
Abstract
The increasing interest in Rift Valley fever virus (RVFV) and its potential impact on naive animal populations deserve revisiting experimental reproduction of RVFV infection, particularly in those animal breeds for which no data about their susceptibility to RVFV infection have ever been recorded. In this study we show the susceptibility of 9–10 weeks old European sheep (Ripollesa breed) to RVFV infection, showing a mild, subacute form of disease. Four different viral isolates efficiently replicated in vivo after subcutaneous experimental inoculation, and consistent viral loads in blood and virus shedding (variable in length depending on the RVFV isolate used) were detected, showing horizontal transmission to a noninfected, sentinel lamb. RVFV infection caused transient pyrexia in adult lambs and no other clinical symptoms were observed, with the exception of corneal opacity (“blue eye”) found in 3 out of 16 subcutaneously inoculated sheep. In conclusion, adult sheep from this European breed are readily infected with RVFV without apparent clinical manifestations.
Introduction
Since the first significant epizootic episodes in South Africa in 1951, the disease expanded dramatically to North and East African countries (WHO 2007) and out of Africa, in the Arabian Peninsula (Ahmad 2000). Recently, large RVF outbreaks occurred in East African countries after heavy rainfall season and also in Madagascar, Swaziland, and South Africa (Ahmad 2000, WHO 2007, Sissoko et al. 2009). Because of climatic conditions, facilitating the expansion of mosquito niches (Anyamba et al. 2009), global human and animal trade (Fevre et al. 2006), and long-distance movement of RVF lineages (Bird et al. 2007), RVF is considered an emerging threat for the nonendemic European countries. Among animals, RVFV is spread primarily by the bite of infected mosquitoes, mainly Aedes, Anopheles, and Culex genus, in Africa (Pagès et al. 2009) and also in the Mediterranean area, and thus mosquitoes have been able to act as potential vector of RVFV (Moutailler et al. 2008).
Little is known about the susceptibility of European ruminant species to RVFV infection and even less about their potential to act as natural amplifying hosts of this virus. Despite being an RNA virus, RVFV shows low (∼5%) variability at the nucleotide level (Bird et al. 2007). Differential pathogenicity between isolates has been reported (Swanepoel et al. 1986, Anderson and Peters 1988, Olaleye et al. 1996). In this work, the pathogenicity of four different RVFV field isolates, from ruminant and mosquito origins, in a European sheep breed has been investigated. The potential of RVFV shedding as well as the risk of horizontal transmission to noninfected, in-contact lambs in the absence of competent mosquito vectors was also evaluated. The final aim of this work was to establish and characterize an infection sheep model for the assessment of future RVF vaccine prototypes.
Materials and Methods
Virus and preparation of viral inoculum
The four RVFV isolates used and their passage history during the study are listed in Table 1. Tissue culture infectious dose (TCID50) titers of each virus stock were determined on BHK-21 cells by the method of Reed and Muench (1938).
Chicken embryo related cells.
Mouse brain.
n.a., not available.
Animals
Nineteen lambs (Ripollesa breed) of both sexes, 9–10 weeks old at the time of experimental inoculation, were used. All animals were in-farm treated with insecticide to eliminate ectoparasites and then moved to the BSL3-CReSA's facility at 10 days before the infection. Lambs were fed following normal procedures used in conventional farms, with water supply ad libitum.
Experimental design and clinical sampling
All lambs were housed in the same box. Groups of four lambs were experimentally inoculated with each one of the RVFV isolates used. In all cases, 1 × 105 TCID50 in 1 mL volume were inoculated subcutaneously, behind the elbow. A fifth group of three lambs remained noninoculated, albeit in contact with all the inoculated animals (as sentinels). Clinical signs, including rectal temperature and behavior, were daily recorded for 14 days. At 8 days post-inoculation (dpi), two sheep from each group with negative quantitative retrotranscriptase polymerase chain reaction (qRT-PCR) blood results were euthanized and necropsied. The remaining sheep were sacrificed at 14 dpi.
Nasal and oral swabs were taken daily to track viral shedding collected with 1 mL of Trizol® (Invitrogen) for viral RNA extraction. Blood was also collected daily and processed as follows: 0.2 mL of total blood was lysed with 0.75 mL of Tri® reagent lysis buffer (Sigma) for viral RNA extraction. Serum samples from each animal taken at 0, 4, 8–9, and 14 dpi were analyzed for the presence of anti-RVFV antibodies.
From animals necropsied at 8 and 14 dpi, liver, kidney, lung, spleen, mesenteric lymph node, and inguinal lymph node samples were taken simultaneously paired for biological determinations and histological procedures (fixed in formalin 10%).
One set of samples were immediately mixed with 0.5 mL RNAlater (Ambion), kept overnight at 2–8°C, and stored at −80°C until viral RNA extraction.
Four-micrometer-thick sections of each paraffin-embedded block of studied animals were cut and processed for routine histological procedures (hematoxylin and eosin stain). All paraffin-embedded tissues were evaluated in a blinded fashion by the same pathologist.
RNA extraction and qRT-PCR
RNA from swabs and tissues was extracted using Trizol (Invitrogen) and from blood using Tri-reagent (Sigma), following the manufacturer's instructions. About 0.15 g of each tissue was homogenated in a volume of 0.75 mL of Trizol reagent. RNA was resuspended in 25 μL of nuclease-free water (Roche) and 3 μL was used for the amplification reaction in a total volume of 20 μL. RVFV RNA from samples was detected following a TaqMan one-step qRT-PCR specific for segment L of RVFV in a Fast7500 equipment (Applied Biosystems). The following primer pairs and TaqMan probe were designed previously (Mwaengo et al. unpublished) and synthesized by Tib Molbiol (Berlin, Germany); RVFV-fwd primer 5′-TTCTTTGCTTCTGATACCCTCTGT-3′, RVFV-rev primer 5′-GTTCCACTTCCTTGCATCATCTG-3′, and RVFV Taqman L probe 5′-(FAM)-TTGCACAAGTCCACACAGGCCCCT-(TAMRA)-3′. The amplification conditions were optimized to perform a one-step RT-PCR: 600 nM of RVFV-fwd, 900 nM of RVFV-rev, and 250 nM of RVFV-probe using OneStep RT-PCR Master Mix Reagents (Applied Biosystems) in a final volume of 20 μL. Then, the amplification cycles performed were reverse transcription at 50°C for 20 min, initial denaturing reaction at 95°C for 10 min, and 40 PCR cycles of 95°C for 30 s, 55°C for 1 min, and 72°C for 30 s.
RVFV-RNA quantitation was achieved by using an in vitro translated RVFV L-specific RNA of 135 nucleotides as a standard. Briefly, a 135-bp L gene (positions 2872 to 3006) was PCR amplified with the aforementioned forward and reverse primers and directly subcloned within the T7 polymerase expression pGEMT vector (Mwaengo et al. Unpublished). The complete insert including the upstream T7 promoter was amplified with a vector-specific primer (M13-40: 5′-GTTTTCCCAGTCACGAC-3′) and the RVFV-rev primer using OneStep RT-PCR reagents (Qiagen). The PCR product was in vitro transcribed to RNA with T7 polymerase (RiboMax; Promega). Finally, in vitro translated RVFV L-specific RNA was purified after several DNAse treatments and quantified by spectrophotometrical measurement (Qubit; Invitrogen). The number of RNA copies was calculated as previously described (Fronhoffs et al. 2002). Serial 10-fold dilutions of this in vitro translated RVFV L-specific RNA were performed and the standard curves were generated. The limit of detection of this one-step qRT-PCR assay was as low as 3.89 copies/reaction of the in vitro transcribed RNA, equivalent to 1.44 log10 RVFV RNA copies per swab or 1.62 log10 RVFV-RNA copies per mL of blood or per gram of tissue.
Sequencing and phylogenetic analysis
RVFV M segment fragments corresponding to the most variable sequence (Bird et al. 2007) were amplified by conventional RT-PCR upon RNA isolation from sheep blood or infected cells using specific primers. Sequence alignments and neighbor-joining distance trees were generated using the ClustalX 2.0 package software.
Serological tests
Sera from animals inoculated were evaluated for the presence of neutralizing antibodies. Heat-inactivated serum samples were serially diluted (twofold or threefold, starting at dilution of 1:4) in Dulbecco's modified Eagle's medium containing 2% fetal bovine serum, mixed with an equal volume (50 μL) of medium containing 4 × 103 pfu of a viral stock (MP12 strain). This mixture was added to Vero cell monolayers seeded in 96-well plates. After 3 days at 37°C, cells were fixed and stained with 0.75% crystal violet and 10% formaldehyde. The neutralization titer of each sample was determined as the reciprocal of the highest dilution of serum at which 75% neutralization was observed.
Detection of anti-RVFV IgM and IgG antibodies was performed as described previously (Lorenzo et al. 2008). Briefly, ELISA plates were coated with rabbit polyclonal serum anti-RVFV diluted 1/2000 in carbonate/bicarbonate buffer (pH 9.6). After blocking reactive sites with 5% skim milk in phosphate-buffered saline, 5 × 103 pfu of MP12 virus strain was added to a 50 μL volume. After three washing steps with 0.1% Tween-20 in phosphate-buffered saline, diluted heat-inactivated sheep sera was added and incubated. After further washing steps, bound sheep sera was detected with either anti-sheep total IgG or IgM (Zymed-Invitrogen) HRP conjugates. Plates were incubated for 1 h at 37°C, washed three times, and 50 μL of soluble tetra methyl benzidine substrate (Sigma) was added to each well. The plates were then incubated in the dark at room temperature for a further 5 min. The reaction was stopped by the addition of 50 μL of 3 N H2SO4 and the optical density was determined at 450 nm.
Interferon (IFN)-γ assays were performed at 14 dpi using the whole blood bovine IFN test (Bovigam TB; Prionics). Whole sheep blood was first incubated overnight in the presence and absence of 105 TCID50 of virus and the levels of IFN-γ secreted in plasma were then measured by Enzyme-linked Immuno Assay (EIA). The results were expressed for each sample in A450 units after subtraction of the negative control values.
Results
Infection-associated clinical and histopathological observations
Very few clinical signs associated with RVFV infection were observed during the infection. No mortality was recorded. The most evident clinical sign was a sharp increase in body temperature (pyrexia) in nearly all inoculated sheep, reaching 41.5°C–42°C at 2–3 dpi, depending on the individuals (Fig. 1), with the exception of sheep #31 (inoculated with sheep isolate AN 1830), which did not present pyrexia throughout the study.
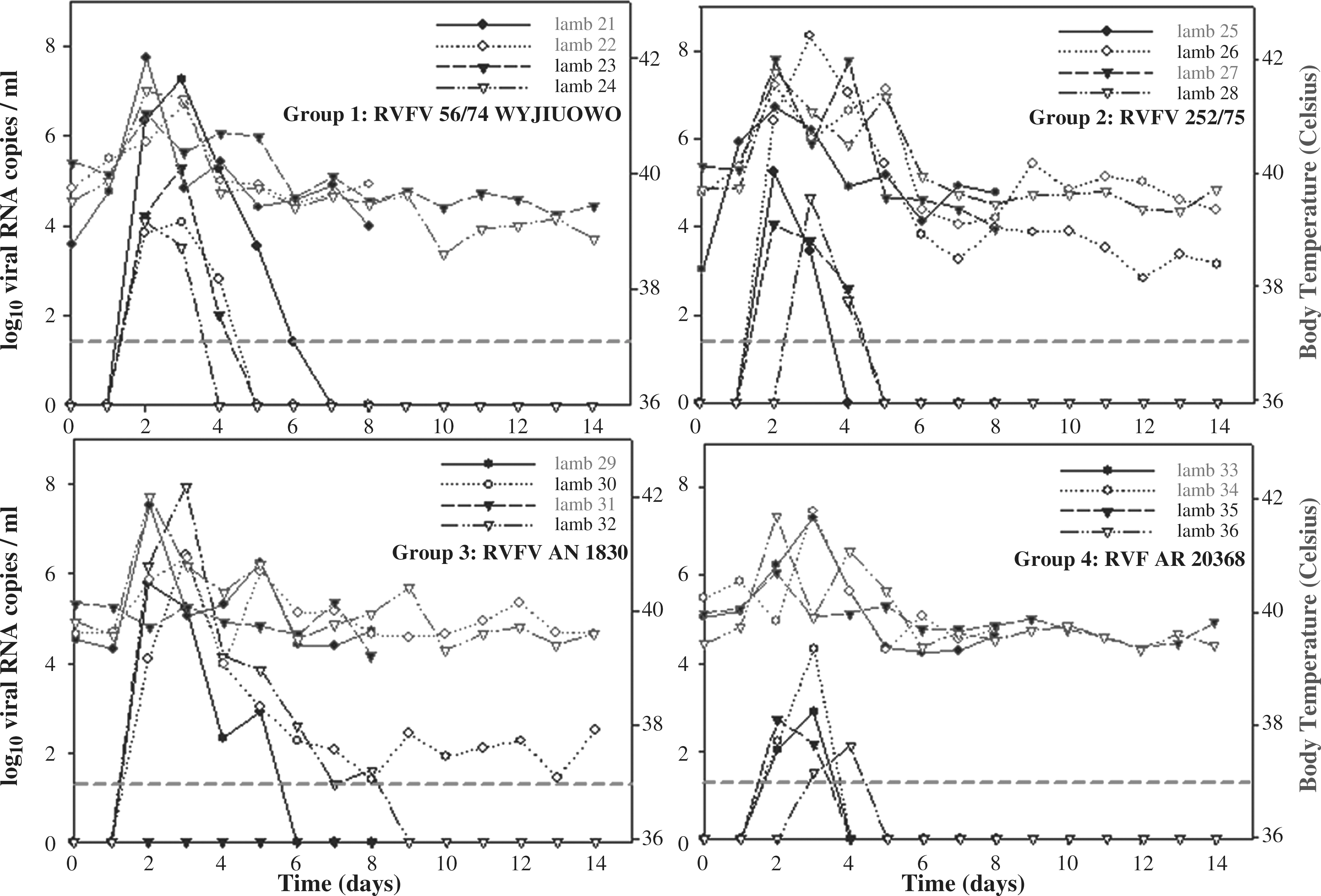
Individual daily body (rectal) temperature and viral load levels in blood samples measured by qRT-PCR of sheep during the whole experiment for each experimental group. Individuals that were necropsied at 8 days post-inoculation are shown in the horizontal dashed line. The limit of detection (LoD) for the qRT-PCR used is shown by the horizontal dashed line qRT-PCR, quantitative retrotranscriptase polymerase chain reaction.
No clinical differences among the inoculated viral isolates were observed. Some lambs became slightly apathic during the fever period, but no loss of appetite was recorded. A well-defined, infiltrative, multifocal, white-spotted, less than 2-mm-diameter hepatic nodules were observed in lambs #23, #26, #28 (14 dpi), and #33 (8 dpi) as major gross finding. No significant pulmonary, lymphatic, or renal gross lesions were recorded. Corneal opacity (“blue eye”) was found at 8–9 dpi in lambs #26 (252/75 group) and #29 and #32 (AN1830 group), being bilateral for sheep #26 and #32. Multifocal lymphoplasmocytic cholangiohepatitis (mild: lamb #34; moderate: lambs #22 and #25; severe: lambs #29 and #33), diffuse lymphoplasmocytic cholangitis (mild: lambs #23, #24, #27, #31, #32, and #35; moderate: lamb #28; severe: lamb #26, associated to fibrosis), and multifocal chronic biliary hyperplasia (related to all cited hepatic lesions) in different intensities were observed histopathologically. Lambs #30, #36, and #38 showed no hepatic lesions. Pulmonary interstitial mononuclear inflammatory infiltrate was observed in some lambs. Such infiltrate was diagnosed as mild in lamb #34. A moderate bronchointerstitial inflammatory pattern was observed in lambs #30 and #33. The rest of findings were considered as incidental following criteria of evidence-based clinical approach (Geyman et al. 2000). Moderate chronic squamous corneal metaplasia, thickness of fibrous stroma, and mild hypertrophy of the posterior corneal endothelium were observed in the three affected corneas. Besides these findings, corneal edema associated with a mild diffuse transmural inflammatory infiltrate was observed in three lambs. Mild perivascular choroidal edema was found at posterior ocular chamber, closely related to retina, in lambs #26 and #32.
RVFV quantitation in blood, swabs, and tissues
Viral loads in blood were quantified by qRT-PCR. Most animals from all groups showed a short increase in viral loads (2–5 days), correlating with pyrexia and peaking at 3 dpi (Fig. 1, black lines). Remarkably, two lambs (#26, 252/75 group, and #30, AN1830 group) showed a long-lasting detection of viral RNA until the end of the experiment (14 dpi).
Despite all RVFV isolates were injected at the same concentration, they differentially replicated in vivo. All sheep inoculated with AN1830 isolates displayed high viral load, ranging 6–8 log10 RNA copies/mL, with the exception of sheep #31, which remained negative for viral RNA detection (and no pyrexic). Also, similarly high viral loads were found in one out of four sheep in the groups inoculated with either the 56/74 cow isolate or the 252/75 isolate (≥106 viral RNA copies/mL). In contrast, the AR20368 mosquito isolate showed the lowest viral load in blood (only 3–4 logs10 RNA copies/mL).
Interestingly, all of the sheep with higher viral loads in blood showed also a longer window for RNA detection. Sheep #21 inoculated with isolate 56/74 was positive for RVFV-RNA detection between 2 and 6 dpi, whereas in sheep #26 RNA was detected for 12 days. Similarly, in the group inoculated with the AN1830 isolate, viral RNA was detected for 4, 12, and 6 days in sheep #29, #30, and #32, respectively. In the remaining sheep the viral RNA was detected for 2–3 days only.
Regarding viral shedding, nasal swabs became positive for RVFV RNA by qRT-PCR, 1 day after the peak in the viral load detected in blood (at 3 dpi) for most animals, lasting from 1 to 6 days (3 days in average). Exceptions to this trend were all sheep infected with the AN1830 isolate, shedding virus as early as 2 dpi, and sheep inoculated with the AR20368 strain, where few nasal swabs scored positive for RVFV-RNA (Fig. 2). Oral swabs became RVFV-RNA positive between 1 and 4 days after the peak of RNA detection in blood samples, lasting for only 1–3 days for most of the animals. Again, only few cases of low RVFV excretion were reported in the AR20368 group. Although nasal swabs showed slightly higher viral loads than oral swabs, both viral loads were consistently lower than those found in blood. There was a clear correlation between viral loads in blood and shedding levels. However, viral excretion occurred even when the viral load in blood was low, as illustrated by the AR20368 group.
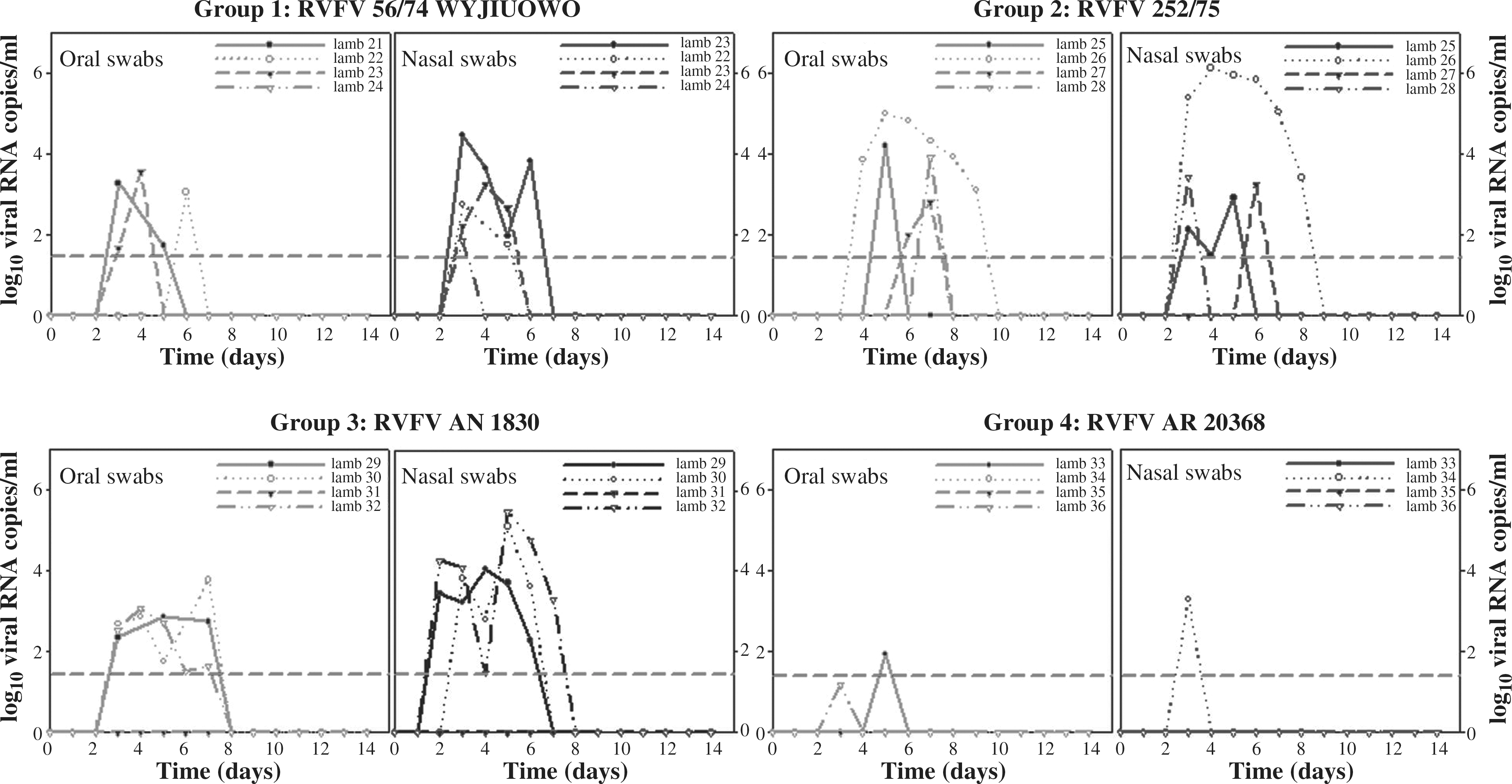
Individual viral shedding as recorded by nasal and oral swabs measured by qRT-PCR, during the whole experiment for each experimental group. Individuals that were necropsied at 8 dpi are shown in red. The LoD for the qRT-PCR used is shown by the horizontal dashed line.
Interestingly, three of the lambs showing high viral load in blood at 3 dpi (#26, 252/75 group; #30 and #32, AN1830 group), ranging between 6.43 and 8.35 log10 viral RNA copies/mL, as well as longer viral load in blood (up to 14 dpi), scored positive for RVFV-RNA in their kidneys, as detected by qRT-PCR, with viral RNA copies ranging between 1.53 and 2.53 log10/g at 14 dpi. No positivity was found by the qRT-PCR assay in tissues from liver, lung, and spleen and lymph nodes taken at 8 or 14 dpi.
Horizontal transmission of RVFV to noninfected, in-contact sentinels
Pyrexia was also recorded in one out of three sentinels, peaking at 4 dpi, delayed 2 days with respect to the inoculated sheep (Fig. 3). This sheep (#38) showed highest viral loads (around 5 log10 RVFV RNA copies/mL) at 4–5 dpi (Fig. 3, blue lines). Sequencing of the variable RVFV M segment region obtained from sheep #38 showed the highest percent of identity with the corresponding sequence of AR20368 mosquito isolate. Therefore, the horizontal infection of sheep #38 was most probably originated by the mosquito isolate.
Viral load in blood (solid lines) and viral excretion (dotted lines) as recorded by nasal and oral swabs and body (rectal) temperature recorded in three lambs from the in-contact control group. The LoD for the qRT-PCR used is shown by a red line.
RVFV-specific immune responses upon in vivo infection
The development of neutralizing antibodies was similar in all infected groups, independently of the strain used and the levels of viral load in blood detected (Fig. 4B). Neutralizing antibody responses were clearly detected at 8 dpi, coincidental with a peak in anti-RVFV IgM titers (Fig. 4A), with maximum titers (up to 1/5000) at the end of experiment (14 dpi), with a concomitant increase in anti-RVFV IgG titers (Fig. 4A). Neutralizing antibody titers (1/2500) were also found in the infected sentinel at 14 dpi, as well as at low levels at 8 dpi, accordingly with a delay in detection of RVFV-RNA in blood and virus excretion.
(
To assess whether other immune mechanisms, such as cell-mediated immune responses, were induced upon RVFV infection, we tested the levels of specific IFN-γ induction in blood at 14 dpi (Fig. 5). Although the overall IFN-γ levels were low, IFN-γ induction was much higher in those sheep infected with cow (56/74) and mosquito (AR20368) isolates, both showing the lowest and shortest viral loads in blood. Contrarily, lower IFN-γ levels were found in those lambs with long, persistent viremia (252/75 and AN1830).
Levels of interferon-γ released in virus-stimulated sheep blood cultures taken at 14 dpi.
Discussion
The risk of the introduction of Rift Valley fever in Europe was assessed recently by a panel of experts (EFSA 2005). In this work, we contribute with some data on the pathogenicity of four African isolates of RVFV in a European breed of adult sheep. The data obtained allow withdrawing some conclusions with respect to the consequences of a RVFV infection of ruminants in Europe. The lack of a severe, fatal disease might be explained by different results, such as the age of the animals used (9 weeks old), as previously reported (Kamal 2009). Alternatively, the passage history of these strains in cell culture might also influence the moderately virulent phenotype observed. In agreement with this last hypothesis, higher viral loads in blood were reached after inoculation with RVFV isolates 252/75 and AN1830, which were less passaged in cell culture than 56/74 and AR20368 (Table 1). The fact that these virus strains were isolated from different animal species might also affect their pathogenic effects. Curiously enough, the two viruses showing lower replication levels were isolated from mosquito and cow.
In all infected animals, viral RNA was detected in blood and quantified by qRT-PCR with inter- and intra-group variations. However, all of them peaked at the same day (3 dpi), starting at 2 dpi, coincidental with the peak of fever. Our results are in good agreement with those from previous studies using different viruses and/or sheep breed (Olaleye et al. 1996). In all animals infected with the AN1830 sheep isolate, viral loads in blood were detected for longer periods (at least 4 days), being able to persist in one lamb in blood and also in kidney throughout the experimental procedure. These same animals shed virus for longer periods and from earlier time points: as soon as at 2 dpi. Interestingly, the only virus isolated from the mosquito reservoir (AR20368) yielded the lowest virus RNA levels in blood and swabs among tested viruses, all isolated from ruminant, reflecting an incomplete adaptation to the mammalian host.
A strong correlation was observed between RVFV-RNA load in blood and kidney, which is considered a secondary target organ (Kamal 2009), with the threshold being around 6.43 log10 RVFV RNA copies/mL at 3 dpi in blood. Therefore, RVF 252/75 and RVF AN 1830 isolates would persist in kidney for longer time than the other isolates once reached certain viremia titers. Interestingly, viral RNA copies were detected in kidney samples from both sheep displaying bilateral blue eye. Viral infections such as RVFV has been related to major ocular changes in humans and animals (Gerdes 2004, Al-Hazmi et al. 2005, Khairallah et al. 2009). Our findings suggest a chronic corneal squeal, which could be associated with drainage failure or inadequate corneal dehydration after a transient viremia (lost of corneal deturgescence). Severe deleterious hepatic pathological changes in kids (necrogranulomas) and hyperplastic overgrowth with vesicular elongated epithelium (even necrotic in some cases) and lymphocytic infiltration were diagnosed in RVF live attenuated vaccinated (Smithburn strain) goats (Kamal 2009). Our results do not show any relation between the presence of hepatic lesion and virus load or excretion in contrast to that reported by Kamal (2009). This fact may point us toward two possibilities with the RFV-infected lambs: (1) a recovery phase of the RVF infection or (2) a chronic hepatic sequel after hepatic replication. Further studies will be carried out to characterize the observed ophthalmic and hepatic lesions. Pulmonary findings were not related to a specific RVF infection. They were considered as incidental findings following criteria of evidence-based clinical approach (Geyman et al. 2000).
Although the route of transmission is not known, the virus isolate responsible for the infection observed in one of the three sentinel animals was, surprisingly, the AR20368 mosquito isolate, as it was stated by genome sequencing. As sheep inoculated with this isolate did not display the highest nasal and oral shedding titers, other factors might be involved in the observed transmission. Also, as all inoculated animals were housed in the same box, potential cross-contamination events cannot be ruled out. In any case, the high and long viral load in blood observed might be advantageous for virus dissemination by competent vectors, and therefore, competence studies with the four virus isolates could be addressed in the future. Further, these long-term viremic infected lambs may be potentially dangerous (Easterday et al. 1962, Erasmus et al. 1981, Woods et al. 2002), for example, in the case of being slaughtered, for its risk of viral transmission to human by direct contact with infected fluids. Our results confirm that horizontal RVFV transmission is not an unusual event, as it has been previously described with in-contact American sheep becoming infected with the RVFV Zagazig 501 isolate (Harrington et al. 1980). Therefore, if infection pressure inside herd is high enough, it could be stated that some strains of RVFV could be horizontally transmitted, independently of vector presence.
Induction of a neutralizing antibody response is a good correlate for controlling infection and protection after challenge with RVFV (Harrington et al. 1980, Hubbard et al. 1991, Morrill et al. 1991). For most animals, virus was cleared from the body fluids before neutralizing antibodies were detected, and thus it is reasonable to speculate that other early immune mechanisms play a role in the elimination of the virus, most probably related with nonspecific innate immune responses. Although a clear “in vitro” neutralizing activity was observed in all sera, this immune response was undistinguishable from the virus isolate used. In contrast, a clear correlation was observed between specific IFN-γ detection at 14 dpi and viral clearance, with persistently infected sheep showing almost no specific IFN-γ responses. Curiously enough, persistently infected cells also showed the lowest IFN-γ response to the unspecific phytohemagglutinin (PHA) stimulation, suggesting a transient immune suppression. Understanding the immune mechanisms involved in viral clearance and protection might be essential to design efficient vaccines in the future.
In conclusion, adult lambs from the European Ripollesa breed are susceptible to experimental infection with African RVFV isolates of different origins. Transient pyrexia, rising viral loads in blood, and viral shedding were recorded for all viruses, with no severe clinical signs of disease. Further, both specific humoral and cellular responses were detected after viral infection, thus opening the possibility of using this animal model to test potential vaccines against RVFV.
Footnotes
Acknowledgments
The authors thank T. Gerdes and S. Smith (ARC-OVI, South Africa) for providing the four RVFV isolates used in this study, and C.J. Peters and R. Tesh (WRCEVA, Galveston, TX) for providing the MP12 strain used for the immunological assays. The authors also thank David Wallace for critical reading of the manuscript and Maite Martín (CReSA) for her excellent technical assistance.
Disclosure Statement
No competing financial interests exist.