
Abstract
A total of 191,244 mosquitoes from 24 species were collected in the Yucatan Peninsula of Mexico from January to December 2008, and tested for the presence of cytopathic virus by virus isolation in Vero cells. Eighteen virus isolates were obtained, all of which were orthobunyaviruses. These were identified by reverse transcription-polymerase chain reaction (RT-PCR) and nucleotide sequencing as Cache Valley virus (n = 17) and South River virus (n = 1). A subset (n = 20,124) of Culex quinquefasciatus collected throughout the year was further tested by RT-PCR using flavivirus-specific primers. Flavivirus RNA was present in this mosquito species year-round. The overall flavivirus minimal infection rate, expressed as the number of positive mosquito pools per 1000 mosquitoes tested, was 7.7 and the monthly flavivirus minimal infection rates ranged from 4.3 to 16.6. Approximately one-third of the RT-PCR products were sequenced and all corresponded to Culex flavivirus, a recently discovered insect-specific flavivirus.
Introduction
Comprehensive surveillance for West Nile virus (WNV) and other mosquito-transmitted viruses has been performed in the Yucatan Peninsula of Mexico since 2001 (Farfan-Ale et al. 2004, Farfan-Ale et al. 2006, 2009, Lorono-Pino et al. 2003, 2004, 2009). Recently, we reported data from entomologic studies performed in the states of Yucatan and Quintana Roo from January to December 2007 (Farfan-Ale et al. 2009). In this study, 96,687 mosquitoes were collected and assayed by virus isolation in mammalian cells. Three isolates were obtained: one was Cache Valley virus (CVV; Orthobunyavirus), another was Kairi virus (Orthobunyavirus), and the identity of the third was not determined. A subset of mosquitoes (210 pools of Culex quinquefasciatus) was further tested by reverse transcription-polymerase chain reaction (RT-PCR) using flavivirus-specific primers. Of these, 145 (69%) pools contained RNA to Culex flavivirus (CxFV), an insect-specific virus first isolated from Culex spp. mosquitoes in Japan and Indonesia (Hoshino et al. 2007), and later from Culex spp. mosquitoes in Guatemala, Mexico, Trinidad, the United States, and Uganda (Morales-Betoulle et al. 2008, Blitvich et al. 2009, Cook et al. 2009, Farfan-Ale et al. 2009, Kim et al. 2009). Another pool contained RNA to a novel flavivirus (designated T'Ho virus) and that is genetically equidistant to several other flaviviruses including WNV (Farfan-Ale et al. 2009).
Here, we report the findings from our mosquito-based virus surveillance performed in the Yucatan Peninsula of Mexico from January to December 2008. The purpose of this study was to increase our knowledge of the biodiversity and distribution of the different species of mosquitoes in the Yucatan Peninsula of Mexico, to detect and isolate new and previously described arboviruses from different mosquito species, and to genetically and phylogenetically characterize these viruses.
Materials and Methods
Description of study sites
Study sites were located in five areas: Merida (urban), Tixkokob (semi-urban), and Celestun (semi-urban) in Yucatan State, and Sian Ka'an (rural) and Isla Mujeres (semi-urban) in Quintana Roo State. The study area in Merida covered approximately 40 km2; all other study areas were less than 4 km2. There were 15 study sites located in Merida, and 5 of these were in the Parque del Centenario Zoologico (the Merida zoo). The Global Positioning System (GPS) locations of the study sites in Merida range from 20°59′25′′ N to 20°55′32′′ N, and 89°35′42′′ W to 89°38′22′′ W. Two study sites were located in Tixkokob (GPS location 20°59′38′′ N and 89°23′14′′ W), an area used for horse and cattle production. In Celestun (GPS location 20°51′7′′ N and 90°23′24′′ W), eight study sites were established. Celestun is a small coastal town and many species of birds overwinter in this area. Eleven study sites were located on Sian Ka'an (GPS location 20°17′54′′ N and 87°15′22′′ W), a small island 3 km off the east coast of the Mexican mainland. The island forms part of the Sian Ka'an Biosphere Reserve. In Isla Mujeres (GPS location 21°12′51′′ N and 86°43′24′′ W), seven study sites were established. Isla Mujeres is a small island town 7 km off the east coast of the Mexican mainland. All study areas used for the mosquito collections have a tropical climate.
Sampling methods and trapping schedules
Mosquitoes were sampled using Mosquito Magnet™ (American Biophysics Corp., North kingstown, RI) traps and Centers for Disease Control and Prevention backpack-mounted aspirators as previously described (Farfan-Ale et al. 2009). In Merida, collections were made from January to December 2008. Each site was trapped 5 days a week every week. One mosquito magnet and/or two backpack-mounted aspirators were used at each site. Tixkokob was visited from January to March. Every site was trapped 5 days a week every other week using one mosquito magnet or two backpack-mounted aspirators. The other locations were visited once (Celestun from December 2–5; Isla Mujeres from November 10–14, and Sian Ka'an from December 16–19). Each site was trapped daily using one mosquito magnet. Mosquitoes from Yucatan State were transported live to the Universidad Autonoma de Yucatan (UADY), frozen at −70°C, and then identified on chill tables according to species and sex using morphological characteristics. Mosquitoes from Quintana Roo State were anesthetized using triethylamine (Kramer et al. 1990), identified, placed into cryostorage vials, and transported in liquid nitrogen to the UADY. Mosquitoes were transported on dry ice from the UADY to Iowa State University.
Mosquito homogenization and virus isolation in Vero cells
Mosquitoes were homogenized in CO2-independent cell culture medium (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum and various other supplements as previously described (Farfan-Ale et al. 2009). Homogenates were centrifuged (10,000 g, 10 min, 4°C), and supernatants were collected. An aliquot of each supernatant was tested for the presence of cytopathic virus by virus isolation in African Green Monkey Kidney (Vero) cells as previously described (Farfan-Ale et al. 2009).
Virus identification by RT-PCR sequencing
Total RNA was extracted from cell culture supernatants and mosquito homogenates using the QIAamp viral RNA extraction kit (Qiagen, Valencia, CA) and analyzed by RT-PCR using primers specific for flaviviruses, orthobunyaviruses, and alphaviruses. The flavivirus-specific primers, FU2 and cFD3, target a 845-nt region of the NS5 gene (Kuno et al. 1998). The orthobunyavirus-specific primers, BCS82C and BCS332V, target a 251-nt region of the small (S) RNA segment (Kuno et al. 1996). The alphavirus-specific primers, VIR966 and VIR966c, target a 98-nt region of the nsP1 gene (Eshoo et al. 2007). Previously described primers specific for WNV (Lanciotti et al. 2000) and T'Ho virus (Farfan-Ale et al. 2009) were also used. Complementary DNA was generated using Superscript III reverse transcriptase (Invitrogen) and PCRs were performed using Taq polymerase (Invitrogen). RT-PCR products were purified using the Purelink Gel Extraction Kit (Invitrogen) and sequenced using a 3730x1 DNA sequencer (Applied Biosystems, Foster City, CA).
Results
Mosquito collections
A total of 191,244 mosquitoes were collected in the Yucatan Peninsula of Mexico from January to December 2008 (Table 1). Of these, 151,851 (79%) were identified as females and 39,393 (21%) were identified as males. The mosquitoes represent 11 genera and 24 species. The most common species was Ochlerotatus (Aedes) taeniorhynchus, which made up 59% of the total sample population, followed by Cx. quinquefasciatus (34%). The majority (82%) of mosquitoes was collected using Mosquito Magnet traps; the remainder (18%) was collected by manual aspiration (data not shown).
Collections were made at multiple sites in five study areas: Celestun, Merida, and Tixkokob in Yucatan State, and Sian Ka'an and Isla Mujeres in Quintana Roo State (Table 2). Most time and effort was devoted to the collection of mosquitoes in Merida, particularly the Merida zoo, because WNV and T'Ho virus activity had previously been detected in this area (Farfan-Ale et al. 2006, 2009). A total of 53,410 mosquitoes were collected in the Merida zoo and another 96,901 mosquitoes were collected at other sites in Merida. Together, these mosquitoes made up 79% of the total sample population. The collections from the Merida zoo consisted of nine species, and the most common species was Cx. quinquefasciatus; it represented 85% of the mosquitoes collected in this site. Sixteen species were collected elsewhere in Merida, and the most common (94%) species was Oc. taeniorhynchus. Six species were collected in Celestun, and the majority (96%) were Oc. taeniorhynchus. In Tixkokob, seven species were collected, and the most common (95%) was Cx. quinquefasciatus. Species diversity was greatest in Sian Ka'an; 20 species were collected in this area, and the most common (37%) was Anopheles vestitipennis. Eight species were collected in the Isla Mujeres, and Cx. quinquefasciatus was the most common (65%).
Does not include mosquitoes collected in the Merida zoo.
Virus isolations in Vero cells
Eighteen of 6009 mosquito pools caused virus-like cytopathic effect (CPE) in Vero cells. All 18 pools consisted of female Oc. taeniorhynchus collected at four study sites in Celestun in December 2008. The sites are within 9 km (6 miles) of each other. Collections were made using Mosquito Magnet traps (13 pools) and backpack-mounted aspirators (5 pools). Seventeen isolates were identified by RT-PCR sequencing as CVV (GenBank accession numbers GU018033–GU018049), and the other as South River virus (SORV; GenBank accession number GU018050). The CVV and SORV minimal infection rates (MIRs) in Oc. taeniorhynchus in Celestun were 0.98 and 0.06, respectively.
Sequence analysis of the CVV small genomic RNA segment
The complete S RNA genome segments of five CVV isolates from Celestun were sequenced, with the exception of an estimated 22-nt at both the 5′ and 3′ termini, using three pairs of overlapping primers (primer sequences are available upon request). As with other members of the genus Orthobunyavirus, the S RNA segment of CVV encodes for the nucleocapsid and NSS proteins in overlapping reading frames (Dunn et al. 1994, Schmaljohn and Nichol 2007). Additional sequencing was also performed on a CVV isolate collected in Merida in 2007 (designated CVV-Mex07) (Farfan-Ale et al. 2009). The nucleotide sequences of the S RNA segments of the five isolates from Celestun were aligned using the CLUSTAL W algorithm (version 2) (Higgins and Sharp 1988, Larkin et al. 2007) and shown to be 98.7% to 100% identical. All but 12 of the 906 nucleotide positions were strictly conserved between all of these isolates. Pairwise alignments of these sequences to the homologous region of the prototype CVV strain 6V633 (GenBank accession number X73465), which is the only CVV isolate for which complete S RNA genome sequence data are available, revealed 91.1% to 97.8% identity. The deduced amino acid sequences of the nucleocapsid proteins of the five CVV isolates from Celestun are 100% identical to one another, as are the deduced amino acid sequences of the NSS proteins. Pairwise alignments of the deduced amino acid sequences of the nucleocapsid and NSS proteins of each isolate to that of the prototype strain revealed 99.1% and 100% identity, respectively, in all cases.
Phylogenetic analysis of the CVV small genomic RNA segment
A phylogenetic tree was constructed with Bayesian methods using a 795-nt region of the S RNA segments of the 5 CVV isolates from Celestun and 19 other orthobunyaviruses from the Bunyamwera serogroup (Fig. 1). Three other CVV isolates are included in this analysis: the prototype strain, CVV-Mex07, and an isolate from Wisconsin in 2003 (designated strain WI-03BS7669); these are the only other CVV isolates that have had most, if not all, of their S RNA segments sequenced. Phylogenetic trees were also generated using neighbor-joining, maximum parsimony, and maximum likelihood methods. In the Bayesian tree, the Celestun isolates of CVV share a close phylogenetic relationship with each other and with the other CVV isolates; the bootstrap supports for these topological arrangements are 92% and 100%, respectively (Fig. 1). Phylogenetically, the S segments of the CVV isolates are most closely related to the homologous region of Potosi virus. The bootstrap support for this topological arrangement is 79%. The topologic arrangements of the neighbor-joining, maximum parsimony, and maximum likelihood trees were similar to the Bayesian tree (data not shown).
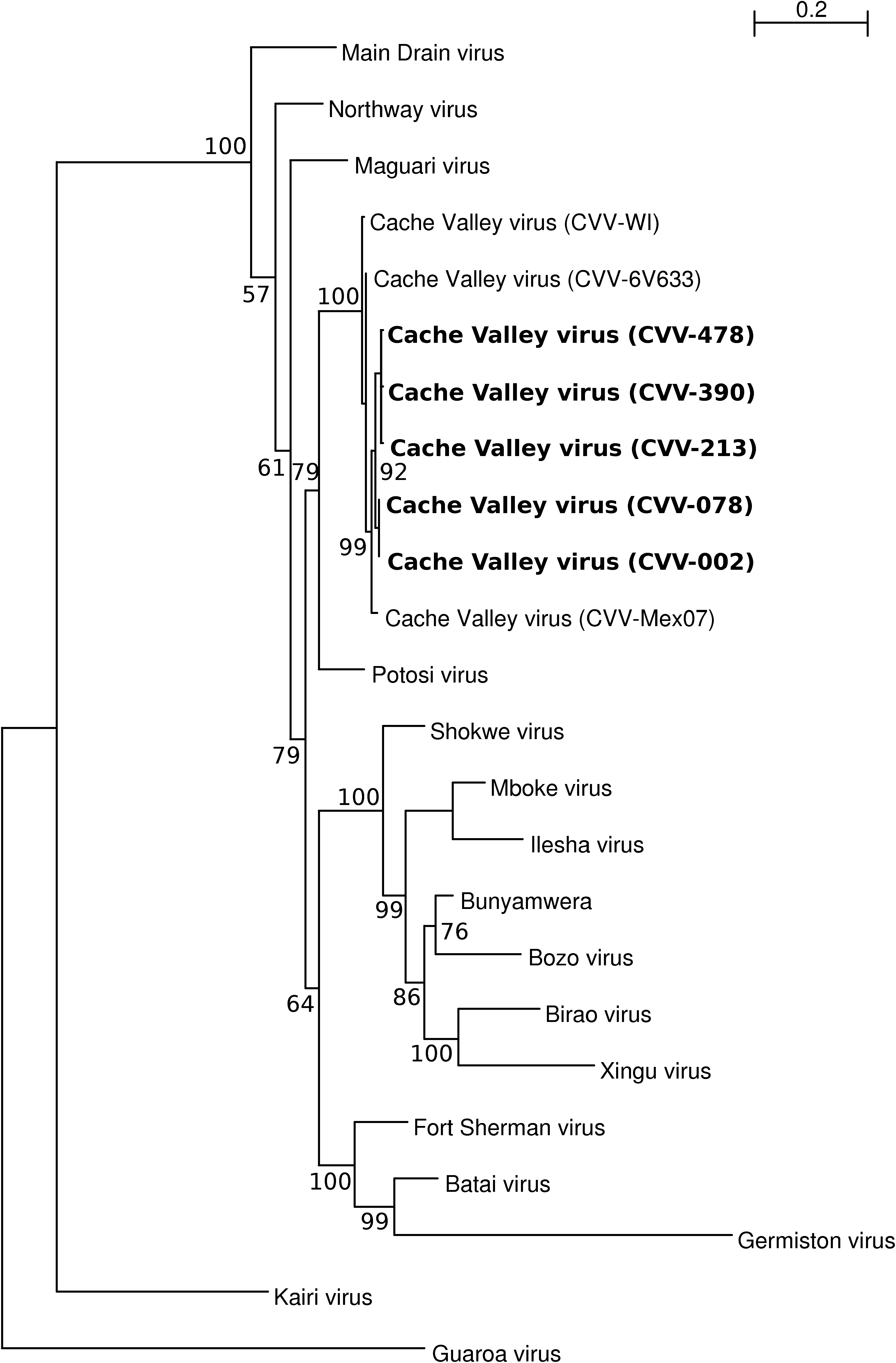
Phylogenetic analysis of a 795 nucleotide region of the small RNA genome segment of 5 CVV isolates obtained in this study and 19 other bunyavirus isolates. The displayed phylogeny was estimated by the program MRBAYES, version 3.1 (Ronquist and Huelsenbeck 2003). Posterior support (out of 100) for selected branches is indicated. An unrooted tree was inferred but is shown rooted using the midpoint method. CVV isolates obtained in the present study are denoted in bold. GenBank accession numbers for sequences used in the phylogenetic analysis are as follows: Batai virus (X73464.1), Birao virus (AM711131.1), Bozo virus (AM711132.1), Bunyamwera virus (D00353.1), CVV strain 6V633 (X73465), CVV strain CVV-002 (GU018033), CVV strain CVV-078 (GU018034), CVV strain CVV-213 (GU018035), CVV strain CVV-390 (GU018036), CVV strain CVV-478 (GU018037), CVV strain CVV-Mex07 (EU879062), CVV strain WI-03BS7669 (DQ315775), Fort Sherman virus (EU564829.1), Germiston virus (M19420.1), Guaroa virus (X73466.1), Ilesha virus (AM709780.1), Kairi virus (EU879063), Maguari virus (M28380.1), Main Drain virus (X73469.1), Mboke virus (AY593727.1), Northway virus (X73470.1), Potosi virus (AY729652.1), Shokwe virus (EU564831.1), and Xingu virus (EU564830.1). CVV, Cache Valley virus.
Detection of CxFV RNA in mosquito homogenates
A subset of mosquito homogenates was directly tested by RT-PCR using flavivirus-specific primers. Overall, 839 pools consisting of 23,561 mosquitoes from 8 species were tested: Aedes aegypti (n = 638; 52 pools), Anopheles albimanus (n = 474; 39 pools), An. vestitipennis (n = 379; 19 pools), Cx. quinquefasciatus (n = 20,124; 600 pools), Ochlerotatus trivittatus (n = 1294; 50 pools), Oc. taeniorhynchus (n = 408; 36 pools), Mansonia titillans (n = 89; 33 pools), and Psorophora cyanescens (n = 155; 10 pools). The Cx. quinquefasciatus used for this study were collected throughout the year (50 pools per month); all were from the Merida zoo, and approximately equal numbers of females and males were used. Aedes, Anopheles, Mansonia, Ochlerotatus, and Psorophora spp. mosquitoes were collected primarily from August to December; all were from Merida and Sian Ka'an, and most were female. Some of the Mn. titillans used in this analysis were collected in 2007 (Farfan-Ale et al. 2009).
Flavivirus RNA was detected in 155 (26%) pools of Cx. quinquefasciatus (Table 3) but in none of the Aedes, Anopheles, Mansonia, Ochlerotatus, and Psorophora spp. mosquitoes. Cx. quinquefasciatus with flavivirus RNA were collected each month. The overall flavivirus MIR in Cx. quinquefasciatus was 7.7. The highest monthly flavivirus MIRs occurred in December and August (16.6 and 12.4, respectively), and the lowest monthly flavivirus MIRs occurred in May to July (4.3–4.6). The annual flavivirus MIRs in female and male Cx. quinquefasciatus were similar (8.3 and 7.2, respectively; data not shown). A subset (n = 50) of RT-PCR products were sequenced using the flavivirus-specific forward primer, and all are >96% identical to the homologous region of CxFV-Mex07, the prototype Yucatan strain of CxFV (Farfan-Ale et al. 2009). Weak bands of approximately 800 nt were observed after the RT-PCR amplification and gel electrophoresis analysis of 19 Oc. trivittatus samples, but nucleotide sequencing revealed that these RT-PCR products were derived from the mosquito 28S ribosomal gene. One of these sequences has been entered into the GenBank database (GenBank accession number GU018051). Lastly, all of the female Cx. quinquefasciatus collected in the Merida zoo from June to December were tested by RT-PCR using a pooled suspension of T'Ho and WNV-specific primers; all were negative (data not shown).
A total of 600 pools (50 per month) were tested.
Discussion
Entomologic surveillance for mosquito-transmitted viruses was conducted in the Yucatan Peninsula of Mexico from January to December 2008. A total of 191,244 mosquitoes in 6009 pools were tested for cytopathic virus by virus isolation in mammalian cells. Eighteen isolates were obtained; 17 were identified as CVV and the other as SORV. CVV is an orthobunyavirus in the Bunyamwera serogroup that is distributed across Canada, Mexico, and the United States (Calisher et al. 1986, Aguirre et al. 1992, Schmaljohn and Nichol 2007, Pabbaraju et al. 2009). CVV has been responsible for two documented cases of human disease, both of which occurred in the United States, as well as disease and pregnancy loss in domestic ruminants (Sexton et al. 1997, de la Concha-Bermejillo 2003, Campbell et al. 2006). Because CVV was isolated from mosquitoes in Merida in 2007 (Farfan-Ale et al. 2009) and Celestun in 2008 (this study), we speculate that this virus is common in the Yucatan Peninsula of Mexico. There is limited genetic and phylogenetic diversity between CVV isolates from Mexico and other geographical locations. SORV is an orthobunyavirus in the California serogroup that was first isolated in New Jersey in 1960 (Sudia et al. 1971) and later in Pennsylvania (Wills et al. 1974) and Georgia (D.G. Mead, pers. comm.). It is not known whether SORV is a pathogen of humans or animals. This is the first report of SORV in Latin America.
One virus was isolated for every 10,625 mosquitoes tested, and thus the proportion of mosquitoes containing cytopathic virus was low. Similar findings were observed in our entomologic investigations in the Yucatan Peninsula of Mexico in 2007 (1 virus per 32,229 mosquitoes) (Farfan-Ale et al. 2009). It is surprising that WNV was not detected in any of these collections, especially those from Merida, where trapping was performed year-round and where serologic studies have provided evidence of previous WNV transmission (Farfan-Ale et al. 2004, 2006, Lorono-Pino et al. 2003, 2009). These findings could indicate that WNV is maintained in the Yucatan Peninsula of Mexico in temporary, dynamic foci. These findings could also be due to a decrease in the intensity of WNV transmission in the Yucatan Peninsula of Mexico since the introduction of this virus into the region. For example, a fourfold decrease in WNV infection rates occurred in Culex tarsalis in a high transmission zone in Colorado from 2003 (MIR 34.5) to 2004 (MIR 8.7) (Bolling et al. 2007). A small number of WNV isolates have been obtained from Latin America and the Caribbean relative to the number of isolations made in the United States. The only regions in Latin America and the Caribbean from where multiple isolations of WNV have been made are Puerto Rico (Barrera et al. 2008) and Guatemala (M.E. Morales-Betoulle, pers. comm.). In both studies, sentinel chicken surveillance was used to monitor ongoing or recent WNV transmission, and mosquito trapping was immediately initiated at sites where seroconversions were identified. Thus, sentinel chicken surveillance followed by targeted entomological surveillance should accompany ongoing mosquito-based surveillance in regions where WNV transmission is not accompanied by widespread human, equine, and avian illness to increase the likelihood of obtaining WNV isolates. However, it is important to note that the inclusion of sentinel chicken surveillance should not detract from the importance of performing ongoing mosquito-based surveillance because the latter is widely regarded to be the primary tool for quantifying the intensity of WNV transmission in an area (Centers for Disease Control and Prevention 2003). T'Ho virus was not detected in any mosquitoes collected in this study despite the considerable amount of time and effort devoted to the collection of mosquitoes in and around the Merida zoo. This finding indicates that T'Ho virus may also be maintained in the Yucatan Peninsula of Mexico in temporary, dynamic foci or that there are notable fluctuations in the intensity of T'Ho virus transmission in this region each year. It is not known whether sentinel chicken surveillance followed by targeted entomological surveillance provides a suitable approach to detect T'Ho virus because the vertebrate hosts of this virus have not been identified.
The flavivirus MIR in Cx. quinquefasciatus collected in the Merida zoo from January to December was 7.7. Nucleotide sequencing data indicate that most (if not all) of these mosquito homogenates contain CxFV RNA. The CxFV sequences were not entered into the GenBank database because the sequencing reactions were performed in one direction using a single primer (FU2). However, a comprehensive sequencing study is currently being performed in our laboratories using CxFV isolates from Mexico. The CxFV MIR (10.9) in Cx. quinquefasciatus in the Merida zoo from June to August 2007 was also high (Farfan-Ale et al. 2009). The presence of CxFV RNA in Cx. quinquefasciatus collected each month suggests that this virus is active in the Yucatan Peninsula of Mexico year-round. In contrast, CxFV was detected in Culex pipiens in Iowa from July through October, but not in any mosquitoes collected in May or June (Blitvich et al. 2009). In Texas, CxFV was identified in Cx. quinquefasciatus and/or Culex restuans from November to March, but in none of the mosquitoes collected in April to August (Kim et al. 2009). These data could indicate that CxFV activity is seasonal in certain areas or that persistence varies between different Culex spp. mosquitoes. The variations in monthly flavivirus MIRs (4.3–16.6) in Cx. quinquefasciatus in Merida may be due to the reasonably small number of mosquitoes tested (50 pools per month). The identification of CxFV RNA in approximately equal numbers of male and female mosquitoes suggests that CxFV is maintained in nature by vertical transmission.
CxFV was not detected in any Aedes, Anopheles, Mansonia, Ochlerotatus, and Psorophora spp. mosquitoes. These findings were not unexpected since all previous isolations of CxFV have been made from Culex spp. mosquitoes (Cx. pipiens, Cx. quinquefasciatus, Cx. restuans, Cx. tarsalis, and Cx. tritaeniorhynchus) (Hoshino et al. 2007, Morales-Betoulle et al. 2008, Blitvich et al. 2009, Cook et al. 2009, Farfan-Ale et al. 2009, Kim et al. 2009). However, other insect-specific flaviviruses have been identified in non-Culex mosquitoes. For instance, Kamiti River virus was isolated from Aedes macintoshi collected in Kenya in 1999 (Sang et al. 2003, Cook et al. 2006). More recently, Aedes flavivirus was detected in Aedes albopictus and Aedes flavopictus in Japan (Hoshino et al. 2009), and Nakiwogo virus was detected in Mansonia africana nigerrima in Uganda (Cook et al. 2009). This study provides no evidence that insect-specific flaviviruses, other than CxFV, are present in Mexico although it is important to note the small numbers of non-Culex spp. mosquitoes tested by RT-PCR for flavivirus RNA.
In summary, we report the detection of bunyaviruses (CVV and SORV) and flaviviruses (CxFV) in mosquitoes in the Yucatan Peninsula of Mexico. Due to the enormous impact that arboviruses have on human and animal health and the sparse number of comprehensive studies that have explored the diversity of viruses in mosquito populations in Mexico, additional entomologic investigations are required in this region.
Footnotes
Acknowledgments
The authors thank the field workers from the UADY. These individuals are as follows: Carlos Baak, Mildred López, Carlos Estrella, Alex Ic, Roger Arana, Wilberth Chi, Hugo Valenzuela, Iván Villanueva, Jesús Miss, Rosa Cetina, Lourdes Talavera, and Roger López. This study was supported by grant 5R21AI067281-02 from the National Institutes of Health.
Disclosure Statement
No completing financial interests exist.