, 923, and 4242) are closer to the strains isolated in foxes in Spain (AY150067 and DQ439541) and Slovakia (DQ869309) (identity 99.3–99.7%), because their identity with the dog strain (FJ608736) is 98.8–99.3% (Fig. 2).

Neighbor-joining phylogenetic tree from 18S rRNA sequences of H. canis strains identified in Italian foxes (*), dogs, and ticks, and that reported in foxes examined in other European countries.
The strain of H. canis found in the specimens of R. sanguineus picked off from H. canis-positive dogs is closely related to the strain we found in dogs (99.5%). The strains detected in R. sanguineus and I. ricinus collected from the environment show 99.0% identity with the aforementioned Spanish (AY150067) and Slovak (DQ869309) fox strains and are less related to the Italian strain found in dogs (98.4% identity).
Discussion
To our knowledge, this is the first report on H. canis in foxes in Italy, detected by molecular tools and evidenced by microscopy. The occurrence of H. canis infection of the ticks has been already published in a paper concerning dog parasites (Cassini et al. 2009). Microscopic observation of Hepatozoon spp. in blood smears or other biological samples of V. vulpes has been reported in France, Portugal, and Japan (Rioux et al. 1964, Maede and Ohsugi 1982, Conceicao-Silva et al. 1988). Serologic and molecular examination evidenced the presence of the parasite in foxes in Israel, Spain, Poland, and Slovakia (Fishman et al. 2004, Criado-Fornelio et al. 2006, Majláthová et al. 2007, Gimenez et al. 2009).
Comparing the results of microscopy on the 63 foxes with the results of molecular diagnostics on the same 63 animals gives, respectively, 4 (6.3%) and 11 (17%) positive animals (almost the threefold amount of infected animals), confirming thus the higher sensitivity of the molecular diagnostic techniques. Nevertheless, the morphologic demonstration of the occurrence of a pathogen is up to now still the golden standard of the available diagnostic tools.
In this study, the molecular screening revealed a high prevalence of positive foxes (13.4%) on a great number of animals (n = 119) scattered in three wide areas, excluding by far the possibility of vertical transmission of the parasite. The differences in infection rates observed in the areas are not significant (p = 0.81); however, environmental and ecological conditions of the province of Siena seem to support the parasite circulation more efficiently, probably because of the climate that favors a long-lasting activity season of the arthropod hosts.
From the same study area the prevalence (4.3%) we observed in a dog sample similar in size (n = 115) (Cassini et al. 2009) is lower than the prevalence we found in foxes (13.4%). This is probably related to differences in food intake, because foxes feed on live prey or carrion more often than dogs and this may raise the probability of ingesting arthropods accidentally. Our findings demonstrate a greater presence of hepatozoonosis in Italy than previously believed and this requires broader epizootiological investigations to assess its real impact on animal health.
The molecular screening of tick specimens collected in the studied area confirms the importance of the vector R. sanguineus (5 ticks positive to H. canis out of the 68 collected), but suggests that I. ricinus might also be implicated in parasite transmission (1 tick out of the 143 sampled). This hypothesis has to be supported by additional examinations on natural and experimental infections. In fact, if other tick species will turn out to be competent vectors, the presence of H. canis recently evidenced in regions considered R. sanguineus-free (Majláthová et al. 2007) could be explained.
From the taxonomic point of view the parasite strains we found in foxes partly cluster with that we found in dogs from the same area and partly with isolates found in foxes from Spain and Slovakia. These findings support the evidence of discrete genetic heterogeneity among the organisms belonging to H. canis species that infect Italian and other European animals (Vojta et al. 2009); the possible relationship of different parasite strains with pathological manifestations in the hosts deserves further examinations.
The genetic screening of the parasite has just begun, and the application of molecular diagnostics to the strains reported exclusively on morphological basis in Canis aureus (Shamir et al. 2001), Canis mesomelas (Macintire et al. 1997), Lycaon pictus (Shkap et al. 1994), Crocuta crocuta, Acinonyx jubatus, Panthera pardus, and Panthera leo (McCully et al. 1975) could define the real prevalence and assess the range of genetic variability of the species.
Finally, red foxes have peridomestic habits, which make them a likely source of infection for hematophagous arthropods that may be ingested by dogs. The increasing urbanization, with the degradation of wild areas and the intensified contact between wild and domestic animals, coupled with the increase of global temperature is expected to amplify the impact of all vector-borne parasitic diseases (hepatozoonosis included) and enhance the transmission of the parasite. In fact, the increase of temperature shortens the developing time of the parasites, stimulate the increase of the arthropod population density, and protract their activity period (Cancrini and Gabrielli 2009). Moreover, it makes areas, formerly too cold to be colonized, suitable for the arthropods, encouraging in this way their migration and their geographical spreading. All these changes make new areas suitable for vector-borne diseases, as hypothesized by the Geographical Information System (GIS) models (Otranto et al. 2009) and this has been demonstrated by the recent detection of animal and human dirofilariosis in several north-eastern European regions never reported as endemic for this parasite so far (Genchi 2009).
Footnotes
Acknowledgments
The authors are grateful to Miss Eleonora Mangifesta and Miss Enrica Liberatoscioli for their excellent technical assistance. This study was partially supported by grants from the Italian Ministry of Health.
Disclosure Statement
No competing financial interests exist.