
Abstract
Flaviviruses of the Japanese encephalitis virus (JEV) antigenic complex, including West Nile virus (WNV), are recognized as emerging and reemerging pathogens. Circulation of flaviviruses has been recently detected in different mosquito and vertebrate species in several European countries. A serosurvey study was carried out to evaluate the circulation of WNV and other flaviviruses of the JEV antigenic complex in different wild bird species in Spain between 2006 and 2009. Seropositiviy against JEV using a competitive enzyme-linked immunosorbent assay was found in common coot, Montagu's Harrier, black kite, black vulture, Bonelli's eagle, Spanish imperial eagle, Egyptian vulture, and Eurasian spoonbill. Seropositivity to JEV antigenic complex viruses was significantly higher in samples collected during autumn compared with animals sampled during summer. Significantly higher seroprevalence was also observed in 2007 compared with 2009, whereas there were no significant differences in seropositivity among taxonomic levels, migratory versus resident behavior, body size (large vs. medium), or habitats (free-ranging vs. captivity). Neutralizing antibodies against WNV were detected in common coot and Spanish imperial eagle using a virus-neutralization test. Oral shedding of WNV was not detected in any of the Spanish imperial eagles, Egyptian vultures, Eurasian Spoonbills, Lammergeiers, and the Black vultures analyzed by means of real-time reverse transcription–polymerase chain reaction. The results indicate that WNV and others flaviviruses of the JEV antigenic group circulated in migratory and resident wild bird species in Spain between 2007 and 2008. Further studies are necessary to determine the precise role that each of these wild bird species, some of them cataloged as “near threatened,” “vulnerable,” or “endangered,” play in the epidemiology of those viruses.
Introduction
WNV was originally identified in Africa, in the West Nile region of Uganda, in 1937 (Smithburn et al. 1940). It is responsible for infection in a wide range of vertebrate species, including birds, reptiles, amphibians, and mammals. The sylvatic cycle of WNV is usually sustained between ornithophilic mosquito species and wild birds, which are reservoir hosts (Smithburn et al. 1940, Burke and Monath 2001, Hayes et al. 2005). Although virus transmission between hosts usually occurs by means of mosquito or tick species (Burke and Monath 2001, Lawrie et al. 2004), direct transmission through cloacal and oral shedding by WNV-infected birds has been also reported (Langevin et al. 2001, Banet-Noach et al. 2003, Komar et al. 2003). Humans and horses are considered incidental or dead-end hosts, and are not thought to be able to transmit the virus. A variety of symptoms including fever and encephalitis, and mortality have been observed in wild birds, horses, and humans infected by WNV.
WNV is widely distributed throughout Africa, America, Australia, Asia, and Europe, where it has caused outbreaks and sporadic cases of the disease (Hubálek and Halouzka 1999, Komar et al. 2001). The first outbreak of WNV in North America was reported in 1999. Since then, the virus was rapidly dispersed causing illness and mortality in humans and occasioning severe disease and mortality among avian hosts (Lanciotti et al. 1999, Steele et al. 2000, LaDeau et al. 2007). WNV was initially considered to have a minor health effect in Eurasian birds from the Mediterranean basin (Zeller and Schuffenecker 2004). However, this is in contradiction with the fact that several outbreaks have been reported in humans in this area, which has also highlighted the public health importance of WNV infections (Barzon et al. 2009, Monaco et al. 2009, Papa et al. 2010).
Evidence of WNV circulation in Spain has been demonstrated by detection of WNV-neutralizing antibodies in humans (Lozano and Filipe 1998, Bofill et al. 2006, Bernabeu-Wittel et al. 2007), horses (Jiménez-Clavero et al. 2007, 2010), and both migratory and resident wild bird species (Figuerola et al. 2007a, 2008, López et al. 2008, 2010). Infections with clinical symptoms in humans (Kaptoul et al. 2007), raptors (Höfle et al. 2008, Jiménez-Clavero et al. 2008), and horses (RASVE 2010) have been also described in Spain. In addition, the circulation of other flaviviruses has been reported in different mosquito species in Spain (Busquets et al. 2008, Aranda et al. 2009, Sánchez-Seco et al. 2009).
Although several studies have assessed the influence of factors such as migratory behavior, body size, or taxonomic group in the epidemiology of WNV in Spain (Figuerola et al. 2007a, 2007b, 2008, López et al. 2008), the information on the role played by different wild bird species remains scarce. The objective of the present study was to evaluate the circulation of WNV and other flaviviruses of the JEV antigenic complex in different wild bird species in southern Spain (Andalusia).
Materials and Methods
Collection of samples
A total of 201 serum samples from 15 different wild bird species were collected in several areas of Andalusia (southern Iberian Peninsula; Fig. 1) between 2006 and 2009 (Table 1). The samples were obtained from clinically healthy free-ranging (n = 150) and captive (n breeding centers: 35; n rescue centers: 16) adult animals during routine health checks and radiotagging. All animals were tested only once.
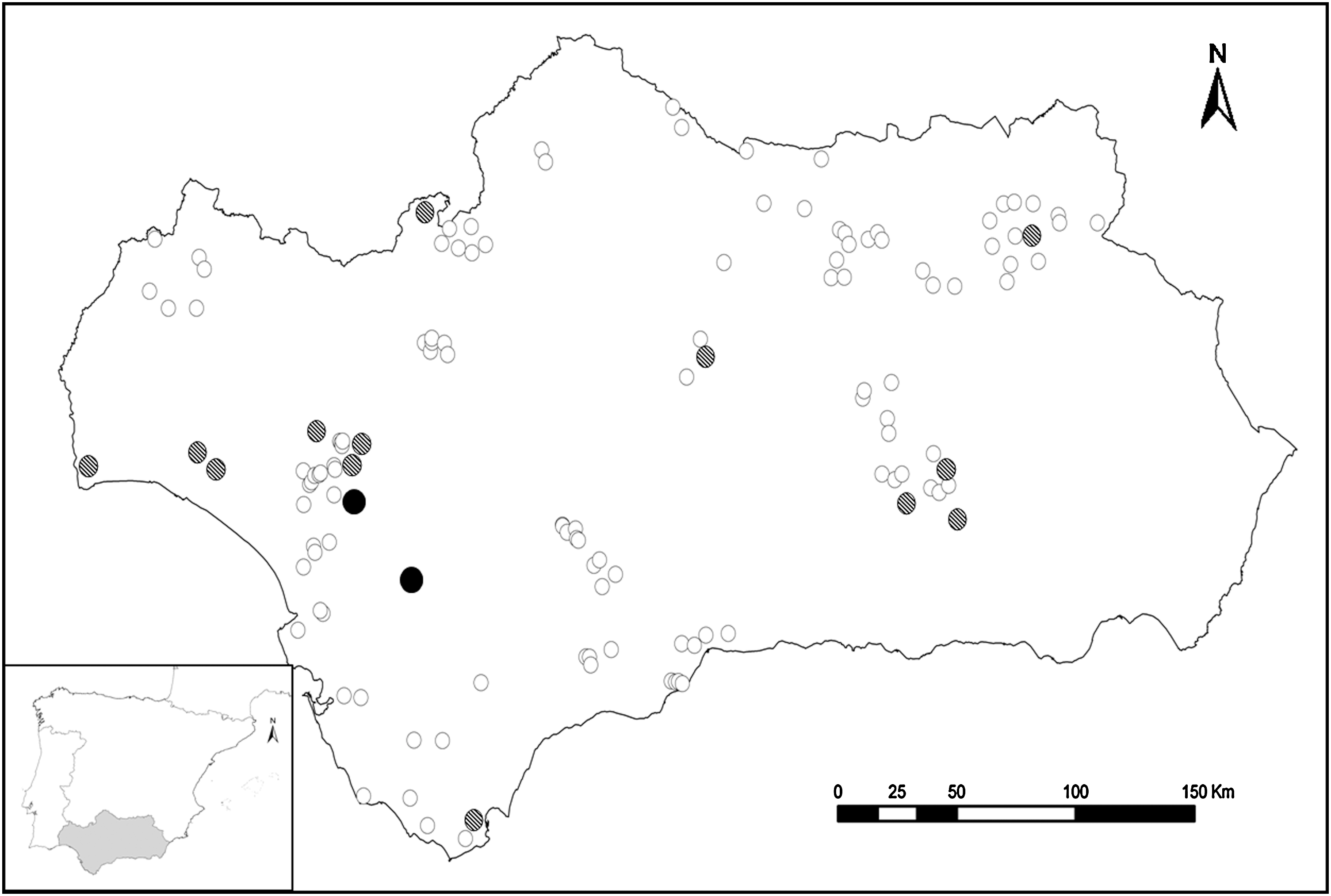
Sampling locations. White and crossed dots indicate absence and presence of antibodies against flavivirus in the sampled birds, respectively. Black dots indicate presence of antibodies against West Nile virus (WNV) in the sampled birds.
Based on IUCN (2008).
Titer 1:20.
Titer 1:80.
All samples were negative.
cELISA, competitive enzyme-linked immunosorbent assay; VNT, virus-neutralization test; RRT-PCR, real-time reverse transcription–polymerase chain reaction.
Although the weight of the sampled animals could not be measured, the birds were classified as large (n = 132) or medium (n = 69) size based on an interspecific size ranking of the species included in the study. Of the 201 birds, 104 were classified as residents and 97 as migrants. Values for these variables were obtained according to the literature (Cramp and Perrins 1994) and the opinion of ornithologists.
Further, oral swabs from 51 birds, including 28 free-ranging birds and 23 captive animals from breeding centers, were collected between May and July 2009.
All samples were provided by the Department of Environment of the Regional Government of Andalusia. Several of the species analyzed are included in the Conservation Programs by the Regional Government of Andalusia and are listed as “near threatened, “vulnerable,” or “endangered” by the International Union for the Conservation of Nature, BirdLife International (IUCN 2008) (Table 1).
Blood samples were taken with syringes from the brachial or femoral vein. Samples were collected into sterile tubes without anticoagulant and centrifuged at 400 g for 15 min. Then, serum was separated and stored at −20°C until analyzed.
Serological assays
A commercial competitive enzyme-linked immunosorbent assay (cELISA; ID Screen® West Nile Competition, ID Vet, Montpellier, France) was used. The microwells in this cELISA were coated with a purified extract of the WNV. It has been designed to detect horse and different avian species antibodies directed against the envelope protein (pr-E) of this virus, which contains an epitope common to JEV antigen complex.
Sera positive for antibodies to flaviviruses of JEV antigen complex were then tested for WNV-neutralizing antibodies (strain Eg101) by a micro virus-neutralization test (VNT) as described previously (Weingartl et al. 2003, Figuerola et al. 2008). Serum samples were inactivated at 56°C for 30 min prior to analysis. Dilutions of tested sera (25 μl) were mixed with a 25 μl volume containing 100 median tissue culture infectious dose (TCID50) of WNV strain Eg101 and incubated for 1 h at 37°C in Eagle's minimum essential medium supplemented with
Real-time reverse transcription–polymerase chain reaction
Oral swabs were analyzed by real-time reverse transcription–polymerase chain reaction (RRT-PCR) for detection of WNV lineages 1 and 2 as previously described (Jiménez-Clavero et al. 2006), using a Fast7500 equipment (Applied Biosystems, Foster City, CA). Briefly, viral RNA was extracted from oral swabs using QIAamp Viral RNA extraction kit (Qiagen, Valencia, CA), obtaining 40 μl of eluted viral RNA. The 3′ NC fragment amplification was carried out using primers, TaqMan MGB probes, and amplification conditions indicated by Jiménez-Clavero et al. (2006). The TaqMan MGB-RRT-PCR was performed using the commercial one-step RT-PCR master mix reagents (Applied Biosystems, Foster City, CA) following the manufacturer's instruction and using 5 μl of eluted RNA in a total volume of 25 μl.
Statistical analysis
The proportion of birds seropositive to flaviviruses of the Japanese Encephalitis antigenic group and its 95% confidence interval (CI) were calculated. The associations between seropositivity to flaviviruses and the variables (taxonomic level, season, year of sampling, migratory behavior, body size, habitat, and the level of interaction among them) were evaluated by means of a generalized linear model. A binomial error distribution with a logit link function was used for the analysis. The model was implemented in the statistical software package SAS (version 9.1; SAS Institute, Inc., Cary, NC). Differences were considered statistically significant when p < 0.05.
Results
Fifteen of 201 sera tested (7.5%; 95% CI: 3.9–11.1) were positive to flavivirus of the Japanese Encephalitis antigenic complex antibodies using the cELISA test. In addition, 8 of the 15 species analyzed (53.3%) had at least one seropositive individual (Table 1). Frequency of seropositivity to flaviviruses among taxonomic levels, seasons, years of sampling, migratory versus resident behavior, body size, and habitats is shown in Table 2. Significantly higher seropositivity against flaviviruses was observed in birds sampled during autumn compared with animals sampled during summer (p = 0.03) and in samples collected in 2007 compared with 2009 (p < 0.0001). No statistically significant differences among taxonomic levels, migratory versus resident behavior, body size, or habitats were observed.
Only free-ranging birds were included.
Neutralization to WNV at 20-fold or higher dilutions was detected in 2 of the 12 birds seropositive to flaviviruses, with titers ranging from 1:20 to over 1:80 (Table 1). Overall seropositivity to WNV was 1.0% (95% CI: 0.0–2.2). Neutralizing antibodies with a titer of 1:10 were also detected in a Eurasian spoonbill. Unfortunately, three of the samples seropositive to flaviviruses (two of the Spanish imperial eagles and one of the Bonelli's eagle) could not be tested using VNT because of limited sample volume.
Oral shedding of WNV was not detected in any of the Spanish imperial eagles (n = 20), Egyptian vultures (n = 10), Eurasian spoonbills (n = 11), Lammergeiers (n = 9), and Black vulture (n = 1) analyzed by RRT-PCR.
Discussion
The results of the present study confirm the circulation of WNV and other members of JEV antigenic complex in captive and free-ranging wild bird species from southern Spain. Fifteen of the 201 wild birds analyzed by means of the cELISA tested positive. Twelve of these 15 birds were retested by VNT, and 2 of them presented neutralizing antibodies against WNV. The cELISA assay has been shown as appropriate for WNV serologic surveys of free-ranging and captive sentinel birds (Ebel et al. 2002, Blitvich et al. 2003), although the VNT is regarded as the gold standard in flaviviruses serology and is generally more specific than other serological techniques (Weingartl et al. 2003). The presence of a Eurasian spoonbill with a VNT titer of 1:10 could be due to an old contact with WNV or contact with a closely related flavivirus (Hubálek et al. 2008b).
On the other hand, the presence of animals that were positive to the cELISA and negative to the VNT may be an indication of cross-reaction with other antigenically related flaviviruses (Gibbs et al. 2005, Bofill et al. 2006). In this sense, USUV has been recently detected in Spain (Busquets et al. 2008) and other European countries (Weissenböck et al. 2002, Buckley et al. 2003, Linke et al. 2007, Bakonyi et al. 2007, Steinmetz et al. 2007, Lelli et al. 2008, Manarolla et al. 2009). Unfortunately, the presence of neutralizing antibodies against USUV could not be assessed by VNT because of limited sample volume. Flaviviruses detection has an ecological relevance, because, even though they are considered nonpathogenic in their natural habitat, they may become pathogenic in other geographic areas with a population immunologically naive. On the other hand, the lack of correlation between values from the ELISA and the VNT titers could suggest that both tests do not necessarily detect the same antibody types (Blitvich et al. 2003).
To our knowledge, this is the first report of flaviviruses circulation in Egyptian vulture, Eurasian spoonbill, Bonelli's eagle, and Montagu's harrier. The presence of antibodies indicates that these birds were exposed and responded serologically to the contact with members of JEV antigenic complex. All animals tested in the present study were adults, so the antibodies detected were not the result of the passive transfer of maternal immunity (Gibbs et al. 2005).
The higher seroprevalence observed in autumn is in agreement with what has been previously reported (Figuerola et al. 2008). Vector or host density and environmental factors are possible causes implicated in the higher seroprevalence observed during 2007 compared with 2009. However, given the low number of animals sampled in autumn and during 2009, the differences found in seroprevalences in the present study must be interpreted with care. No statistical differences between seroprevalence in captive and free-ranging birds were observed, which is in agreement with what has been previously reported (López et al. 2010).
Migratory behavior was not directly related to antibody prevalence. Although some authors reported a higher seroprevalence in migratory birds (Lopez et al. 2008), our results are in accordance with other studies (Figuerola et al. 2008, López et al. 2010). The presence of seropositive resident birds is an evidence of the local circulation of WNV between 2007 and 2008 in the study area.
Our results indicate limited WNV activity among the species analyzed in Spain between 2006 and 2009, as only 1.0% of the wild birds analyzed were seropositive against WNV. This is in accordance with findings from other studies carried out in the last few years, in which no or limited WNV circulation was reported in Spain [3.8% (López et al. 2008) and 2.2% (López et al. 2010)] and other Mediterranean and European countries: Morocco [3.5% (Figuerola et al. 2009)], Germany [1.6% (Linke et al. 2007)], France [0.0–4.8% (Lena et al. 2006, Jourdain et al. 2007, 2008, Balança et al. 2009)], Poland [5.2% (Hubálek et al. 2008a)], or the Czech Republic [5.9% (Hubálek et al. 2008b)]. However, higher seroprevalences have been also reported in Spain [7.8–10.4 (Figuerola et al. 2007a, 2008)], France [8.0% (Hars et al. 2004)], the United Kingdom [14.7% (Buckley et al. 2003)], and Italy [49.1% (Monaco et al. 2009)]. Variations in the species analyzed, serological methods, migratory behavior, climate conditions, epidemic time, size, or origin of the samples may explain these differences.
Although antibodies against flavivirus were detected in different areas from Andalusia, seropositivity to WNV was only confirmed in the western region (Fig. 1). This area is a usual route of different migratory species between Europe and Africa. In fact, WNV circulation has been recently reported in western Andalusia (Figuerola et al. 2008, López et al. 2008, 2010).
The only specimen of common coot analyzed presented antibodies against WNV. A high seroprevalence and seroconversion were reported in this species in the same study area during the period 2003–2005 (Figuerola et al. 2007a, 2008, López et al. 2010), indicating that WNV circulated in the common coot population in the different periods. Although reasons for this apparent susceptibility remain unclear, preference of this species for mosquito-rich habitats, its relative size, and the easier accessibility for feeding mosquitoes because of bald spots in the plumage of the head and long skinny legs might be implicated in this pattern (Figuerola et al. 2007b, Hubálek et al. 2008b). The WNV circulation detected in common coot in different countries (Figuerola et al. 2007b, Hubálek et al. 2008b) suggests that this species may be a good candidate for monitoring the WNV presence and circulation in a geographical area.
Overall seroprevalence of WNV in Spanish imperial eagle (3.0%) was lower than that previously found in central Spain (23.8%) during the period 2001–2005 (Höfle et al. 2008). The highest titers of neutralizing antibodies against WNV (1:80) were detected in this resident species, which indicates local WNV circulation. Höfle et al. (2008) found virus infection in Spanish imperial eagles, including healthy animals as well individuals with symptoms and lesions compatible with WNV. The Spanish imperial eagle is cataloged as “vulnerable” (IUCN 2008) and is considered to be the most endangered eagle in Europe. Therefore, we recommend periodic analysis of WNV in the health programs of this species.
Oral shedding of WNV was not found in any of the species examined. Taking into account that oral shedding of the virus is in general of less than 10 days (Komar et al. 2003), the possibility of detecting the virus in clinically healthy animals is low.
In conclusion, we evidenced the circulation of WNV and other flaviviruses of the JEV antigenic complex in wild and captive avian populations from southern Spain during the period 2007–2008. The presence of these flaviviruses could have implications in public health as has been recently demonstrated in Italy (Pecorari et al. 2009, Rizzo et al. 2009) and Greece (Papa et al. 2010). Further, our findings are relevant from an ecological point of view, as several of the birds seropositive are cataloged as “near threatened, “vulnerable,” or “endangered” by the IUCN. Long-term active monitoring and research programs need to be carried out to determine the precise role that wild bird species play in the epidemiology of the WNV in the Mediterranean context.
Footnotes
Acknowledgments
This work was supported by the Department of Environment of the Regional Government of Andalusia. The authors thank everyone involved in the Epidemiological Monitoring Program in Wildlife of the Regional Government of Andalusia for providing the valuable samples. The authors also thank Erika Serrano (CReSA) and José Miguel Ramírez (EGMASA) for their technical assistance.
Disclosure Statement
None of the authors of this article has a financial or personal relationship with other people or organizations that could inappropriately influence or bias the content of the article.