
Abstract
Altogether, 430 nymphs and 570 adult Ixodes ricinus ticks were collected in 2006 (n = 506) and 2007 (n = 494) from a forest area in Middle Germany (Thuringia). Single ticks were investigated by polymerase chain reaction and restriction fragment length polymorphism or sequencing for Borrelia spp. (ospA gene) and Babesia spp. (18S rRNA gene). Overall, 27.0% (270/1000) were infected with Borrelia species. Out of these, Borrelia garinii was detected most frequently (133/270)—especially, OspA serotype 6 (51/270), followed by Bo. burgdorferi (70/270), Bo. afzelii (42/270), Bo. valaisiana subgroup I (28/270), not typable Borrelia spp. (5/270), Bo. spielmanii (3/270), Bo. valaisiana subgroup II (2/270), and Bo. lusitaniae (1/270). In 1.4% of investigated ticks mixed infections with several Borrelia spp. occurred. Babesia spp.–specific DNA was detected in 5.0% of ticks. Babesia microti was slightly more prevalent (28/50) than Babesia divergens (20/50). Moreover, 5.9% (16/270) of Borrelia spp.–infected ticks were coinfected with Babesia spp. Knowledge on the degree of heterogeneity of Borrelia species and OspA types is prerequisite not only for local risk assessment, but also for diagnostic test and vaccine development.
Introduction
The aim of our study was to contribute epidemiological data on prevalence of different Borrelia species and OspA types, and on different Babesia spp. for Middle Germany (Thuringia). The investigation on incidence and coincidence, as well as on interactions and the role of different reservoirs of the well-known Borrelia spp. with other tick-borne pathogens, like Babesia spp., is important for risk assessment, diagnosis, and therapeutic and prevention strategies. Since there are no reliable diagnostic tools for Babesia spp. available, diagnosis and treatment of human babesiosis and coinfections with Borrelia spp. are difficult so far.
Material and Methods
The investigation mixed-forest area “Zeitzgrund,” about 700 m2 in size along the main hiking trail, is located in Thuringia. The climate is moderate with an average annual temperature of 10°C. The annual precipitation was 595.6 mm in 2006 and 736.7 mm in 2007. A total of 1000 I. ricinus ticks were collected during the years 2006 (May–October) and 2007 (April–September) by blanket-dragging. Collection was carried out on 2 days (9:00 a.m. to 2:00 p.m.) per month with an average of 60–80 collected ticks per month. Ticks were washed with 70% ethanol and sterile water, crushed with sterile pistils and forceps, and suspended in 200 μL sterile phosphate-buffered saline using separate sterile laboratory instruments for each tick. DNA extraction was carried out further with the High Pure PCR Template Preparation Kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer's instructions. Amplification targeting the ospA gene of Borrelia spp. was performed according to Michel et al. (2004). DNA of the strains PKo (Borrelia afzelii, OspA type 2), PBi (Borrelia garinii, OspA type 4), PKa2 (Borrelia burgdorferi, OspA type 1) [courtesy of Max von Pettenkofer Institute, Munich, Germany], and VS116 (Borrelia valaisiana) [courtesy of Institute for Nutritional Sciences, Jena, Germany] served as positive controls for amplification.
As specific target for detection of Babesia spp., a genus-specific part of the 18S rRNA gene was amplified by an optimized single polymerase chain reaction (PCR) protocol according to Hartelt et al. (2004). All positive amplicons were confirmed in a further PCR targeting another part of the 18S rRNA gene (582 bp) with the primers Bab 18S F3 (5′-CCA GAC ATA GAG AGG ATT G-3′) and Bab 18S R3 (5′-CCT TCC GCA GGT TCA CC-3′). DNA of Ba. microti and Ba divergens (courtesy of Ute Mackenstedt, University of Hohenheim, Oldendorf, Germany) served as a positive control. Negative controls were included in each PCR. The amplified products were separated by electrophoresis in a 2% agarose gel, stained with SYBR-Green (Biozym Diagnostic, Germany), and observed under UV light. To identify Borrelia species and OspA types, all positive amplicons were digested separately with the restriction enzymes Kpn2I, BglII, SspI, HindIII (MBI Fermentas, St. Leon-Rot, Germany), and SfuI (Roche Diagnostics) as described previously (Michel et al. 2004). PCR products were optionally digested with the restriction enzyme XbaI (MBI Fermentas) to distinguish OspA type 6 from OspA type 8 and Bo. valaisiana subgroup II from Borrelia spielmanii (former A14S) (Lenčáková et al. 2006). Only Borrelia spp.–specific amplicons that could not be identified by restriction fragment length polymorphism (RFLP) were sequenced. To confirm Babesia spp.–specific DNA, all positive PCR amplicons were digested with AluI (MBI Fermentas). Additionally, all positive PCR products were sequenced to differentiate between Ba. microti and Ba. divergens. Purification was carried out by Agarose Gel Extraction Kit (Jena Bioscience, Jena, Germany) and afterward used for DNA sequencing. Sequencing reactions were performed using the DYE namic™ ET Dye terminator Cycle Sequencing Kit (GE Healthcare, Munich, Germany). This was followed by preparation for running onto the ABI PRISM 310 genetic analyzer (Applied Biosystems, Darmstadt, Germany), according to the manufacturers' instructions. Sequences were compared to data obtained from the NCBI database. Statistical analyses were performed using the χ 2 test. A p-value of ≤0.05 was considered statistically significant.
Results
Altogether 1000 I. ricinus ticks were investigated: 430 nymphs (2006: n = 206, 2007: n = 227), 293 females (2006: n = 159, 2007: n = 134), and 277 males (2006: n = 141, 2007: n = 136) (Table 1).
n, nymphs; f, female adults; m, male adults.
Headlines of the total number of collected and infected ticks in 2006, 2007, and both years appear in bold.
In 27.0% (270/1000) of the ticks Borrelia spp. DNA was detected (Table 1). The Borrelia spp. infection rate of 32.3% (184/570) in adult ticks was significantly higher (p < 0.001) than that in nymphs (20.0%, 86/430). There seems to be only a slight year-to-year variation in prevalence. Thus, the infection rate in 2006 was 27.1% (137/506) and in 2007, 26.9% (133/494) of ticks were infected with Borrelia spp. Borrelia OspA types were further characterized with the RFLP method. Bo. garinii (133/270; 49.3%) was detected most frequently, followed by Bo. burgdorferi (70/270), Bo. afzelii (42/270), Bo. valaisiana subgroup I (28/270), not typable Borrelia spp. (5/270), Bo. spielmanii (3/270), Bo. valaisiana subgroup II (2/270), and Bo. lusitaniae (1/270) (Fig. 1). Among all identified Bo. garinii we detected the OspA types 3, 4, 5, 6, 7, and 8 with a predominance of type 6 (51/270) and type 5 (37/270; Fig. 2). Altogether 17/270 Borrelia spp. amplicons had to be identified by sequencing. In 12 samples Bo. garinii resulted from sequencing without further OspA type characterization (not identified OspA type of Bo. garinii), and in 5 samples sequencing only allowed the confirmation of Borrelia spp.–specific amplicons (not typable Borrelia spp.). The distribution of species and OspA types did not vary significantly between both years of investigation (p < 0.001). Bo. garinii OspA types 4 and 7, Bo. valaisiana subgroup II, and Bo. lusitaniae were only detected in adult ticks. All three Bo. spielmanii infections occurred in nymphs (Table 2). In 1.4% (14/1000) of the ticks, six adults and eight nymphs, double infections with several Borrelia species were found. Bo. garinii dominated in combination with Bo. burgdorferi, Bo. valaisiana, and Bo. afzelii. Nymphs and adult ticks were involved in these combinations (Table 2). The prevalence of Borrelia species and Bo. garinii OspA types in double-infected ticks was comparable to the distribution in ticks harboring only a single infection.

Borrelia species.
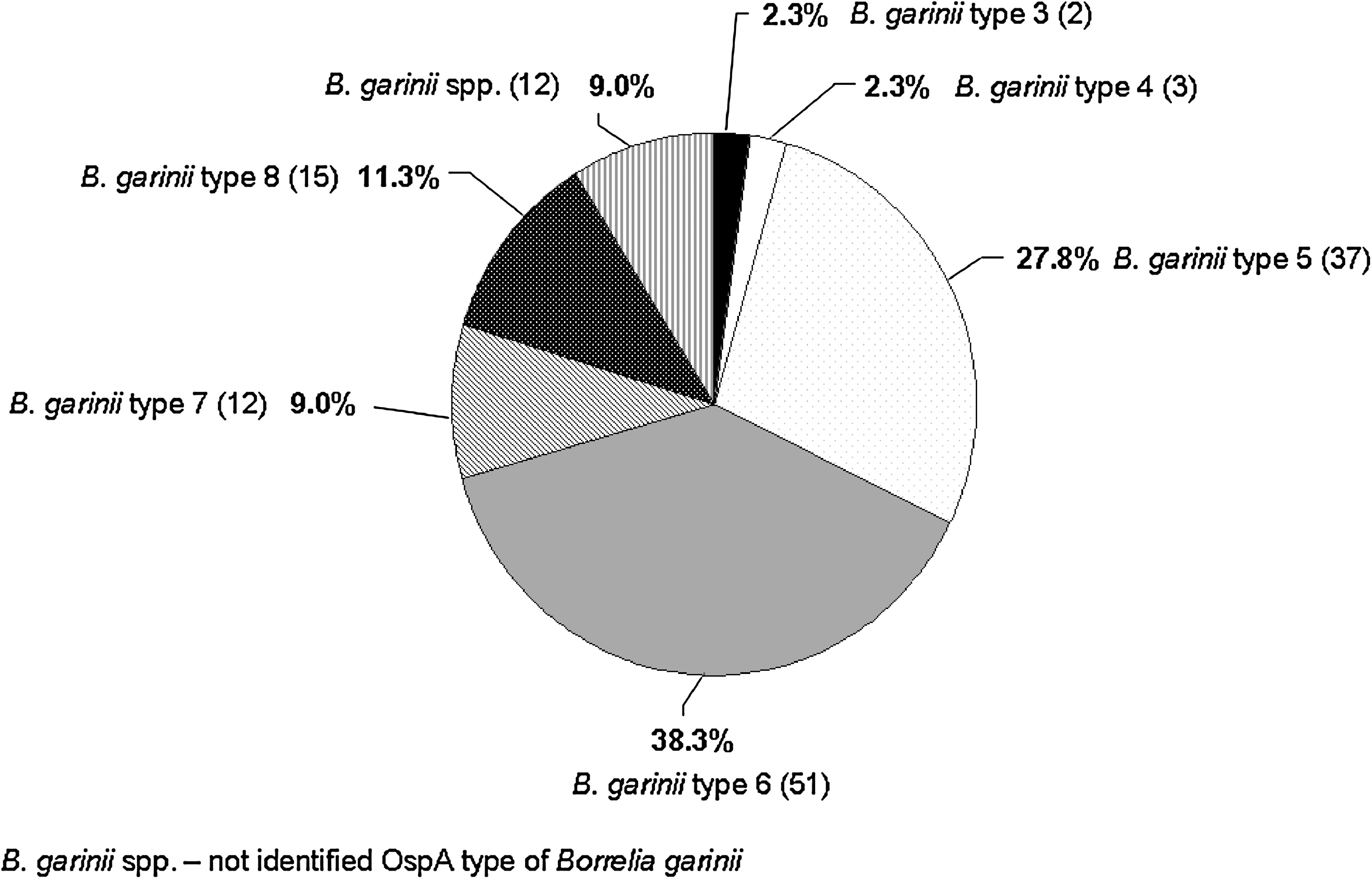
Borrelia garinii OspA types. Bo. garinii spp., not identified OspA type of Bo. garinii.
(): Double infections with several Borrelia spp. Altogether, in 1.4% (14/1000) ticks (6 adults and 8 nymphs), mixed infections with several Borrelia species were detected: 8/14 occurred in 2006 and 6/14 in 2007.
[]: Borrelia spp./Babesia spp. coinfections. In 1.6% (16/1000), coinfections with Borrelia spp. and Babesia spp. were detected: 9 occurred in 2006 and 7 in 2007. Borrelia afzelii, Borrelia garinii (OspA type 4, 5, 6), Borrelia burgdorferi, and Borrelia valaisiana were combined with Babesia microti (10/16) and Babesia divergens (6/16). Only adult ticks were involved in these coinfections.
[*]: One adult tick was infected with Bo. garinii type 5, Bo. burgdorferi, and Ba. microti.
B.b., Borrelia burgdoreri; afz, Borrelia afzelii; gar, Borrelia garinii; n.i., not identified OspA type of Borrelia garinii; val I, Borrelia valaisiana subgroup I; val II, Borrelia valaisiana subgroup II; lus, Borrelia lusitaniae; spiel, Borrelia spielmanii; Borr. spp., not typable Borrelia spp.
Headlines of the years 2006, 2007, and the total numbers appear in bold.
Babesia spp.–specific DNA was detected in 5.0% (50/1000) of the ticks. Infection rate was significantly higher in adults (7.4%, 42/570) than in nymphs (1.9%, 8/430; p < 0.0001; Table 1). Prevalence of Babesia spp.–infected ticks seems to vary between different years of investigation. Thus, the infection rate in 2006 was 7.9% (40/506) and in 2007 only 2.0% (10/494; Table 1). After sequence analyzing of all Babesia spp.–positive amplicons, Ba. microti (28/50) was slightly more prevalent than Ba. divergens (20/50). Two isolates were only characterized as Babesia species.
Borrelia spp. and Babesia spp. coinfections were detected in 1.6% (16/1000) of all ticks examined with an infection rate of 2.4% (7/293) in females and 3.2% (9/277) in males, respectively. Interestingly, these coinfections occurred most frequently during the peak time of Borrelia spp. infection, in July 2006 and May 2007 (Table 1). One female tick was even infected with Ba. microti, Bo. garinii type 5, and Bo. burgdorferi. Only adult ticks were coinfected with Borrelia spp. and Babesia spp. (Table 2).
Discussion
Since different species and OspA types of Lyme borreliosis spirochaetes obviously have a different pathogenic potential, information on their distribution in tick populations is a basic requirement for local risk assessment and for the development of diagnostic tests as well as vaccines based on OspA. This study provides data on the distribution of a broad variety of Borrelia spp. and OspA types and Babesia spp. in free-living I. ricinus ticks in Middle Germany (Thuringia). The prevalence of Borrelia spp. in questing I. ricinus of 27.0% (270/1000), found in this study, is in line with previous reports from Germany, where infection rates in unfed nymphs and adults ranged from 11.1% to 28.7% (Hildebrandt et al. 2003, Fingerle et al. 2008). Most studies throughout Europe recognized Bo. garinii and/or Bo. afzelii as predominant species, whereas Bo. burgdorferi and Bo. valaisiana are less frequent, and detection of Bo. lusitaniae is rare (Hildebrandt et al. 2003, Jouda et al. 2003, Michel et al. 2004, Rauter and Hartung 2005, Richter and Matuschka 2006). Accordingly, Bo. garinii was the predominant species in this study, but surprisingly followed by Bo. burgdorferi. This is in sharp contrast to the above-mentioned studies. Interestingly, in two other studies Bo. burgdorferi was frequently detected as well (Lenčáková et al. 2006, Fingerle et al. 2008). In mixed infections the combination of the predominated species Bo. garinii/Bo. burgdorferi was most prevalent. The same result was reported from Slovakia recently (Lenčáková et al. 2006), whereas another study detected the combination Bo. garinii/Bo. valaisiana mostly (Kurtenbach et al. 2001).
It is suggested that certain Borrelia species seem to be associated with certain vertebrate hosts. Bo. afzelii and some Bo. garinii strains appear to use rodents as main reservoirs, whereas Bo. valaisiana and most Bo. garinii strains were found in enzootic cycles with birds as main vertebrate hosts (Kurtenbach et al. 1998a, 1998b, 2001). Further, these associations seem to be mirrored in the complement sensitivity of these species or strains; that is, rodent-associated strains are resistant to rodent complement but sensitive to bird complement and vice versa. It was therefore proposed that the complement is a key determinant for the host preference of different Borrelia species (Kurtenbach et al. 2002). For Bo. burgdorferi, to our knowledge, such a specific cycle has not been described for Europe so far. This species displays partial resistance to avian and mammalian complement and might be more a generalist (Kurtenbach et al. 2002). In this study Bo. burgdorferi was surprisingly the clearly predominant species in questing I. ricinus nymphs accounting for 28% of all strains found. Notably, a broader variety of Borrelia species and subspecies were present in adult ticks (Table 2). Occurrence of the different Borrelia species may vary considerably between different European countries and even between closely located areas (van Dam et al. 1993, Hubalek and Halouzka 1997, Kurtenbach et al. 2001, Rauter et al. 2002, Michel et al. 2004, Casati et al. 2006, Lenčáková et al. 2006, Fingerle et al. 2008). We found neither a significant difference of Borrelia spp.–infected ticks during the year, nor a difference of infection rate between both years of investigation (p > 0.05) (Table 1), confirming the results described in Germany previously (Hildebrandt et al. 2003, Fingerle et al. 2008). In 17/270 Borrelia spp.–specific amplicons RFLP analysis did not allow clear species characterization. Partly, analysis of the 16S rRNA gene, ospC gene, or flaβ gene could be more sufficient to concretize the Borrelia species in such samples. Our findings demonstrate that almost all relevant Lyme borreliosis spirochaetes described for Europe cocirculate in the investigation area. OspA has been shown to be a promising candidate for an effective vaccine for Europe (Gern et al. 1997). According to our data, all of the known human pathogenic species and Bo. garinii subtypes must therefore be included in the development of vaccines and diagnostic tests. Further information on the distribution of different Borrelia species and subspecies (OspA types) in their natural reservoir hosts and vectors is needed for a better understanding of circulation in natural foci as well as for the establishment of effective preventive strategies against Lyme borreliosis (Wilske et al. 1996a, 1996b; Michel et al. 2004, Fingerle et al. 2008).
In Europe infection rates of Babesia spp. in ticks range from 0.9% to 20% (Skotarczak and Cichocka 2001, Hartelt et al. 2004, Halos et al. 2005, Casati et al. 2006, Piccolin et al. 2006). An Austrian study reported a very high infection rate of Babesia spp. (51%) assuming that the tick density of the investigated area is the determining factor (Blaschitz et al. 2008). Not all studies were able to differentiate between Ba. microti and Ba. divergens, and only few of them included characterization of the Ba. divergens–like species EU1 (Wielinga et al. 2009). Currently, the prevalence of babesiae in ticks in Germany as determined by PCR approximates only 1% (Hunfeld et al. 2002, Hartelt et al. 2004). In this study we detected Babesia spp.–specific DNA in 5.0% (50/1000) of questing ticks. However, it must be considered that the sensitivity of PCR for the detection of various Babesia spp. in the different developmental stages of unfed I. ricinus ticks has analytical limitations, especially in unfed larvae and nymphs (Casati et al. 2006). Moreover, the prevalence of babesiae in ticks can vary substantially even at the local level and from year to year (Foppa et al. 2002). In seroepidemiological investigations, an overall seroprevalence ranging from 1.7% in healthy blood donors to 11.4% in tick-infested individuals in Germany suggests that Babesia spp. infections in humans may occur more frequently than previously assumed (Hunfeld et al. 2002). Although less than 50% of tick bites are noted, individuals doing frequently outdoor activities suffer from several tick bites each year (Wilske et al. 2000). Therefore, a high prevalence of Babesia spp. in ticks and frequent tick exposure are important risk factors for acquiring the pathogen (Hunfeld et al. 2002, Hunfeld and Brade 2004, Hildebrandt et al. 2008). Recently, the first two cases of human babesiosis have been reported in Germany. They occurred in geographic areas where the presence of Babesia spp. in enzootic cycles was obvious for decades without the awareness of the risk of acquiring Babesia spp. infection in humans before (Häselbarth et al. 2007, Hildebrandt et al. 2007).
Coinfection rates of Borrelia spp. and Babesia spp. in questing ticks range from 0.1% in Italy up to 19% on Nantucket Island, MA (Skotarczak and Cichocka 2001, Piccolin et al. 2006, Wójcik-Fatla et al. 2009). In the 1.6% (16/1000) coinfected ticks the human pathogenic Lyme borreliosis spirochaetes Bo. garinii, Bo. afzelii, and Bo. burgdorferi were involved (Tables 1 and 2). Mixed infections and coinfections may result from feeding on different infected hosts or feeding on one host infected with multiple pathogens at the same time. Further, cofeeding transmission, that is, transferring infections directly from one tick to another while feeding together on the same host individual, may result in multiple infections in single ticks (Gern and Rais 1996). As questing adult I. ricinus ticks have a history of two blood meals on diverse hosts (Randolph and Craine 1995), Borrelia spp. and also infections with other pathogens like Babesia spp. may accumulate with the number of infectious blood meals (Kurtenbach et al. 1998a). In our study prevalence of Babesia spp. was significantly higher in adults than in nymphs (p < 0.0001), and coinfections with Borrelia spp. only occurred in adult ticks. Nevertheless, nymphs can also harbor both pathogens even it is detected rarely (Skotarczak et al. 2002, Wójcik-Fatla et al. 2009). In Lyme borreliosis and babesiosis, patients frequently report fatigue and other flu-like symptoms. Symptoms of Borrelia spp./Ba. microti coinfection are more severe and can even be fatal in some cases. Borrelia-specific DNA is much more persistent in the blood of the coinfection-affected patients than in those infected by a single pathogen only (Krause et al. 1996, Skotarczak et al. 2002). The therapy applied to these patients has to differ from that used for those with Lyme disease only (Krause et al. 1996).
Systematic large-scale seroepidemiological and molecular epidemiological surveys are urgently needed to learn more about the distribution and medical relevance of Babesia spp. in Germany and in several parts of Europe. Both pathogens, Borrelia spp. and Babesia spp., should be taken into consideration in patients with unspecific symptoms and history of tick infestation.
Footnotes
Acknowledgments
We want to thank Volker Fingerle and Bettina Wilske (Max von Pettenkofer Institute, Munich, Germany) and Wolfram Dorn (Institute of Nutrition, Friedrich Schiller University, Jena, Germany) for providing the Borrelia-positive control strains PKo (Bo. afzelii, OspA type 2), PBi (Bo. garinii, OspA type 4), PKa2 (Bo. burgdorferi, OspA type 1), and VS116 (Bo. valaisiana); Ute Mackenstedt (University Hohenheim, Germany) for the positive controls of Ba. microti and Ba. divergens; and Gisela Gaschler (Institute of Medical Microbiology, Friedrich Schiller University) for helpful discussions and excellent laboratory support.
Disclosure Statement
No competing financial interests exist.