
Abstract
West Nile virus (WNV) is a mosquito-borne flavivirus naturally circulating in wild bird populations. The virus is also capable to infect a broad range of vertebrate species. Humans and equines are highly susceptible and can develop mild flu-like illnesses as well as severe encephalitis leading to fatalities. Most recently, WNV was found to circulate in countries close to Germany, such as France, Czech Republic, Italy, Austria, and Hungary. Given this epidemiological situation its spread to Germany cannot be ruled out. As no data on the WNV situation were available for Germany for the most recent past, we have conducted a serological survey to reveal WNV antibodies in wild birds. More than 2700 blood samples from migratory and resident birds representing 72 species that were collected during 2005–2009 were tested using an immunofluorescence assay and partly by micro-virus neutralization test. By immunofluorescence assay WNV-reactive antibodies could be demonstrated in 11 wild bird species. Similarly, WNV-neutralizing antibodies were revealed in migratory birds belonging to 10 species, but not in resident birds. According to the absence of WNV-reactive antibodies in resident birds and the absence of WNV-specific RNA in all investigated bird samples, there is currently no evidence for a WNV circulation in Germany.
Introduction
However, in the light of global trade and travel and also possible climate changes and because of the first appearance of WNV in Austria, Hungary, and Italy, we decided to carry out a large-scale study on the WNV antibody status of migratory and resident birds belonging to a broad variety of species in Germany. The demonstration of antibodies in resident birds would ultimately prove that WNV is already circulating locally.
Materials and Methods
Sera/plasma
Birds were bled by puncturing their wing veins, and after blood separation sera were stored at −20°C and cruors at −70°C. Reference sera from vaccinated and naturally WNV-infected chicken with a known titer (kindly provided by the U.S. Department of Agriculture) were used to calibrate the test system. Prevaccination and postvaccination (West Nile-Innovator®; Fort Dodge Animal Health) sera from chicken and ducks served as negative and positive controls.
Indirect immunofluorescence assay
A modified protocol of a commercial WNV immunofluorescence assay (IFA) slide test kit (Euroimmun) was used generally following the manufacturer`s instructions. Bird sera were used at 1/30 dilutions and goat anti-bird fluoresceine-isothiocyanate-labeled antibodies (Bethyl Laboratories) diluted 1/100 for the detection of binding antibodies. As these labeled antibodies were only low or nonreactive to immunoglobulins (Ig) of some wild bird species as determined in a dot blot format (Table 1), IgY of these species was observed by using another conjugate, that is, rabbit anti-bird antibodies (ICL) diluted 1/200 followed by goat anti-rabbit fluoresceine-isothiocyanate-labeled antibodies (Invitrogen) at 1/200 dilution. Endpoint titrations were carried out for reactive and inconclusive samples.
IgY, immunoglobulin Y.
Micro-virus neutralization test
Micro-virus neutralization tests (micro-VNTs) were carried out using a 96-well plate format using a published method with minor modifications (Linke et al. 2007). Dilutions of heat-inactivated sera in the tissue culture medium were incubated in duplicate for 1 h at 37°C with equal volumes of 100 tissue culture infectious doses (TCID50) of a WNV lineage 1 strain (NY99) and then added to 1-day-old Vero E6 cell monolayers. After 1 h incubation at 37°C, a fresh cell culture medium was added and cytopathic effects were recorded by crystal violet staining after 6–7 days. Titers were calculated following the Behrens–Kaerber method. Neutralization effects at 1/10 or higher dilutions were qualified positive. Positive samples were retested using dilutions ranging from 1/10 to 1/2560 to determine end point titers. Reference sera from vaccinated and naturally infected chicken were used for calibration.
One-step quantitative real-time–polymerase chain reaction assay
Viral RNA was extracted from whole blood or cruor by using the QIAmp Viral RNA kit or RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions. Viral RNA was detected by a one-step quantitative real-time–polymerase chain reaction (qRT-PCR) assay (Eiden et al. 2010). Primers and probe were targeted to the region coding for the nonstructural protein NS2A, which enables the detection of both WNV lineages 1 and 2. The qRT-PCR assay was performed employing the Mx3000P QPCR system (Agilent/Stratagene) using the Quantitect Probe RT-PCR Kit (Qiagen) in a total volume of 25 μL.
Results
In this study 2736 avian blood samples originating from 72 species of migratory and resident birds were tested for the presence of WNV antibodies (Table 2). Samples were collected in the period 2005–2009 at multiple rural and urban locations in several German states (Fig. 1). The birds were trapped with mist nets and funnel traps (small passerines) or cannon nets (greylag goose), individually marked with numbered metal or plastic rings, bled, and released. Small migratory passerine birds were captured and bled at the time of return from their wintering sites. Nestlings were ringed and bled at nest sites (white stork, osprey, and hooded crow). Several injured wild birds were bled in animal clinics during veterinary treatment.
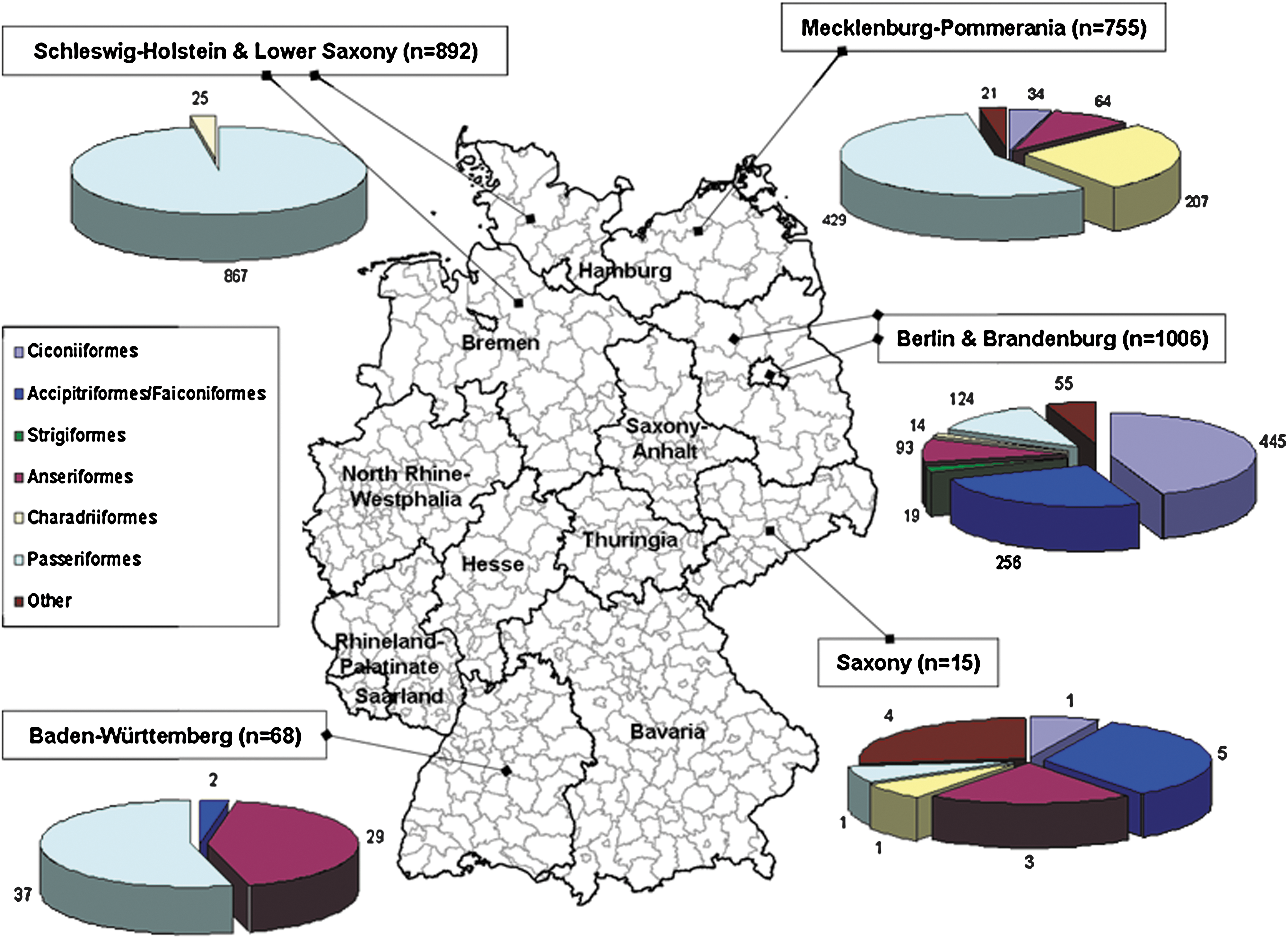
Location of sampling sites of wild bird species from Germany.
Zoo/park bird.
Including 15 cytotoxic samples not analyzable.
Neozoon bird species.
IFA, immunofluorescence assay; VNT, virus neutralization test; R, resident species; PM, partial migrant; SD, short-distance migrant; LD, long-distance migrant.
Using an indirect IFA, 23 of the 2736 tested samples representing 11 wild bird species showed antibodies to WNV with titers ranging from 1/15 up to 1/1920 (Table 3). Five of these avian species were long-distance (trans-Saharan) migrants, four species were short-distance or partial migrants, and two species were residents. The majority of 463 tested white storks were nestlings, whereas 2 of the 3 IFA-reactive samples originated from adult birds. Another three samples of white stork nestlings remained inconclusive. All 22 ospreys tested by IFA—of which 2 were reactive—originated from nestlings or juveniles. Two samples of an adult barn swallow that was accidentally sampled twice in June and August 2008 were IFA reactive with a considerable drop in titer from 1/1920 down to 1/480 a few weeks later. Three IFA-reactive sera from resident birds originated from a juvenile common magpie (Pica pica) and two adult tawny owls (Strix aluco). Sera from two wood pigeons (Columba palumbus), one garden warbler (Sylvia borin), one tawny owl (S. aluco), one long-eared owl (Asio otus), and one lesser spotted eagle (Aquila pomarina) gave inconclusive IFA results.
n = tested individuals of species.
Including two samples of one barn swallow accidentally captured and bled twice in summer 2008.
Results for 1086 samples that were also analyzed by micro-VNT are shown in Table 4. Two of the 28 white stork sera that exhibited WNV-neutralizing antibodies originated from adult individuals, whereas all the others were taken from nestlings. Because of the very small sample volumes, it was not possible to investigate most of the Passeriformes by micro-VNT. Therefore, the findings in some IFA-reactive small Passerine plasmas (common redstart, blackcaps, etc.) could not be reconfirmed by micro-VNT. However, none of the resident bird species sera with IFA reactive (common magpie and two tawny owls) or inconclusive results contained WNV-neutralizing antibodies, whereas a few IFA negative sera showed virus-neutralizing activity (gray heron, red kite, herring gull, and Eurasian woodcock). In summary, of 23 samples found reactive by IFA, 6 samples could not be tested by micro-VNT because of insufficient sample volumes. From the 17 IFA-reactive samples that were also tested by micro-VNT, 9 showed also neutralizing antibodies, but 8 IFA-reactive samples could not be confirmed by micro-VNT. On the other hand, 30 IFA-negative samples from migratory birds showed WNV-neutralizing antibodies. Compared to the micro-VNT as gold standard, the IFA used here was found to be highly specific (99%), although not as sensitive (25%) due to the false-negative results.
Titers >1:10 are considered to be positive; n = tested individuals of species.
Including two samples of one Barn Swallow accidentally captured and bled twice in summer 2008.
Detection of viral RNA was also attempted by a one-step qRT-PCR on 237 blood samples (94 birds of prey, 69 Ciconiiformes [mainly white storks], 43 Anseriformes, 19 Columbiformes, 4 Gruiformes [Eurasian coots], 3 Charadriiformes [2 herring gulls and 1 black-headed gull], 3 Passeriformes [2 common redstarts and 1 rook], 1 great spotted woodpecker, and 1 little grebe). In none of these blood samples WNV-specific RNA was revealed.
Discussion
WNV outbreaks or virus activity have been observed in Central European countries most recently (Bakonyi et al. 2006, Figuerola et al. 2007, Höfle et al. 2008, Hubálek et al. 2008a, 2008b, Jiménez-Clavero et al. 2008, López et al. 2008, Weissenböck et al. 2010). Migratory birds can act as carriers for this zoonotic pathogen and carry it over long distances (Hubálek 2000, Malkinson et al. 2001, 2002, Malkinson and Banet 2002). The detection of WNV antibodies in resident bird populations is indicative for a local virus circulation. Germany is located not far away from the most recent outbreaks in Austria and Hungary; therefore, it is necessary to closely monitor the WNV status of birds as indicators for a potential virus introduction. Thus, we carried out a serological study on migratory and resident birds captured in Germany. In most cases, where only spurious amounts of sera were available, these were initially screened by indirect IFA. Reactive sera as well as sera from species from which larger amounts of blood samples were available were also analyzed by micro-VNT. Two different IFA conjugates were used that detect IgY of a broad variety of avian species. Most reactive samples with high antibody titers gave corresponding results in both methods, and 96% of the micro-VNT-positive sera were detected by IFA too. As the micro-VNT displayed a higher analytical sensitivity, IFA missed some low reactive samples. However, the small serum volumes, for example, from passerines, make the IFA indispensable and the method of choice. No indirect WNV antibody enzyme-linked immunosorbent assay for detection of avian IgY antibodies from a wider range of species is available to date. A commercially available competitive enzyme-linked immunosorbent assay (ID Screen®; West Nile Competition, ID Vet) is suitable for the multi-species detection of antibodies, but it requires sample volumes that are larger than those that can be obtained from small birds (e.g., passerines).
Antibodies to WNV were found in 23 of 2736 (0.8%) tested samples by IFA and in 41 of 1086 (3.7%) sera by micro-VNT. These results compared generally to those reported from earlier studies (Linke et al. 2007). IFA reactions can be caused by cross-reacting antibodies due to closely related Flaviviruses. Cross-reactions with closely related viruses from the Japanese encephalitis virus group can also occur in the highly specific virus-neutralization test. Usutu virus (USUV) is such a close relative of WNV that is also circulating in Central Europe; it emerged in Austria in 2001 (Weissenböck et al. 2002, Bakonyi et al. 2004), and spread recently to Hungary (Bakonyi et al. 2007), Switzerland, and Italy (Manarolla et al. 2010). The endemic in Austria has been well characterized. Initially, USUV caused major fatalities primarily among resident birds like blackbirds, owls, and birds of prey (Weissenböck et al. 2003, Chvala et al. 2007), peaking in 2003, whereas from 2004 onward, USUV-associated bird mortality decreased in Austria significantly. Serological studies indicated in 2003 and 2004 a rather small proportion of seropositive wild birds (<10%), whereas the percentage of seroreactors raised to >50% in 2005 and 2006, suggesting the establishment of herd immunity (Meister et al. 2008). For Germany, only few data on the USUV situation are available. In samples collected in the period 2002–2005, Linke et al. (2007) found that 3 out of 25 WNV-IFA-reactive samples (originating from migratory birds, i.e., white stork, common redstart, and osprey) contained USUV-neutralizing but not WNV-neutralizing antibodies. Hence, although unlikely, IFA-WNV-reactive samples in the present study could also indicate USUV infections in these birds. Forty-five sera that were reactive in the WNV IFA or micro-VNT were therefore also assayed in a USUV-micro-NT (using USUV strain Vienna_2001) with negative results (titers <1:10) in all cases (data not shown).
Further, there is no evidence for wild bird infections with tick-borne encephalitis virus, another closely related flavivirus that is endemic in some areas of Germany, although birds can be infested by tick-borne encephalitis virus–infected ticks (Ernek et al. 1968, Waldenström et al. 2007).
In the study presented here three sera from resident birds (one common magpie and two tawny owls) were reactive in the IFA screen. These IFA reactions were not confirmed by micro-VNT, which indicates nonspecific or cross-reactive antibodies. Vice versa, WNV-neutralizing antibodies were found exclusively in migratory birds (i.e., long-distance, short-distance, and intermediate migrating birds) and were most likely the consequence of WNV infections at the resting sites in Africa or Southern Europe or during their journey to Central Europe. Low or borderline micro-VNT titers were also found in nestlings or juveniles of such migratory birds (e.g., white storks). These antibody levels are much lower than the titers that are found in experimentally (Komar et al. 2003) or naturally WNV-infected birds (Figuerola et al. 2007). We therefore assume that these antibodies represent maternal antibodies that have been passed on via egg yolk from WNV-competent adults. This assumption is also supported by the absence of WNV-neutralizing antibodies in all resident bird species investigated. Assuming cross-reacting antibodies to other non-WNV Flaviviruses would be another, although less likely, explanation.
For a successful long-distance spread and the establishment of a self-sustained transmission cycle in a new environment, several factors are critical: suitable population densities of relevant host and vector species, congruent population ecologies, and also environmental and climatic conditions that allow the productive amplification in these hosts. Essentially, infected birds from WNV enzootic areas must endure the long and exhaustive flight to Central Europe and then be blood-sucked by susceptible mosquitoes. Alternatively, a sustained infection cycle within groups of migratory birds is also conceivable, but less likely. WNV infections are characterized by only a short viraemia in most, if not all, affected birds (Komar et al. 2003) and a minimum blood virus titer is required for a successful vector infection (Goddard et al. 2002, Turell et al. 2002). The WNV amplification in mosquitoes strongly depends on ambient temperatures and in new vertebrate species on their innate and adaptive immune responses (Reisen and Brault 2007).
On the other hand, the multiple WNV cases in the United States and Canada as well as the emergence and recent spread of a lineage 2 WNV in Hungary and Austria (Bakonyi et al. 2006, ProMED-mail 2009, Weissenböck et al. 2010) strongly emphasize that this virus–bird–mosquito infection cycle can also operate in more temperate regions. Therefore, a WNV introduction into Germany by migratory birds or other routes followed by a self-sustained perpetuation is also conceivable. Resting sites for wild birds where a high population density exists and where susceptible vectors live are high-risk areas for such an introduction. The use of sentinel birds that are detained at such sites could be a suitable WNV-monitoring approach (Buckley et al. 2006, Chevalier et al. 2008, Globig et al. 2009, Ziegler et al. 2010). Likewise, monitoring programmes for horses could be useful for predicting the risk of human WNV infections (Ward and Scheurmann 2008).
Footnotes
Acknowledgments
We are indebted to F. Bairlein and P. Becker (Institute of Avian Research “Vogelwarte Helgoland,” Wilhelmshaven, Germany), W. Fiedler (Max Planck Research Centre of Ornithology, Vogelwarte Radolfzell, Radolfzell, Germany), U. Köppen (Landesamt für Umwelt, Naturschutz und Geologie Mecklenburg-Vorpommern, Beringungszentrale, Greifswald, Germany), L. Brunnberg (Fachbereich Veterinärmedizin, Kleintierklinik, Freie Universität Berlin, Berlin), E.F. Kaleta (Fachbereich Veterinärmedizin, Klinik für Vögel, Reptilien, Amphibien und Fische, Justus-Liebig-Universität Giessen, Giessen), J.B.W. Wolf (Max Planck Institute for Evolutionary Biology, Plön, Germany), N. Kenntner (Leibniz Institute for Zoo and Wildlife Research, Berlin, Germany), and the Veterinary Authorities of the Federal States of Germany for providing samples and technical support. Many thanks go to all bird ringers for their essential assistance during the fieldwork and to I. Nedow for excellent technical assistance. This project was supported by the German Federal Ministry of Education and Research (network project “Emerging Arthropod-Borne Viral Infections in Germany: Pathogenesis, Diagnostics and Surveillance”).
Disclosure Statement
No competing financial interests exist.