
Abstract
A Rift Valley fever (RVF) outbreak occurred in Madagascar from January to May 2008. The objectives of this study were (1) to assess the current and past circulation of RVF virus (RVFV) in livestock in Madagascar and (2) to evaluate the extent and magnitude of the 2008 RVF outbreak in livestock. The results of a country-wide serosurvey conducted in August 2008 on small and large ruminants are reported here. The study included 3437 cattle and 989 small ruminants (227 sheep and 762 goats) sampled in 30 of the 111 Malagasy districts, selected to be representative of the different ecozones and livestock density areas. Sera of animals were tested for the detection of immunoglobulins M (IgM) and G (IgG) against RVFV using commercial enzyme-linked immunosorbent assays kits. Recent infections (presence of IgM against RVFV) were detected in only 9 cattle (0.3% [0.1–0.4]) and 33 small ruminant (3.3% [2.2–4.5]) samples. Past infections (presence of IgG and absence of IgM against RVFV) were detected in 887 cattle (25.8% [24.3–27.3]) and 244 small ruminant (24.7% [22.0–27.4]) samples. Past infections were detected in all sampled sites. All ecozones were affected. In the southern and northwestern areas, the prevalence of cattle showing evidence of past infection with RVFV increased with the age of the animals. Our results suggest that there has been country-wide circulation of RVFV in 2008 in Madagascar, including in parts of the country where no clinical illness, either in animals or in humans, was reported. The data also suggest that the southern and northwestern areas may be endemic for RVFV, and that the virus may spread when ecological conditions are favorable for its amplification.
Introduction
More than 70% of Malagasy people live in rural areas, and are highly reliant on animal traction, livestock products, and livestock trade for their livelihoods. In 2004–2005, there were 9,500,000 cattle, 1,219,000 goats, and 695,000 sheep in the country (Recensement de l'Agriculture 2007). The presence of RVF was first demonstrated in Madagascar in March 1979, during the rainy season (November–April), when RVFV was isolated from mosquitoes collected in the moist-tropical primary forest of Perinet, Moramanga district (120 km east of the capital, Antananarivo) (Fontenille 1989). Then, in April 1990, RVFV was identified as responsible for a significant wave of abortions in cattle in the Fenoarivo Atsinanana district on the eastern coastal plain. Fifteen suspected human cases were tested in hospitals, of which five were confirmed and one died. RVFV seroprevalence among cattle owners in the area where livestock abortions were recorded reached 8%, a large majority of whom were young men. Seroprevalence of cattle in the same area was 26% (Morvan et al. 1991a, 1991b). The following year, from February to April 1991, high rates of abortion in cattle were reported in the central highlands, around Antananarivo, and fatal human cases were confirmed (Morvan et al. 1992a, 1992b, 1992c).
RVF outbreaks dramatically affected countries in the Horn of Africa (Kenya, Somalia, and the United Republic of Tanzania) in late 2006, through the first half of 2007, and in Sudan in September 2007. Southern African countries (Swaziland and South Africa) and islands in the Indian Ocean (Union of the Comoros and Mayotte) were affected in 2007 and 2008 (World Health Organization 2007a, 2007b, Bird et al. 2009, Sissoko et al. 2009). In Madagascar, the disease reemerged during the rainy season (January) 2008 and severely affected the human and livestock populations. The exact extent of this epizootic is still unclear because of limited reporting and limited sampling (Andriamandimby et al., 2010).
We report here the results of a national serosurvey performed among ruminants sampled in the months following the 2008 Malagasy outbreak, as part of efforts to retrospectively evaluate the extent and magnitude of the RVF 2008 outbreak among ruminants and to estimate the past circulation of RVFV in the livestock of Madagascar.
Materials and Methods
Sampling
A country-wide cross-sectional survey was conducted among livestock (cattle, sheep, and goats). To optimize the sampling strategy, two factors of stratification, type of ecozone and cattle density, were selected for their likelihood to impact on distribution of RVF at the country level. In Africa, Davies (1998) linked the suitability for RVF occurrence and the potential activity of the virus with the main agro-climatic ecozones described in Africa (Pratt et al. 1966). This relationship can be schematically summarized as follows: (1) enzootic activities (+/−cryptic) in high rainfall forests and forest edges; (2) periodic increased virus activity to epizootic proportions in moist bushed and wooded grasslands of moist areas; and (3) rare but explosive epizootic RVFV activities in dry grassland and semiarid zones associated with flood plains. A map of the different ecozones for Madagascar is given in Figure 1, and while not perfectly fitting with the classes of Davies (developed for Kenya), they have been used to explore potential differences in disease transmission. In 2004–2005, the Ministry of Livestock carried out a cattle census. The highlands and the western and southern parts of Madagascar hosted the highest densities of cattle (Recensement de l'Agriculture 2007). Using these data, areas classified by cattle density <9 cattle/km2, 9–18 cattle/km2, and >18 cattle/km2were demonstrated.
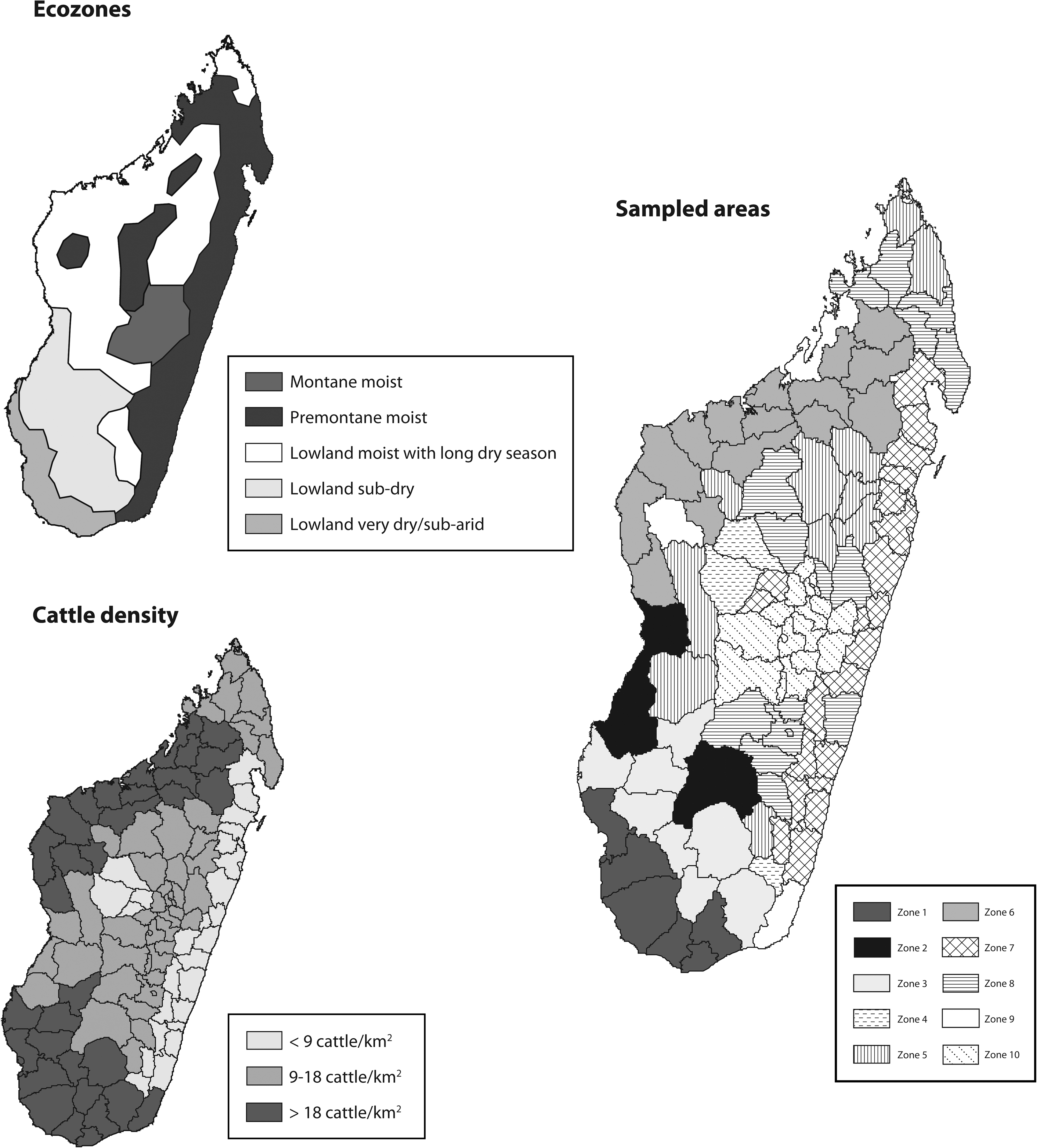
Data layers for stratification and resulting subdivisions of Madagascar for livestock sampling. The definition of ecozone is extracted from the classification of UNEP/WCMC; statistics on cattle distribution are derived from the national livestock census 2005.
Finally, the country was divided into 10 sampling areas representing the combination of the two stratification factors (Fig. 1). Sample sizes were calculated using WinEpiscope software, based on an expected prevalence differing between the sampling areas (from 10% to 40%, the prevalence expected to be higher in ecozones known to be favorable). As a result, sample sizes ranged from 220 to 580 individuals, and only animals that were born and lived in the same sampling area were considered in the survey. Animals were sampled by veterinarians in the local slaughterhouses, on farms or at marketplaces. Sampling on small ruminants was limited to four high-density districts because of a limited geographical distribution of these animals in the country.
A standardized questionnaire was used, including an identification number for the sample, information on the species, gender and age, origin of the animal sampled, and the place of sampling. Blood samples were collected in a 5 mL plain tube and were left for coagulation and centrifuged within 72 h after collection. Sera were transferred in individual, identified microvials, kept under cold conditions (max 4°C), and sent to the Laboratoire National de Diagnostic Vétérinaire in Antananarivo.
RVFV serological assays
At Laboratoire National de Diagnostic Vétérinaire, commercial sandwich enzyme-linked immunosorbent assay for the detection of anti-RVFV IgG and capture enzyme-linked immunosorbent assay for the detection of anti-RVFV IgM were used (Paweska et al. 2003). In the IgG test, cattle and small ruminants sera producing percentage positivity (PP) values ≥30 and 25, respectively, were considered to be positive, and below values to be negative. In the IgM test, sheep, goats, and cattle sera producing PP values ≥8, 9.5, and 14.3, respectively, were considered to be positive, and below values to be negative. The tests were performed according to the manufacturer's instructions (Biological Diagnostic Supplies Limited, Dreghorn, United Kingdom).
IgG can persist for months and even years after infection and is therefore used as a reliable indicator of a past infection by the virus. In contrast, the persistence of IgM is low. A previous study in Madagascar established that 27% of the RVFV-infected cattle still had IgM against RVFV 2 months after natural infection, 15.9% after 3 months, and only 3.6% after 5 months (Morvan et al. 1992c). To our knowledge, there are no data available on duration of IgM in small ruminants sera. However, we considered that the presence of IgM against RVFV in serum of livestock was indicative of recent infections, while presence of IgG against RVFV and absence of IgM against RVFV were indicative of past infections.
Data analysis
All the data were compiled in a unique Microsoft Office Access 2007 database. Statistical analyses were performed using STATA/SE 10.0 software. Different tests were used depending on the nature and distribution of the data, including the Mann–Whitney test, a nonparametric test for trends across ordered groups, and the chi-square test.
Results
Samples
In line with the sampling protocol, a total of 3450 cattle and 989 small ruminants (227 sheep and 762 goats) from 30 of the 111 districts of Madagascar were sampled. The survey was conducted over a limited period (August 2008) to assure consistency between the results. Of the 3450 cattle sampled, 3437 met the criteria to be tested.
The range of age in the cattle population was from 4 months to 19 years, with a median age at 5 years and a mean at 5.57 (117 missing data). The range of age in the small ruminant population was from 6 months to 9 years, with a median age at 2 years and a mean at 2.4 years (245 missing data). The sex ratio (male/female) was 1.4 (30 missing data) and 0.4 (5 missing data) in cattle and small ruminants, respectively.
Twelve percent of cattle were sampled in slaughterhouses, 64% on farms, and 14% in markets. Most of the small ruminants (98%) were sampled on farms.
Prevalence of past infections (presence of IgG and absence of IgM against RVFV)
Past infections were detected in 887 cattle (25.8%) and 244 small ruminants (24.7%) (Table 1). PP values of negative and positive sera of small ruminants were well differentiated, whereas the difference was less perceptible for cattle sera. However, a negative exponential distribution of PP values was observed for the negative sera (Fig. 2). Past infections were observed in all areas, confirming a wide circulation of RVF (Table 1 and Fig. 3). In most of the areas, these prevalences in cattle were between 15% and 35%, with lowest values in the south of the country. When considered at the district scale, prevalence of the past infections in cattle and small ruminants ranged from 5.5% [3.2–7.7] to 52% [45.8–58.3] and from 0.4% [0–1.2] to 60.1% [54.0–66.2], respectively, highlighting high circulation areas in the country.
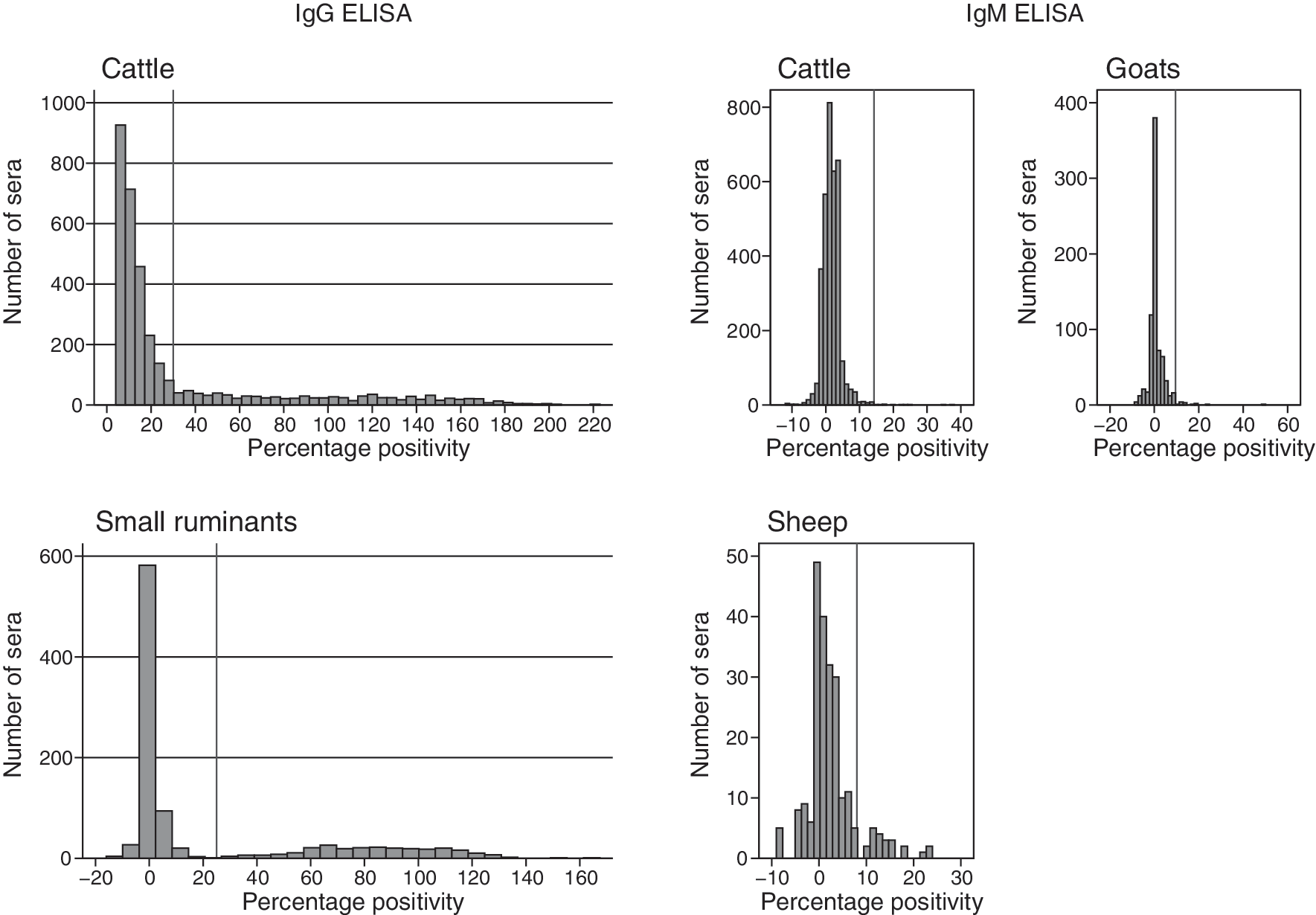
Distribution of percentage positivity values in cattle and small ruminants sera, Rift Valley fever immunoglobulin G (IgG) enzyme-linked immunosorbent assay (ELISA), Madagascar, August 2008 survey. The vertical reference line on X-axis indicates the percentage positivity threshold value.
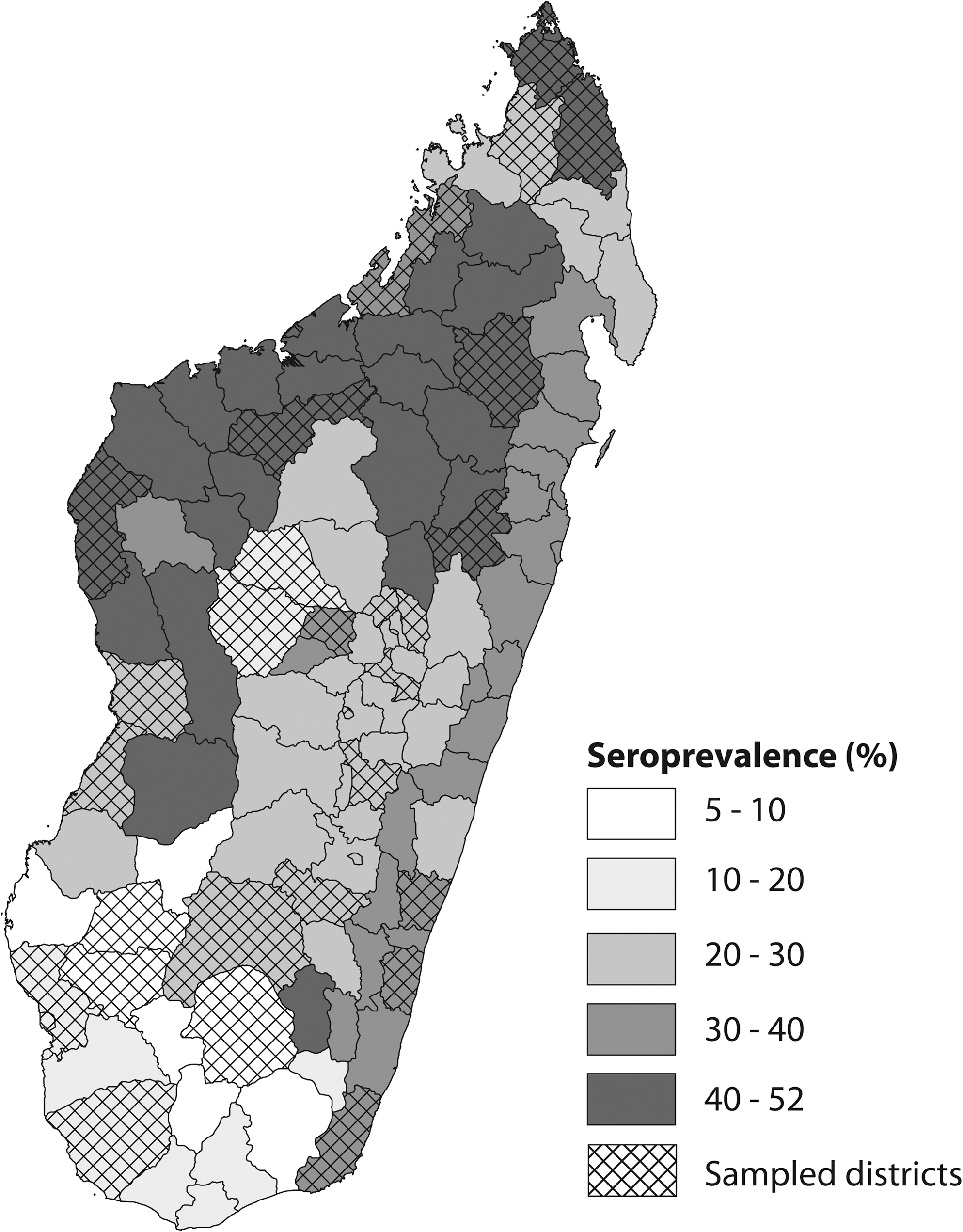
Prevalence of past Rift Valley fever virus infections in cattle, Madagascar, August 2008 survey.
The prevalences are given in percentage, with limits of the 95% interval of confidence given in parentheses.
The prevalence of past infections was significantly higher in males than in females in cattle (p = 0.001) and in small ruminants (p < 0.001). Prevalence in cattle was lower when the animals were sampled on farms (p < 0.001). It increased with the age of the animals (p < 0.0001). When further explored, it appeared that this difference is mainly driven by the results obtained in the southern and northwestern areas. Prevalence in small ruminants decreased with age (difference only driven by the results obtained in Morondava). Because of disequilibrium in the sampling for small ruminants, the link between prevalence and place of collection could not be tested.
Prevalence of recent infections (IgM against RVFV)
Recent infections were detected in 9 cattle (0.3%) and 33 small ruminants (3.3%) (Table 1). The distribution of PP values of the sera around the threshold was not very informative about the robustness of our test, because of the low number of positive sera (Fig. 2). Positive cattle samples originated from 4 of the 10 selected areas (Fig. 4), whereas for small ruminants, recent infections were detected in the four districts investigated (Table 1). The maximal prevalence of RVF recent infections per area was 1.1% (0–2.1) in cattle, and no significant difference between age (p = 0.32), sex (p = 0.618), and place of sampling (p = 0.989) was observed (Table 2). The prevalence of recent infections per district ranged from 5.9% (0–12.4) to 10.6% (5.6–15.5) in sheep and from 0.8% (0–1.9) to 3.3% (0–6.9) in goats. No significant difference between sex or age was observed (p = 0.936 and 0.966, respectively). Because of disequilibrium in the sampling for small ruminants, the link between prevalence and place of collection could not be tested.
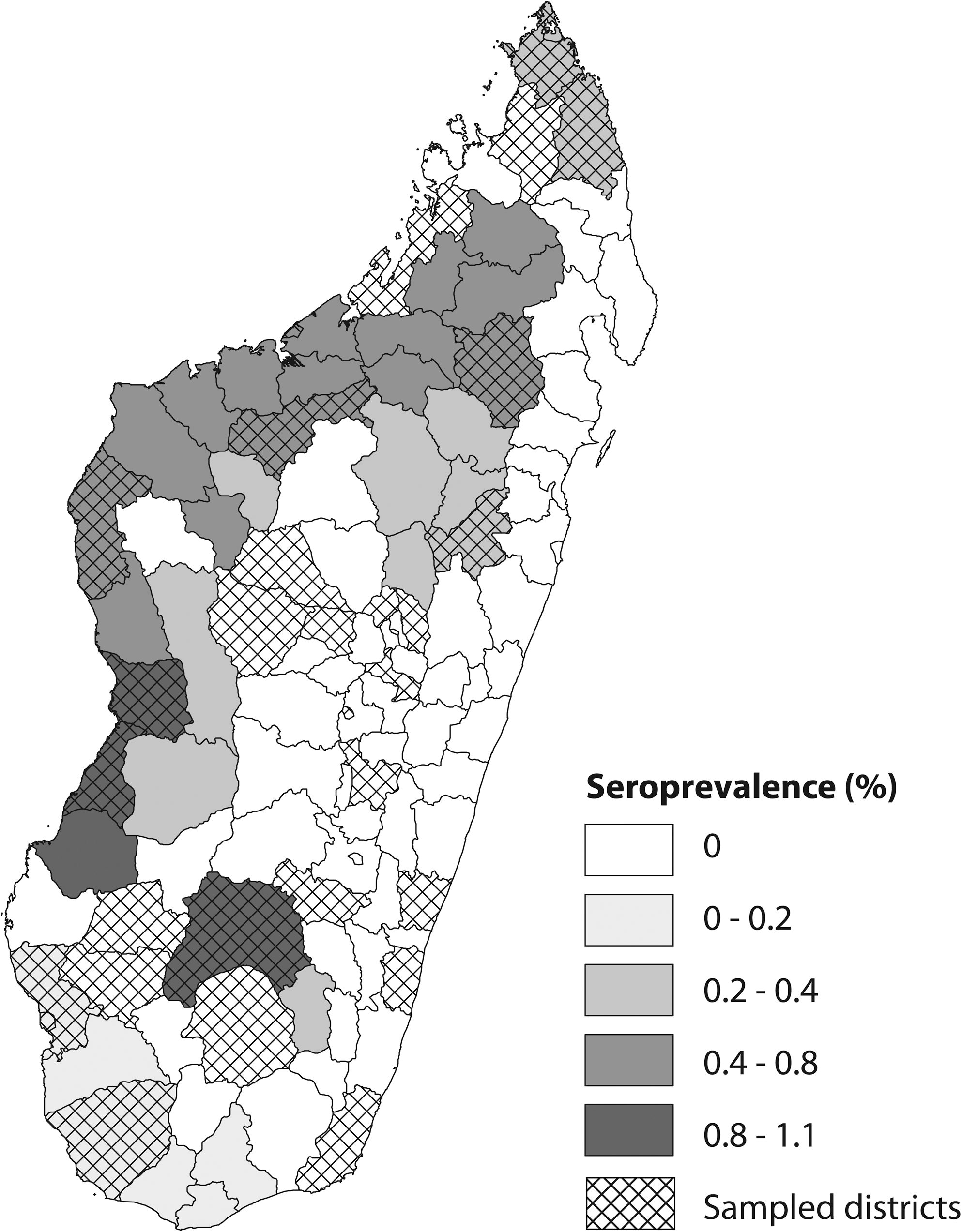
Prevalence of recent Rift Valley fever virus infections in cattle, Madagascar, August 2008 survey.
Of the 42 small ruminant and cattle samples positive for IgM, 25 were IgG negative. These samples were collected in the southern and northwestern districts.
Differences between factors of stratification
A significant difference in the prevalence of past infections in cattle was observed between ecozones (p < 0.001), with higher values in the moist lowland and premontane areas, but arid and dry ecozones also hosted a significant number of infected animals (Table 3). A significant difference in the prevalence of past infections was also observed between the classes of cattle density, with the intermediary being the most infected. Because of the limited number of positive samples, results with recent infections were not convincing either for ecozones (p = 0.153) or for cattle density (p = 0.437).
Discussion
The results of this cross-sectional country-wide serosurvey in livestock suggest that RVFV has circulated in all the regions of Madagascar. These results complement those of a postoutbreak serological survey conducted in humans over recent months (Andriamandimby et al., 2010). In this late study, no evidence of RVF in humans was found in some of the southern districts, while our results confirm that RVFV has circulated in livestock in some of these districts (namely, Midongy-Atsimo, Vangaindrano, Ampanihy, Sakaraha, Betioky, and Toliara-I).
Animal infections have been documented in all types of ecozones. Whereas the persistence and the circulation of the virus may differ between these ecozones, such a wide distribution of serological results has already been observed during large outbreaks. During the 2006–2007 epizootic in Kenya, infected livestock was reported in all types of ecozones, including semidesert grassland and shrubland in the north of the country, the acacia bushland and savannahs in the south, and the montane and moist forests on the coast (National Geographic 2001, Bird et al. 2008). Based on the hypothesis that the virus can survive in desiccated Aedes spp. eggs during interepidemic periods, sites of possible endemicity have been described in Kenya, and typical habitats, namely, dambos, have been identified (Linthicum et al. 1985). The virus is thought to reemerge when climatic conditions favor the flooding of these habitats, and from there disseminate and colonize other ecozones, mainly through the movements of livestock (Davies 1975, 1985). The increase of past infection prevalence with age in the southern and northwestern areas suggests that virus transmission occurs annually. Indeed, with a sub arid climate and spiny thickets (National Geographic, 2001), the ecozone in the south is very similar to those known to host low levels of endemic transmission of RVF in East and West Africa (Davies 1998), whereas the northwestern area is characterized by mangroves and dry deciduous forest (National Geographic 2001). This hypothesis is also supported by the results of a cross-sectional country-wide serosurvey performed on livestock in 1996, when six of the seven RVF IgM-positive animals (from among 9614 sampled) originated from the southern area, indicating circulation of the virus during an interepizootic period (Zeller, unpublished data). Consequently, the southern and the northwestern areas may be endemic areas for RVFV, from where the virus may spread when ecological conditions are favorable for its amplification. However, because partial sequences of 2006–2007 Kenyan and 2008–2009 Malagasy RVFV isolates were close (Andriamandimby et al., 2010), we cannot exclude repeated introductions of RVFV from Africa in these two areas.
Interestingly, the prevalences were low in the sub-arid and arid lowlands of the south and increased gradually in the central, temperate highlands, in the very moist lowlands of the eastern coast, and in the moist lowlands of the northwest region. This observation is not inconsistent with the previous hypothesis if we attribute these lower prevalences to the reduced period of transmission because of climatic conditions limiting mosquito populations, whereas the transmission occurs all year long in the northwest moist ecozone, as described in Kenya (Labeaud et al. 2007). In contrast, long-lasting RVF outbreaks in animals and humans can be observed following the introduction of viraemic animals in rice field production systems, where several species of water-dependant mosquitoes known to be efficient vectors for RVF can develop almost throughout the year, such as Culex and Anopheles spp. (Swanepoel and Coetzer 2004). These rice production fields are very abundant in the moist ecozones of Madagascar.
Animal transportation for trade has probably played a major role in the spread of the disease in Madagascar. Livestock from the southern breeding areas are transported on boats from the port of Tulear to different destinations in Madagascar. Significant numbers of animals also reach the slaughterhouses around Antananarivo by road (Fieloux and Lombard 1987). Other types of movement are also known, such as the transport of stolen cattle from the northwest to the northeast of the country (Peter Fenozara, pers. comm.). Consequently, RVFV could be transferred from these possibly endemic areas to other parts of the country in a very short period, via viremic animals.
Evidence of recent infection was detected in only 42 of the 4425 tested animals. However, the duration of the IgM antibody against RVFV in cattle is known to be relatively short. In this study, blood sampling was carried out 3 months after the last notification of a confirmed case (Andriamandimby et al., 2010), and it is probable that on this date most of the animals no longer had IgM against RVFV. It remains that 24 sera tested positive for IgM against RVFV and IgG negative, suggesting that the virus was still circulating during the dry season, especially in the south and in the northwest part of the country. Finally, the global prevalence of IgG against RVFV observed was very high compared to Kenya 2006–2007 (Bird et al. 2008), confirming the huge scope of the outbreak in Madagascar.
The highest prevalences of RVF in livestock were detected where no clinical cases of RVF in livestock or humans were reported during the outbreak. This reflects the difficulties of case reporting from remote areas, especially with unknown emerging diseases, and highlights the need for extensive cross-sectional and longitudinal surveys to get a proper picture of the situation. Thanks to the greater involvement of the veterinary staff in the different regions of the country, the capacities for surveillance, early detection, and disease management were significantly improved during this outbreak. A surveillance system based on livestock sentinel herds was recently set up to monitor the circulation of the virus in the coming months. This will also contribute to a better understanding of the epidemiology of the disease in Madagascar and will help to identify options for its control and to confirm endemic ecozones.
Footnotes
Acknowledgments
We are grateful to the emergency rehabilitation coordination unit and Amadou Moustapha Kamara (Food and Agriculture Organization [FAO] of the United Nations, Antananarivo), Lanto Tiana Razafimanantsoa (Direction des Services Vétérinaires, Antananarivo), and Susanne Munstermann (Emergency Centre for Transboundary Animal Disease, FAO of the United Nations, Gaborone) for encouraging and facilitating this study. This study was supported in part by funds raised by World Health Organization and FAO through the Central Emergency Response Fund of the United Nations.
Disclosure Statement
No competing financial interests exist.