
Abstract
Arboviruses are frequently associated with outbreaks in humans and represent a serious public health problem. Among the Brazilian arboviruses, Mayaro virus, Dengue virus (DENV), Yellow Fever virus, Rocio virus, Saint Louis Encephalitis virus (SLEV), and Oropouche virus are responsible for most of human cases. All these arboviruses usually produce undistinguishable acute febrile illness, especially in the acute phase of infection. In this study we investigated the presence of arboviruses in sera of 519 patients presenting acute febrile illness, during a dengue outbreak in São José do Rio Preto City (São Paulo, Brazil). A multiplex-nested RT-polymerase chain reaction assay was applied to detect and identify the main Brazilian arboviruses (Flavivirus, Alphavirus, and Orthobunyavirus genera). The molecular analysis showed that 365 samples were positive to DENV-3, 5 to DENV-2, and 8 to SLEV. Among the positive samples, one coinfection was detected between DENV-2 and DENV-3. The phylogenetic analysis of the SLEV envelope gene indicated that the virus circulating in city is related to lineage V strains. These results indicated that during that large DENV-3 outbreak in 2006, different arboviruses cocirculated causing human disease. Thus, it is necessary to have an efficient surveillance system to control the dissemination of these arboviruses in the population.
Introduction
DENV is considered the most important arbovirus that infects humans. DENV, serotypes 1–4, are responsible for large urban outbreaks, especially when the cocirculation of different serotypes is observed or when a new serotype is introduced (Figueiredo 2000, Gubler 2002, De Simone et al. 2004, Fonseca and Figueiredo 2005). Since 1986, large dengue epidemics have been reported in Brazil, initially after introduction of DENV-1 in 1985, followed by introduction of DENV-2 in 1990. In 2000, DENV-3 was introduced in Rio de Janeiro State. DENV-3 and DENV-2 have been responsible for most cases of dengue fever (DF) and dengue hemorrhagic fever and/or dengue shock syndrome in Brazil (Nogueira et al. 2005, Vasconcelos et al. 2005, 2009). Recently, the reintroduction of DENV-4 was reported in Manaus, north region of Brazil (Figueiredo et al. 2008).
DF is characterized initially by abrupt high fever (39–40°C) followed by headache, myalgia, arthralgia, prostration, eye and abdominal pain, rash, and positive tourniquet test during illness for 5–7 days basically (Brazilian Ministry of Health 2005). World Health Organization defines dengue hemorrhagic fever as the manifestation of high fever, hemorrhagic phenomena with hepatomegaly, and signs of circulatory failure. The evolution to hypovolemic shock and plasma leakage is characterized as dengue shock syndrome and can be fatal to patient (WHO 1997)
In São José do Rio Preto City (northwest region of São Paulo State, Brazil), cases of DENV-1 were initially identified after 1990 through 1995 (Mondini et al. 2005). In 1996, DENV-2 became the most important predominant agent of dengue (Brazilian Ministry of Health 2008), and in 2005 and 2006, São José do Rio Preto City suffered from a large DENV-3 outbreak with more than 15,000 reported cases (Mondini et al. 2009).
In this way, the objective of this study was to identify Brazilian arboviruses that caused acute febrile illness in patients from São José do Rio Preto (SP), which were considered positive for DENV-3 during dengue outbreak according to epidemiological criteria (legality of epidemics and consistent clinical symptoms) by the Municipal Office of Health and Hygiene and by the Center of Epidemiological Surveillance of the city.
Materials and Methods
Clinical samples
We analyzed 519 clinical samples, collected consecutively from May to July, 2006, from patients who presented acute febrile disease for ≤5 days. They were attended by the São José do Rio Preto Health Service and the illness was diagnosed as dengue based only on clinical–epidemiological data (clinical signs compatible with dengue and occurrence of an outbreak) according to Brazilian Ministry of Health (2009). These patients were attended in Basic Unities of Health and Hospitals, where information on clinical presentation of the disease was collected and later registered in the National System of Injury of Notifications—SINAN. All patients were investigated by Public Health Office and they were considered autochthones cases. Blood samples were collected; the sera were separated and stored at −80°C. Viral RNA was extracted from 140 μL of each serum with the QIAamp Viral RNA Mini kit (Qiagen) as described by the manufacturer and RNAs were submitted to polymerase chain reaction (PCR) test. This study was approved by the Internal Ethical Review Board and all samples were in storage (at −80°C) when the study was initiated.
Study strategy
As different arboviruses cause similar diseases, clinical samples were initially tested to check the presence of Flavivirus and Alphavirus. First, Flavivirus and Alphavirus genus-specific primers, targeting target NS5 and nsP1 regions, respectively, were used simultaneously in a Duplex-reverse transcription (RT)-PCR (D-RT-PCR) assay (Fig. 1). Second, DENV 1–3, YFV, and SLEV species-specific primers were used, because those viruses are known to circulate in that region. In this way, a Multiplex-Nested-PCR (M-N-PCR) was used to identify DENV 1–3 and YFV and another N-PCR was used to detect SLEV. After those initial PCRs, only negative samples were tested for other arboviruses. Different N-PCRs were used to detect DENV-4, ROCV, Ihéus virus, Bussaquara virus, Iguape virus, and West Nile virus (WNV). Another M-N-PCR was used to detect Venezuelan Equine Encephalitis virus, Eastern Equine Encephalitis virus, Western Equine Encephalitis virus, and MAYV. Finally, a RT-PCR followed by a semi-nested assay (SN-PCR), using specific primers to S segment of OROV, was used. All primers used in this study are listed in Table 1. Precautions to avoid contamination were followed, positive and negative controls were used in all reactions, and the procedure was reproduced several times (Borst et al. 2004).

Strategies for arbovirus detection in sera samples from patients in São José do Rio Preto. DENV 1–4, dengue virus serotypes 1 to 4; YFV, Yellow Fever virus; SLEV, Saint Louis Encephalitis virus; ROCV, Rocio virus; IGUV, Iguape virus; ILHV, Ilhéus virus; WNV, West Nile virus; BSQV, Bussuquara virus; MAYV, Mayaro virus; VEEV, Venezuelan Equine Encephalitis virus; WEEV, Western Equine Encephalitis virus; EEEV, Eastern Equine Encephalitis virus; OROV, Oropouche virus.
(+), genomic sense primer; (−), primer antisense.
Degenerated primers. Single letter code: Y (C or T); S (C or G); R (A or G); H (A or C or T); W (A or T); N (A or C or G or T); K (G or T); D (A or G or T).
With primer cM3W.
With primer cM3W based on virus sequence from GenBank (NWEE AF214040; NMAY U94602).
With primer FG1.
With primer FG1 based on virus sequence from GenBank (NDENV 1 AB074760; NDENV 2 AF38403; NDENV 3 AY923865; NDEN 4 NC_002640; NWN DQ411034).
Primers to S segment of OROV.
Pb, pair of bases.
Virus and RNA extraction
The viral strains used as positive controls were MAYV BeAr20290, DENV-1 Mochizuki, DENV-2 SpH 125367, DENV-3 RPDen06/41, DENV-4 Boa Vista, SLEV BeH 355964, Ilhéus virus BeH7445, ROCV SpH34675, West Nile NY99, YFV 17DD (vaccine), and OROV BeAn 19991. They were propagated by intracerebral inoculation into 2-day-old suckling mice or in C6/36 Aedes albopictus cell cultures as previously described (Shope and Sather 1979, Figueiredo 1990). Viral RNAs were extracted from 140 μL of a 1/20 dilution of mouse brain tissue macerated suspensions or from cell culture supernatants with the QIAamp Viral RNA Mini kit (Qiagen) according to the manufacturer's instructions.
PCR assays for Alphavirus and Flavivirus detection
D-RT-PCR, M-N-PCR, and N-PCR were performed as previously described (de Morais Bronzoni et al. 2005). For the D-RT-PCR, the RT mixture was incubated at 50°C for 50 min and at 70°C for 15 min to inactivate the reverse transcriptase. The thermal cycling was performed with termocycler GeneAmp® PCR System 9700 (Applied Biosystems). According to the authors, M-N-PCR presented 99% of sensitivity and 83% of specificity for the arbovirus.
SN-RT-PCR assays for OROV identification
The RT-PCR and SN-PCR were performed as described (Terzian et al. 2009). Primers used were c2ORO (RT-PCR) and s1ORO (PCR). Primers s1ORO and c1ORO were used together in the SN-PCR. The PCR mixture and cycling conditions were performed as for M-N-PCR Flavivirus assays (“PCR assays for Alphavirus and Flavivirus detection” section).
Viral isolation
Viral isolation was performed in C6/36 cell culture as previously described (Figueiredo 1990) in selected samples to isolate a new SLEV strain. C6/36 cell culture confluent monolayer was infected, for 60 min, with 100 μL of sera samples diluted 1/10 in culture medium. After incubation, 5 mL of Leibovitz medium L-15 (Cultilab, Campinas, Brazil) with 1% bovine fetal serum (Cultilab) was added to cell monolayer. After 7 days, the cell culture was frozen at −80°C and thawed and the supernatant was analyzed for viral RNA presence by the specific PCR assay described earlier (Bronzoni et al. 2004, de Morais Bronzoni et al. 2005).
Nucleotide sequence
Amplicons obtained by (M)-N-PCR were purified with ethanol as previously described (Sambrook 2001) and then sequenced by dideoxynucleotide method (Sanger et al. 1977). Sequencing was realized using Big Dye Terminator Kit v3.1 (Applied Biosystems) according to the manufacturer's instructions. In the sequencing PCR reactions, species-specific primers (3.2 μM) were used. DNA obtained was centrifuged for 20 min (4°C, full speed), precipitated with 80 μL of 75% isopropanol, and homogenized with 2.0 μL of 0.5 M ethylenediaminetetraacetic acid (pH 8.0) buffer and Bleu Dextran. DNA was denaturated at 95°C for 2 min and applied on sequencing gel. Electrophoresis was realized on ABI PRISM 377 DNA Sequencer (Applied Biosystems). Sequences were analyzed using DS Gene 2.0 (Accelrys) and BLAST (
Analysis of partial sequence of envelope gene from a SLEV isolate
Attempting to identify the genotype of SLEV isolate, the ORF that codes for the envelope (E) protein was partially amplified and then sequenced. RNA obtained from the SLEV-isolated sample was submitted to RT-PCR and N-PCR assays using the primers previously described (Chandler and Nordoff 1999, Kramer and Chandler 2001) and listed in Table 1. The amplicon was cloned into pCR2.1 vector (TA Cloning Kit; Invitrogen) and then several clones were sequenced using Big Dye Terminator Kit v3.1 (Applied Biosystems), according to manufacturer's instructions. Sequences were analyzed using DS Gene 2.0 (Accelrys). Nucleotide and deduced amino acid sequences were used to perform a similarity search in sequence databases, using FASTA (
and (M) genotyping according to Auguste et al. (2008) and May et al. (2008), respectively, and both based on the Saint Louis Encephalitis virus envelope gene sequence; (B)genotyping according to Baillie et al. (2008), based on the whole genome.
According to Auguste et al. (2008), this strain belongs to genotype IV.
Genotype determined in this study.
WNV, West Nile virus; MVEV, Murray Valley Encephalitis virus; JEV, Japanese Encephalitis virus.
Statistical analysis
For statistical analysis, t, F, and χ 2 (comparison of proportion) test using Epi Info™ 6.4 from the Center of Disease Control and Yates test were used. Standard deviations were used for mean, variance, and deviation standard.
Results
Arbovirus detection by PCR
From 519 clinical samples that were analyzed, arboviruses genomes were detected in 374 samples (72.07%). DENV-3 genome was detected in 365 samples and SLEV and DENV-2 genomes were detected in eight and five samples, respectively. Among these, coinfection of DENV-3 and DENV-2 was detected in one sample (Fig. 2). The remaining 145 samples of febrile acute illness patients (27. 93%) were all negative for tested arboviruses. All positive amplicons were sequenced to confirm the specificity of the amplification. Viral isolation was performed in SLEV-positive samples using the same sera samples from PCR assay. As result, one SLEV strain was isolated (SJRP06/155).
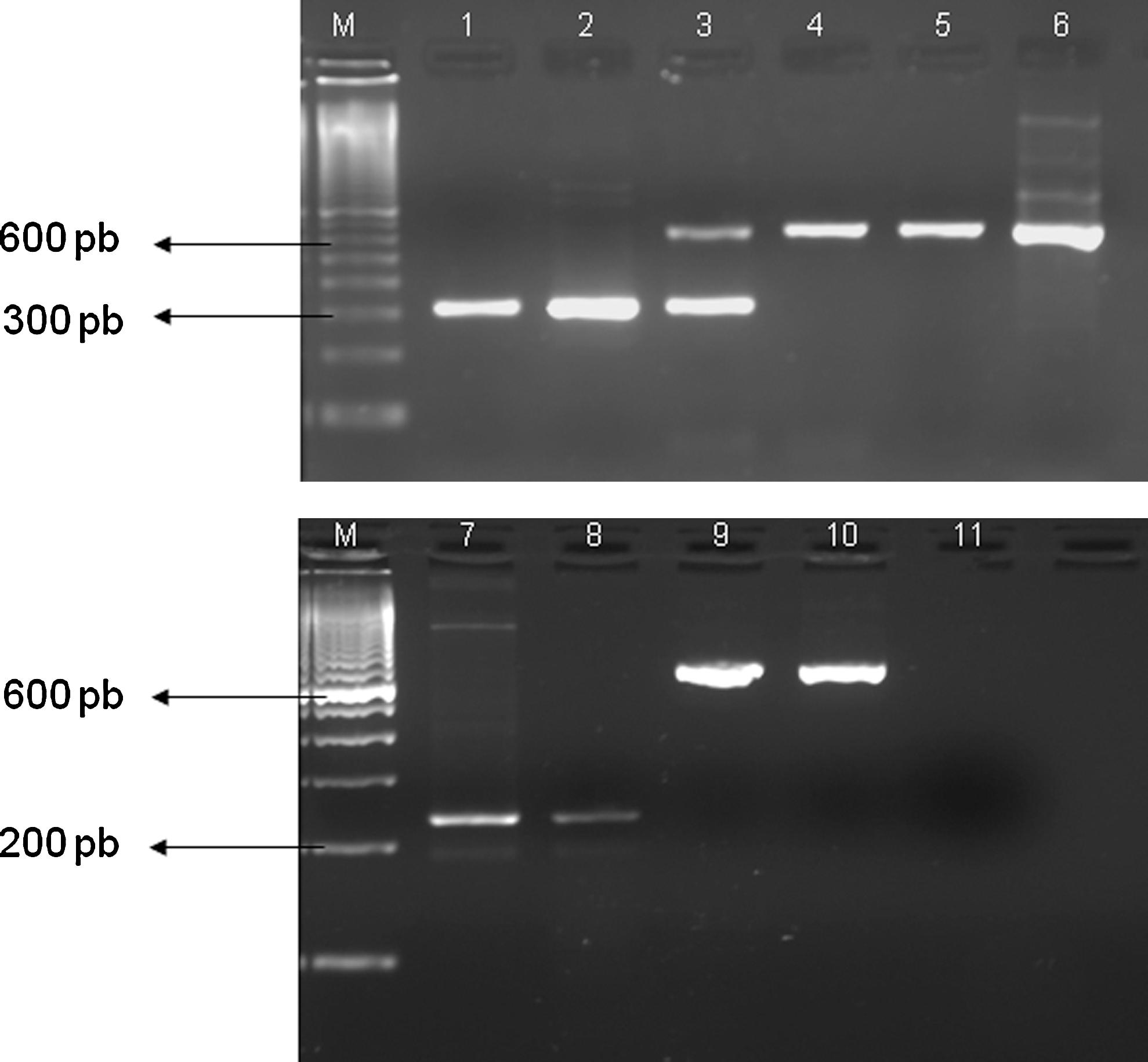
Agarose gel (2%) electrophoresis of amplicons from multiplex-nested-PCR (M-N-PCR) for Flavivirus detection from clinical samples of São José do Rio Preto patient. Line 1: DENV-2-positive control; line 2: DENV-2-positive clinical samples; line 3: coinfection by DENV-2 and 3 in clinical samples; lines 4, 5, and 9: DENV-3-positive clinical samples; lines 6 and 10: DENV-3-positive control; line7: SLEV-positive control; line 8: SLEV-positive clinical sample; line 11: negative control. M, molecular size marker (DNA ladder, 100 pb). For lines 1 to 6, 9, and 10, FG1, NDEN1, NDEN2, NDEN3, and NYF primers in multiplex system were used. For lines 7 and 8, primers FG1 and NSLE were used. Multiplex N-PCR and N-PCR mixture was subjected to 25 cycles of 94°C for 1 min, 53°C for 1 min, and 72°C for 2 min. A final extension step was carried out at 72°C for 5 min. PCR, polymerase chain reaction.
Epidemiological and clinical findings
This study was realized with 519 clinical samples collected at the beginning of an outbreak indicating high positivity to Flaviviruses. DENV-2, DENV-3, and SLEV were detected in 374 samples.
The epidemiological and clinical data showed no statistically significant difference between the Flavivirus-positive patients by PCR and the negatives, with the exception on the tourniquet test (Table 3), which was found more frequently in Flavivirus-positive patients as expected (p > 0.005). Only one case presenting a positive tourniquet test was not dengue but it was a SLEV infection. Pleural and pericardic hemorrhage, hepatomegaly, cardiomyopathy, and shock were not referred by nonarbovirus cases. No other considerable symptoms were referred. One case of nasal bleeding (0.68%) and one case of pain on inferior limb and lumbar region (0.68%) from negative cases were referred. Among negative patients for Flavivirus, 6.90% of them were hospitalized; however, no statistically significant difference was found compared with positive cases (p-value: 0.666461). No significant difference (p-value: 0.77) was observed on fever duration (3.125 days mean) in arbovirus-infected (3.125 days mean) and uninfected individuals (3.054 days mean).
Yates test.
Insufficient n for proportion comparation (n < 5).
Based on RT-PCR assay, 145 individuals were negative for arboviruses; however, based on Ministry of Health clinical criteria (SMSH/VE/SEI/SINAN) in this same group, 101 cases (69.65%) were classified as having DF and 1 case was classified as having atypical severe dengue.
Analysis of E gene from a SLEV isolate
The E gene of the SLEV isolate (strain SJRP06/155) was partially amplified and sequenced generating a 492 pb nucleotide sequence. An initial search for similar nucleotide sequences in nucleotide databases was performed using FASTA, which performs the search by global alignments of sequences. Partial envelope nucleotide sequence presented higher similarity of values 90.2–98.0% when compared with other SLEV sequences. Similarity values below 69.7% were observed when the SJRP06/155 E gene sequence was compared with other sequences from virus belonging to Japanese Encephalitis group, such as WNV, Japanese Encephalitis virus, and Murray Valley Encephalitis virus. When the SJRP06/155 E gene was translated and compared with protein database, SJRP06/155 sequence also presented higher similarity and identities values when compared with SLEV sequences than when compared with other Flaviviruses. These results once more confirmed that SJRP06/155 is a SLEV isolate.
Envelope nucleotide sequences from different SLEV genotypes were retrieved from GenBank and aligned with SJRP06/155 sequence (Table 2). When the deduced amino acid of SJRP06/155 envelope sequence was compared with other deduced amino acid sequences of different SLEV strains (Table 2), two unique amino acid changes were observed in the SJRP06/155 sequence. At position 44, SJRP06/155 had a glutamic acid residue, whereas all other sequences presented a residue of lysine. At position 88, SJRP06/155 presented a leucine residue, whereas all other studied sequences presented a residue of proline (with exception of ARG79 that presented a residue of serine). No amino acid residues specific to lineage II or III (previously defined) were observed in the deduced SPJR06/155 partial envelope amino acid sequence. A residue of asparagine at position 66, previously described as specific for virus belonging to lineage VII, was observed in the deduced SJRP06/155 amino acid sequence and in sequences of ARG-67 and ARG-66. The specific amino acid residues observed for lineages III to V are not located at the analyzed portion of the deduced SJRP06/155 amino acid sequence (Fig. 3).

Analysis of envelope (E) gene and deduced amino acid sequence of SLEV. (
Phylogenetic trees were reconstructed based on 411 nt of E gene (corresponding to nucleotides 963–1373 of complete genome sequence of strain Kern217, NC_007580). Maximum likelihood (data not shown) and distance methods (Fig. 3) gave similar results. The seven SLEV lineages were observed and SJRP06/155 clustered with strains from genotype V, including some Brazilian isolates such as BRA73A, BRA-UB, BRA-73B, BRA-71, and BRA-72 from the Brazilian Amazon region.
Discussion
DENV has been responsible for large urban outbreaks generally associated with introduction of new serotypes (Figueiredo 2000, Gubler 2002, De Simone et al. 2004). Since 2004, DENV-3 was considered as the only serotype circulating in São Paulo State, differently of previous years, when different serotypes were concomitantly detected (Boletim Epidemiológico Paulista 2005). In this study, 72.07% of the studied patients had arboviruses genome in the serum, where most of the cases (97.6%) were positive to DENV-3. However, DENV-2 was detected in five cases, which was also detected in São José do Rio Preto in the previous years (Mondini et al. personal communication), corroborating our results. Besides, in 2007, DENV-2 was isolated in 20% of dengue cases that occurred in Brazil, and until April 2008, São Paulo State registered 60% of dengue cases caused by this serotype (Brazilian Ministry of Health 2007, 2008).
A simultaneous DENV-2/DENV-3 infection was detected in one patient. Coinfection cases of DENV-3 and DENV-2 have been previously reported in Somalia, Taiwan, China, and Brazil (Kanesa-thasan et al. 1994, Wang et al. 2003, Wenming et al. 2005, Araujo et al. 2006). Other coinfection cases had also been reported among DENV serotypes (Gubler et al. 1985, Laille et al. 1991, Wang et al. 2003, Araujo et al. 2006, Bharaj et al. 2008). It is believed that these coinfections only occur during outbreaks with circulation of multiple serotypes and where there is high prevalence of the urban vector, being capable to transmit more than one virus at the same time (Wenming et al. 2005, Bharaj et al. 2008).
All studied patients had been previously considered as having dengue based on clinical and epidemiological criteria. However, from 519 samples tested by RT-PCR, 27.93% were negative to dengue as well as other tested arboviruses. Many other infectious agents, such as poliomyelitis, rabies, measles, rubella, hepatitis, malaria, or leptospirosis, can induce similar symptomatology (Fonseca and Figueiredo 2005, Vasconcelos et al. 2005), and the laboratory diagnosis is fundamental in this context. This work shows that the utilization of clinical and epidemiological criteria for dengue diagnosis can be a major problem, even in the presence of a major outbreak. The samples collected for this study were all collected in less than 5 days of the onset of the disease, suggesting that a false negative due to absence of viruses circulating in the blood is not the point here.
SLEV is widely distributed in the Americas, from Canada to Argentina (Sabattini et al. 1998, Burke and Monath 2001). SLEV infections have been reported in the United States and less frequently in Central and South America (Sabattini et al. 1998). However, 5% of population in north and southeast of Brazil seemed to have antibodies against the virus. Nevertheless, these data must be carefully analyzed, because it may be due to cross-reaction between antibodies of different Flavivirus, which are induced especially by Yellow Fever vaccination or exposure to DENV. In this way, SLEV may be circulating in these areas and causing human infections, although most of them are undiagnosed (Lopes et al. 1979, Vasconcelos et al. 1998, Figueiredo 2000).
In Brazil, in 2004, SLEV was isolated from an acute febrile illness patient living in São Pedro county, São Paulo State (Rocco et al. 2005). In 2006, a serological study in birds carried out in the southern coast of São Paulo State showed a high increase of the prevalence of antibodies against SLEV (Suzuki et al. 2006). Finally, also in 2006, concomitant to a DENV-3 outbreak in São José do Rio Preto, 12 dengue suspected cases had SLEV genome detected in clinical samples and it has been considered the first SLEV outbreak registered in Brazil (Mondini et al. 2007b). Serology is an important tool in SLEV monitoring, especially for bird reservoirs and horses. It is known that horses can be infected but they rarely develop symptoms. Studies with infected horses can indicate virus circulating among animals, and in the presence of mosquitoes (Culex vectors), they can be a potential source for human infection. In 2009, researches of São Paulo State, using a SLEV IgG-ELISA, detected a 31.11% of positivity among 180 studied horses. This indicated that human infections can be occurring in locations where these animals live (Silva et al. 2009).
The fact that SLEV was the agent of human disease serves as advice to health professionals about the need of a complete clinical and epidemiological investigation about febrile diseases, especially during outbreaks, as SLEV infections can be unrecognized or be mistaken with dengue because of similar clinical manifestations. Epidemiological investigation associated with the methodology applied in this study, using FG1/FG2 and NSLE primers in a multiplex system with dengue primers and associated alphaviruses PCR, can be an important tool in a trustworthy and fast diagnosis, not only for SLEV but also for other arboviruses. In this study, it was demonstrated that SLEV circulation in São José do Rio Preto was more important than previously reported because eight dengue-suspected cases were diagnosed in the laboratory as having SLEV.
Phylogenetic analysis of different SLEV isolates suggests that this virus is mainly maintained locally, although migration between different areas also occurs (Kramer and Chandler 2001, Twiddy and Holmes 2003). This analysis showed that SLEV isolates are clustered in seven different lineages based on E gene or the genome sequences (Kramer and Chandler 2001, Auguste et al. 2008, Baillie et al. 2008) and it is mainly correlated with the geographical characteristics of those viruses and not phenotypic characteristics such as virulence (Trent et al. 1980, Kramer et al. 1997). Lineages I and II are predominantly from North America, whereas lineages III to VII belong to South and Central Americas. The Brazilian SLEV strains described so far belong to genotypes II (BRA-04), III (BRA-68), and V (BRA-72, BRA-71, BRA-73A, BRA-73B, BRA-73C, BRA-73D, BRA-UA, BRA-UB, and BRA-60) (Auguste et al. 2008, Baillie et al. 2008, May et al. 2008).
Although the clustering of SRJP06/155 within lineage V (also including BRA72, BRA71, BRA73A, BRA73B, BRA-UA, BRA-UB, TRIN-55, PER-75, CA00A, and CA01A) was not supported by high bootstrap values (as also observed by May et al. 2008), similar results were observed by other authors when analyzing bigger sequences of E gene (Auguste et al. 2008, May et al. 2008), where the same strains were clustered within lineage V. In the same way, the strains BRA-73A, BRA-73B, PER-75, and TRI-55 were also shown to belong to lineage V, based on the analysis of the whole genome of SLEV (Baillie et al. 2008). Analysis of other regions of SJRP06/155 genome could help elucidating the genotyping of SJRP06/155.
May et al. (2008) defined some specific amino acid changes that are observed in the E protein sequence for individual lineages. Although it was not possible to check for amino acid residues specific to lineages III to V, no amino acid change specific to lineage II or III was observed in the deduced SPJR06/155 partial envelope amino acid sequence. An amino acid residue specific to lineage VII (at position 66) was observed in SJRP06/155 E protein sequence; however, the residue of glycine observed at position 29 in isolates from genotype VII was not observed in SJRP06/155 sequence.
In summary, we reported here a dengue outbreak where DENV-3 infections predominated, consistent with the introduction of this new serotype in the region. Some infections were caused by DENV-2, the previous causative agent of outbreaks in the region. However, the unexpected presence of SLEV causing acute febrile illness was also detected by an unusual diagnostic approach looking for arboviruses instead of dengue only. These results showed the real situation of cocirculation of some arboviruses during an outbreak, initially caused only for DENV-3, which warns health professionals and requires the inclusion of infection caused by SLEV in the differential diagnosis of acute febrile illness and encephalitis. In Brazil, SLEV situation is worrisome because probably the disease has been unrecognized or mistaken with dengue and SLEV human cases and is occurring in other places of Brazil. São José do Rio Preto is an important geographic area for arboviruses surveillance (especially Flaviviruses) because of outbreaks caused by DENV-1, −2, and −3, as well as identification of SLEV and the risk Sylvatic Yellow Fever in the region. In this way, it is necessary to implement an active surveillance, based on prevention and control of patients with acute febrile illness, reservoirs, domestics' animals, and mosquito vectors, to avoid introduction or reintroduction of these viruses. We also believe that the use of clinical and epidemiological criteria without laboratory confirmation should not be done even in the presence of a large DENV outbreak.
Author's Contributions
A.C.B.T., L.T.M.F., F.C.N., and M.L.N. conceptualized the study. A.C.B.T. and B.P.F. processed and performed the manipulation of samples. A.M. collected samples and clustered patients' clinical information. R.V.M.B. contributed to technique used for this study and intellectual and technical knowledge. B.P.D. contributed to sequence and phylogenetic analysis. E.M.S.C. contributed to statistical analysis. L.T.M.F. contributed with viral control samples. M.L.N. critically appraised the manuscript and contributed to the intellectual content. All authors helped in drafting and approved the final manuscript. M.L.N. is guarantor of the manuscript.
Footnotes
Acknowledgments
The authors thank all staff of the County Office of Health and Hygiene, Basic Unities of Health, Emergency Unities and Hospitals from the São José do Rio Preto City. They also thank Milena Polloto for sample sequencing. This work was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (grant no. 04/11098-2) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (grant no. 401396/2004-5). This work was also partially supported by the Viral Genetic Diversity Network (VGDN–FAPESP–Brazil).
Disclosure Statement
No competing financial interests exist