
Abstract
Longitudinal mosquito surveys were carried out in southern Portugal from 2004 to 2007, in a wetland area (Comporta, District of Setúbal) and around the perimeter of a dam irrigation plant that created the largest artificial lake in Europe, 250 km2 (Alqueva, Districts of Évora and Beja). Our aim was to study the diversity, abundance, and seasonal dynamics of mosquitoes, comparing these two different areas, to screen mosquitoes for West Nile Virus (WNV), an arboviral agent already detected in Portugal, because these areas are populated with abundant avian fauna. Monthly collections of adult mosquitoes were carried out by Centers for Disease Control light-traps with CO2 and by indoor resting collections. Mosquitoes were identified and screened for arboviruses by reverse transcriptase (RT)-polymerase chain reaction directed toward amplification of a 217-bp fragment of the NS5 gene. Mosquito peak densities were observed in July–August in Comporta and May–June, with a plateau in July–October, in Alqueva. However, densities were far higher in Comporta area (220,821 specimens) than in Alqueva area (9442 specimens), with a clear difference in species distribution, as in Comporta the predominant species was Culex theileri (85%), followed by Aedes caspius (6%), Anopheles atroparvus (4%), and Culex pipiens sensu latu (s.l.) (3%), whereas in Alqueva the predominant species was Cx. pipiens s.l. (56%), followed by An. atroparvus (18%), Cx. theileri (14%), and Culiseta longiareolata (9%). Female mosquitoes (8842 in 175 pools) of the species Ae. caspius, An. atroparvus, Culex mimeticus, Cx. pipiens Sensu latu (s.l.), Cx. theileri, and Culex univittatus were screened and found to be negative for WNV genomic RNA. Although there was no detection of WNV sequences in mosquitoes, vigilance should continue as the circulation of virus has been previously detected more than once in Portugal, in humans, animals, and mosquitoes, and in other surrounding Mediterranean countries.
Introduction
In Portugal, epidemiologic surveys for WNV in human and animal populations have been carried out since the 1970s, revealing low levels of seroreactivity for this virus (3% of 1649 humans examined by hemaglutination-inhibition assay) (Filipe and Andrade 1990). An epizootic of equine encephalomyelitis was recorded in southern Portugal before 1970 and 29% of the surviving animals had neutralizing antibodies against WNV (Filipe et al. 1973, Filipe and Andrade 1990). However, the first WNV isolation was reported in 1971 from Anopheles maculipennis s.l. mosquitoes (Filipe 1972). Two linked WNV cases were reported in tourists in the Algarve in the summer 2004 (Connell et al. 2004), after which WNV was isolated from Culex pipiens s.l. and Culex univittatus mosquitoes in the same region (Esteves et al. 2005). Phylogenetic analysis of whole genome sequences of these isolates strongly suggested two different introductions, in 1970 and 2004 (Parreira et al. 2007), supporting the idea that Portugal has the ecologic conditions for WNV reemergence as there are several wetlands that act as bird sanctuaries and also because of its location on the westernmost migratory bird route between Africa and Europe. In fact, recent serologic surveys in southern Portugal (1999–2002) revealed 12% positivity in birds and 3% in horses (Formosinho et al. 2006).
Throughout its worldwide distribution, WNV is maintained in nature by enzootic cycles between ornithophilic mosquitoes, predominantly Culex species and birds (Hayes et al. 2005). However, mosquitoes can act as bridge vectors, with the capacity to transmit WNV out of their natural cycle to accidental hosts, such as domestic animals, particularly horses, and to humans, leading to disease (Rappole et al. 2000, Rappole and Hubálek 2003, Linke et al. 2007, Kramer et al. 2008).
In temperate regions of Europe, WNV episodes occur in the late summer and early fall months, when bird migration is at its peak and mosquito populations are at their highest densities, and viral isolation has been achieved from actively migrating birds (Malkinson et al. 2002). WNV outbreaks in humans can occur at urban sites near wetlands where migratory birds, ornithophilic mosquitoes, and humans are concentrated (Rappole et al. 2000). The presence of wetlands and lagoons, used by migratory birds, can favor new viral introductions. The associated presence of suitable breeding sites for mosquitoes and favorable climatic conditions facilitate the abundance of mosquito populations, which are vectors of WNV, and can thus maintain viral enzootic cycles (Medlock et al. 2006).
Comporta is a coastal wetland area located in the estuary of the river Sado with salt marshes and large irrigated rice farming fields, which on the whole constitute favorable mosquito breeding sites. Such areas can have an impact on mosquito-borne arboviral diseases as has been documented elsewhere (Keiser et al. 2005b).
Dam construction and new irrigation systems generally imply ecosystem disturbances, such as changes in mosquito populations (Tubaki et al. 2004), and can have negative health impacts on bordering communities, particularly if the associated irrigation schemes are not properly managed associated with vector-borne diseases, such as malaria or arboviruses (Tetteh et al. 2004, Keiser et al. 2005a). Alqueva dam is a major multipurpose water resource project (water storage, irrigation, energy production, recreation, and river Guadiana flow control). Filling of the reservoir started in February 2002, but irrigation canals and subsidiary dams are still under construction (EDIA, 2009). Because of its dimensions the Alqueva dam created the largest artificial lake in Europe, 250 km2, and consequently, conditioned profound changes occurred in the region's landscape and natural environment, creating an “artificial wet habitat” in an originally inland dry area (thousands of hectares flooded, over 1 million trees cut). Both areas have a large avian fauna, as well as natural mosquito populations, and thus can harbor WNV reservoirs.
The study of the seasonal dynamics of mosquito vectors is an important contribution for the surveillance of arboviral diseases (White 2001, Medlock et al. 2006). Evaluation of both biological and ecological data is cornerstone of mosquito control programs in a public health perspective (Rydzanicz and Lonc 2003, Şimşek 2004).
The goals of this work were to determine the abundance, diversity, and seasonal dynamics of mosquitoes in two different areas of southern Portugal, a coastal wetland area, Comporta, and an inland area in the process of an ecological change due to a dam construction, Alqueva, as well as to study the possible influence on the mosquito fauna of this region, and as both these areas have abundant avifauna, to screen these mosquitoes for WNV, which has already been repeatedly detected in Portugal.
Materials and Methods
Study area and design
Longitudinal mosquito surveys with monthly collections were carried out in a wetland area, area 1—Comporta, District of Setúbal, Portugal, 38°22′60N, 8°46′60W, in the years 2004–2005, and in the perimeter of a dam irrigation plant, area 2—Alqueva, Districts of Évora and Beja, Portugal, 38°17′N, 7°25′W, in the years 2004–2005 and 2007 (Fig. 1).
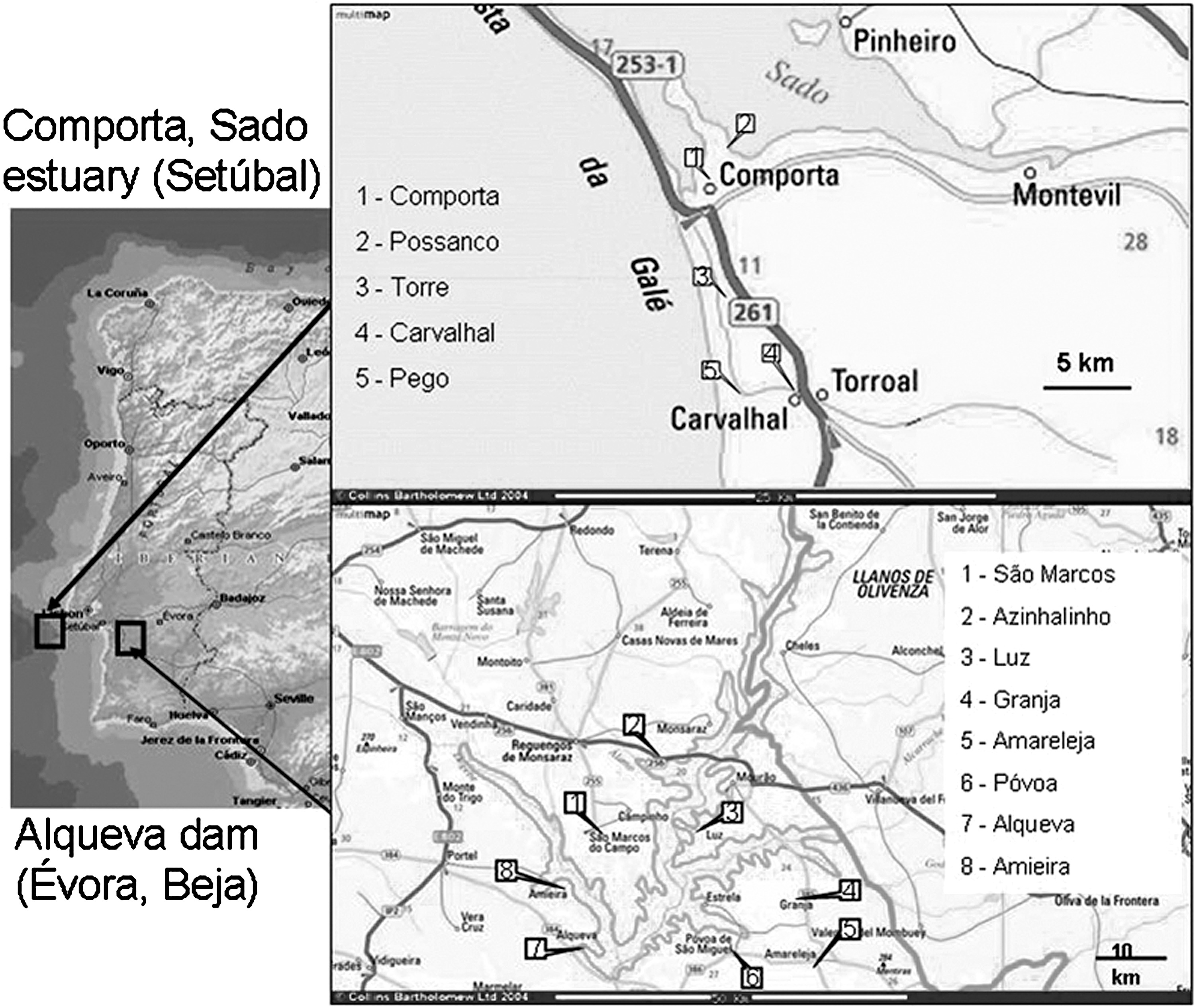
Map of the Iberian Peninsula with Portugal and the insets showing the study areas: Comporta, Sado estuary (Setúbal) and Alqueva dam (Évora, Beja).
Area 1, Comporta, is a low-lying area (altitude <60 m) with diverse ecotypes, consisting of a coastal strip 20 km in length and 5–10 km in width, with marshland and abundant rice cultivation. Residential areas are situated along a national road that crosses the study region from north to south. The south and east are mainly occupied by pine forest (Pinus pinaster A., Pinus pinea L.) and seminatural agroforest systems of cork-oak (Quercus suber L.). In the west there are extensive areas of rice fields and a system of sand dunes. The north and northwest are parts of a protected landscape area occupied by marshland, rice fields, and saltpans. This protected area extends northward into the national wildlife reserve of Estuário do Sado. The reserve harbors over 240 bird species. These include migratory birds such as the European starling (Sturnus vulgaris L.), the mallard (Anas platyrhynchos L.), and the white stork (Ciconia ciconia L.), which have been reported as WNV hosts (Rappole and Hubálek 2003).
The region has a humid subtropical mid-latitude climate (class C, Köppen Classification System), subtype Mediterranean, with a dry warm summer and a mild winter. Monthly averages of mean daily temperatures vary between 10°C and 21°C and relative humidity between 76% and 89%. Monthly averages of daily rainfall fluctuate between 0.12 and 3.4 mm.
Area 2, Alqueva, corresponds to the perimeter of the Alqueva dam reservoir, spreading for approximately 994 km2. It is a traditionally inland dry area where a dam irrigation plant was constructed and started to be filled in 2002, creating the largest artificial lake in Europe, 250 km2. Altitude ranges from 140 to 250 m, and the main ecotype is a seminatural agroforest landscape system of cork-oak (Q. suber L.), olive trees (Olea europaea L.), and vineyards (Vitis vinifera L.), with small and disperse residential areas. Avian fauna is also abundant, including the white stork (C. ciconia L.), black stork (Ciconia nigra L.), and other migrating or semimigrating birds. The region has Mediterranean climate and moderate humidity, with dry and hot summers (class Csa, Köppen Classification System). Monthly averages of mean daily temperatures vary between 9°C and 24°C and relative humidity between 31% and 83%. Monthly averages of daily rainfall fluctuate between 0.10 and 2.89 mm.
Mosquito sampling and processing
Adult mosquitoes were collected monthly, in fixed sampling sites, five for area 1 and eight for area 2, with (i) Centers for Disease Control (CDC) traps (Sudia and Chamberlain 1962) baited with CO2 (ca. 1 kg/trap) (Newhouse et al. 1966) for a minimum of 12-h periods covering sunset to sunrise, and (ii) indoor resting (IR) mosquito collections with the aid of battery-powered hand aspirators, mainly in animal shelters; collected mosquitoes were transported in refrigerated boxes to laboratory where they were frozen, identified (Ribeiro and Ramos 1999), pooled according to site, species, sex, and females by gonotrophic stage, and kept at −80°C until viral screening.
Viral screening
Mosquito pools were ground with glass beads and alumina (Huang et al. 2001) to detect the presence of Flavivirus. Mosquito homogenates were used in a random primed reverse transcription reaction carried out with Moloney Murine Leukemia Virus (MMLV) RT (Invitrogen, Paisley, UK), and the RT products subsequently were taken as templates for heminested-polymerase chain reactions performed with puRe Taq Ready-to-Go PCR Beads (Amersham Biosciences, Little Chalfont, Buckinghamshire, United Kingdom). Primer sets and polymerase chain reaction conditions for amplification of a 217-nucleotide fragment from the flavivirus NS5 coding sequence were previously detailed (Scaramozzino et al. 2001, Esteves et al. 2005). Amplification products were analyzed on 1.5% agarose gels containing ethidium bromide.
Statistical analysis
Mosquito density per region was calculated as the arithmetic mean of collections performed in all sites of each region, analyzed separately for CDC and IR methods and according to collection effort measured in time (trap-night for CDC and per person per hour for IR).
Data were analyzed for normality with the Kolmogorov–Smirnov and Shapiro–Wilk tests and for homogeneity of variance with Levene's test. Whenever raw or logarithmic converted data do not meet normality and homogeneity of variance, nonparametric tests (Mann–Whitney U-test for two samples and Kruskal–Wallis test for more than two samples) were carried out (Siegel and Castellan 1988). The statistical package SPSS (Chicago, IL) 14.0® for Windows was used.
Results
Mosquito peak densities were observed in July–August in Comporta and May–June, with a plateau in July–October, in Alqueva (Fig. 2). However, peak densities were far higher in Comporta, with over 6000 mosquitoes per trap-night and 600 mosquitoes per collector per hour by IR, whereas in Alqueva area the CDC densities reached 70 mosquitoes per trap-night and IR collections reached 300 mosquitoes per collector per hour.
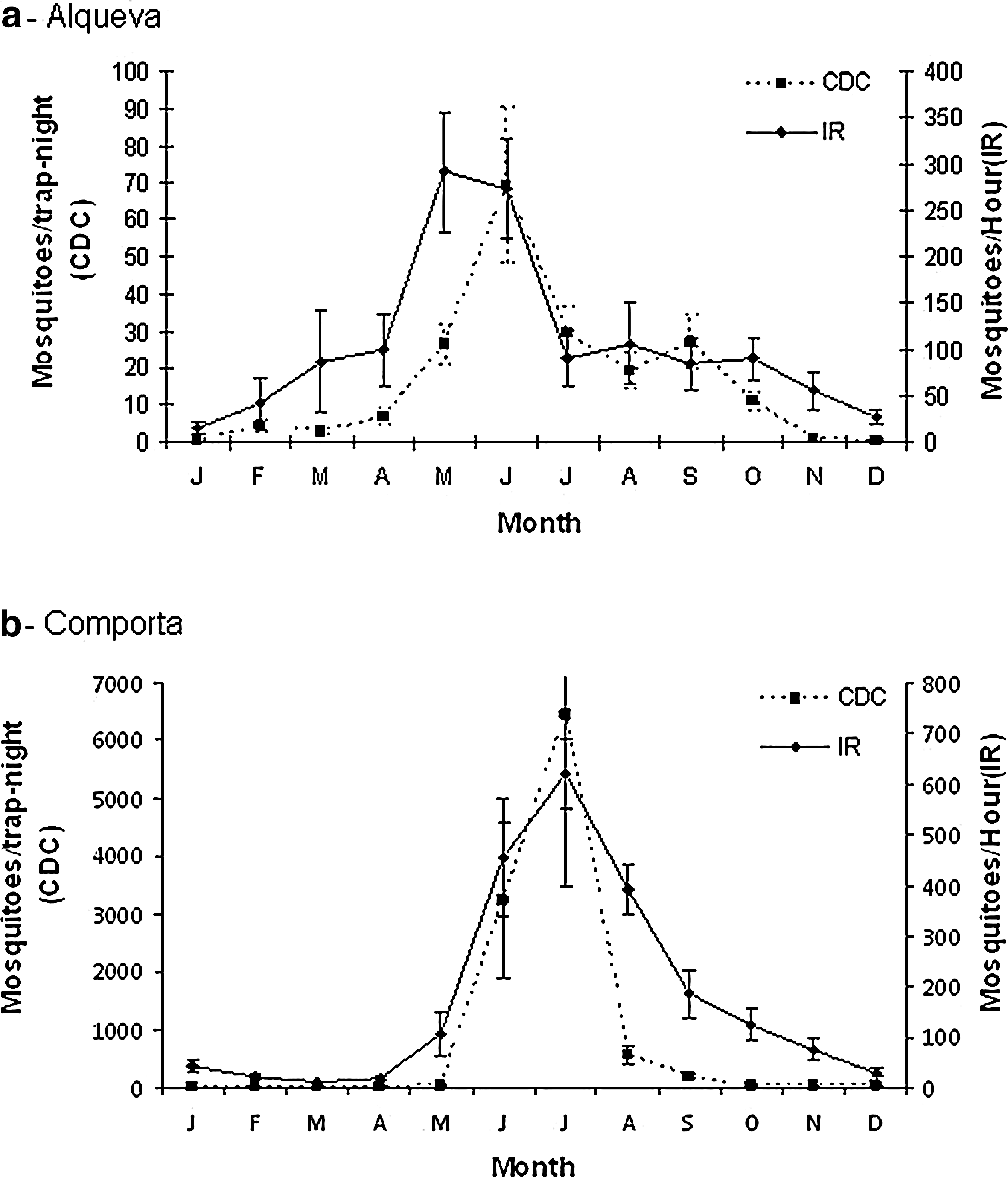
Mosquito seasonal dynamics from study areas: (
Morphological identification of 220,821 adult specimens from Comporta and 9442 from Alqueva confirmed the presence of 9 species in the former and 13 species in the latter (Table 1). Species distribution was markedly different in these two areas, with Culex theileri (85%), followed by Aedes caspius (6%), Anopheles atroparvus (4%), and Cx. pipiens s.l. (3%) as the predominant species in Comporta, whereas Cx. pipiens s.l. (55%), followed by An. atroparvus (18%), Cx. theileri (14%), and Culiseta longiareolata (9%) as the predominant species in Alqueva (Table 1).
N ID, nonidentified specimens.
The relative mosquito species distribution differed also according to collection method and study area (Fig. 3).
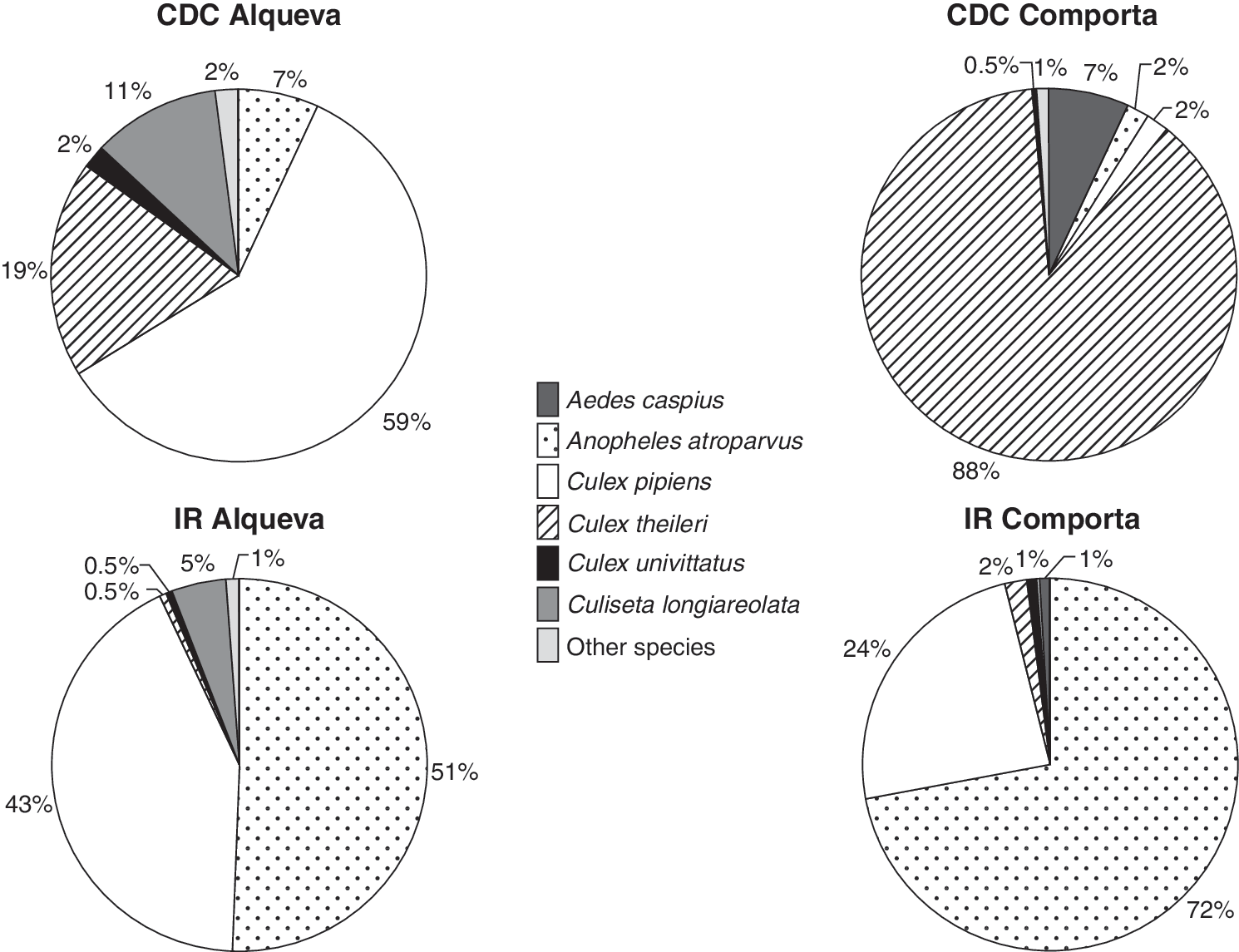
Relative mosquito species distribution according to collecting method (CDC traps [CDC] or IR) in the Alqueva and Comporta areas of southern Portugal.
From a total of 8842 female mosquitoes screened (4% of the total mosquitoes collected) there were no detection of WNV RNA sequences (Table 1).
Discussion
Fourteen of the 40 mosquito species known in continental Portugal (Ribeiro et al. 1988) were collected in this survey and are in agreement with the distribution found in previous whole country cross-sectional surveys (Almeida et al. 2008). The relative species distribution in these two study areas was clearly different, in agreement with the differences in habitat types. Comporta, a wetland area with extensive marshes and rice fields at the estuary of the Sado river, exhibited a mosquito fauna where Cx. theileri is the predominant species, followed by the halophilic Ae. caspius, thus expectable in this south European habitat. Cx. theileri breeds abundantly in the extensive rice fields, reaching 60,000 mosquitoes per trap-night in some of the sampling localities between July and August, whereas Ae. caspius breeds not only in the salt marshes but also in the rice fields. Both these species reveal a compatible behavior to this man-made habitat resulting from this farming activity (Novo 2008). Together, the natural and the man-made habitats are responsible for the high mosquito densities in this area, which peak when climatic conditions and the abundance of larval habitats are the most favorable.
Alqueva dam has conditioned profound changes in the natural landscape and habitats of that inland area, with the creation of the largest artificial lake in Europe. However, the mosquito fauna therein collected was essentially similar in diversity of species to previous collections (Pires et al. 1982). However, it is not possible to conclude if mosquito densities have changed, because longitudinal surveys had not been conducted in this area before the dam construction. The predominant species were Cx. pipiens s.l., An. atroparvus, Cx. theileri, and C. longiareolata, which were related to the type and abundance of breeding habitats, mainly composed of peridomestic and agricultural containers, and small- to medium-sized ground pools and streams, but mosquito breeding was seldom found in the margins of the dam lake (Almeida et al. in preparation).
Aedes albopictus, already established in neighboring Spain (Aranda et al. 2006), was not found in this survey, neither in adult collections nor as immature despite the large amount of breeding habitats surveyed (Almeida et al. in preparation).
The relative proportion of mosquito species varied according to collection method and area. Although IR collections were not so markedly different between the two areas, CDC collections were strikingly different between the two areas because of the nearly absolute dominance of Cx. theileri in Comporta (88%). An. atroparvus constituted the majority of mosquitoes collected resting inside animal shelters (51–72%) but had low proportions (2–7%) in CDC traps, in both areas, in agreement with previous surveys (Almeida et al. 2008). Cx. pipiens had similar proportions in either CDC or IR in Alqueva (59–43%), was lower in IR in Comporta (24%), and was hardly present in CDC in Comporta (2%). The differences in IR collections between the two areas, regarding An. atroparvus and Cx. pipiens, are likely attributed to the abundant breeding of the former in the wide area of rice fields of Comporta (Novo 2008) and the nature of the breeding biotopes in Alqueva, mainly peridomestic and agricultural containers, more suitable for the latter (Almeida et al. in preparation). Ae. caspius was present only in CDC traps in Comporta, hardly appearing in IR collections, which reveals its predominantly exophilic behavior.
Cx. theileri was the predominant species in Comporta area, because of its association with the rice farming activity. In fact, it was most abundantly found in southern Portugal, particularly in the river estuaries of Tagus and Sado, with maximum densities in rice farming areas (districts of Santarém and Setúbal) (Anonymous 2001, Almeida et al. 2008). This species is a competent vector of Rift Valley Fever, WNV, and Sindbis virus (Jupp et al. 1972, McIntosh et al. 1980, Jupp 1985). Further, its public health importance is heightened by its nuisance as it can reach 400 bites per person per hour in Comporta area (Sousa 2008). The impact of irrigated rice culture has been documented on the incidence of arboviruses such as Japanese encephalitis elsewhere (Keiser et al. 2005b).
Ae. caspius, the second most abundant species in Comporta, is present in coastal and wetland areas such as estuaries and marshlands, with highest densities found in southern Portugal, particularly in the river estuaries of Tagus (district of Santarém), Sado (district of Setúbal), and Ria Formosa (district of Faro) (Almeida et al. 2008). It is an aggressive species of opportunistic behavior (Cambournac 1944, Rioux 1958), reaching in Comporta as much as 300 bites per person per hour in twilight periods during the summer months (Sousa 2008). It is a competent vector for Rift Valley Fever (Turell et al. 1996), Tahyna (Lundstrom 1999), Chikungunya virus (Vazeille et al. 2008), and WNV (Hubalek and Halouzka 1999), although French populations of this species revealed low competence for WNV (Balenghien et al. 2008).
Cx. pipiens s.l. is abundant in both areas and is actually the mosquito that presented the highest mean densities in the whole country (Ribeiro et al. 1988, Almeida et al. 2008). It was collected by CDC traps, IR, and also by human baited collections, evidencing a considerable degree of anthropophily (Almeida et al. 2008, and this work). This species complex includes ornithophilic and anthropophilic forms, of varying degrees, which can act as bridge vectors with a higher potential for spreading zoonotic viruses such as WNV (Fonseca et al. 2004), highlighting its public health importance. Their role as vectors of WNV has been thoroughly documented in both the Old World (Hubalek and Halouzka 1999) and the New World (Anderson et al. 1999). In Portugal, particularly in the Comporta region, both the pipiens and the molestus forms have been found in sympatry with considerable degree of introgression, further heightening their importance as bridge vectors (Gomes et al. 2009).
An. atroparvus is the only member of the An. maculipennis complex found in southern Portugal (Ribeiro et al. 1988, Sousa 2008) and in considerable densities, particularly in IR captures in animal shelters (Almeida et al. 2008).
Previous WNV mosquito infections in Portugal have been detected from An. atroparvus (Filipe 1972) and from Cx. pipiens and Cx. univittatus (Esteves et al. 2005). The first isolation, in 1971, was in an area relatively close to the Alqueva dam, the Roxo dam also in the southern province of Alentejo (Filipe 1972). Coincidently, the first WNV neuroinvasive disease in Spain was registered in 2007 in a neighboring area (Province of Badajoz) (Kaptoul et al. 2007). Although in our survey no mosquito infections were detected, this should not be a deterrent from continuous entomologic surveillance. Both these areas have large avifauna, including several migrating species. Serosurveys in these areas (1999–2002) revealed 12% positivity in birds and 3% in horses (Formosinho et al. 2006) and recent reintroductions have been documented (Parreira et al. 2007). In fact, WNV has been circulating in the Mediterranean area in the last decade (Zeller and Schuffenecker 2004), and in the last 2 years it has caused several human cases of considerable gravity, with casualties, in Italy (Barzon et al. 2009, Rizzo et al. 2009).
Footnotes
Acknowledgment
This work was funded by project “RARIMOSQ” (ref. 35-60624-S), Fundação Calouste Gulbenkian, Portugal. “The research leading to these results has received funding from the European Community's Seventh Framework Programme (FP7/2007-2013) under Project Arbo-zoonet, grant agreement number 211 757.”
Disclosure Statement
The authors declare no conflict of interest.
This work was presented at the First Annual Arbo-Zoonet Annual Meeting, St. Raphael, France, 2009.