
Abstract
Triatoma infestans, one of the main vectors of Trypanosoma cruzi, is strongly associated with rural human ecotopes. Infested peridomestic places could act as a source of house infestation as a result of the movement of vectors among habitats. The quantitative study of phenotypic traits and nutritional variables could be informative about the population structure and the relative mobility of vectors. This is the first approach toward analyzing the phenetic characteristics of T. infestans and the relationship with their potential role to invade and colonize other habitats in the arid Chaco region. This article compares the phenotypic structure of T. infestans populations living in chicken coops and goat corrals, analyzing the geometric morphometry of wings and heads, antennal phenotypes, and the nutritional status of 257 adults and nymphs. The study area remained untreated with insecticides during the 15 years before the present study. The insects collected in peridomestic structures located in two zones 80 km apart exhibited significant differences in their morphotypes, suggesting an environmental effect and/or the absence of individual exchange between zones. The population structure was clear in T. infestans from goat corrals and chicken coops in one zone and less pronounced in the other. These results suggest the existence of a different rate of individual exchange among ecotopes within each zone. Morphometric variables of heads and wings were significantly correlated in insects collected in goat corrals but not in chicken coops, suggesting a habitat effect and supporting the hypothesis of different canalization forces that affect the two organs. The nutritional status of adults indicated a low dispersal probability with zonal differences. This article provides the first combined morphometric analysis of the head of adults and fifth instar nymphs in triatomines and the first combined analysis of head and wing in T. infestans.
Introduction
The region of Los Llanos, in the province of La Rioja, is one of the driest and poorest regions of Argentina. Goat breeding is the most important productive activity for small farmers (Porcasi et al. 2006). Goat corrals are the main refuge for the peridomestic populations of T. infestans and one of the ecotopes in which pyrethroid insecticides show low efficiency against these insects (Cecere et al. 1997, Gürtler et al. 2004). Chicken coops are also very frequent and maintain abundant vector populations. Infested peridomestic places could act as sources of house reinfestation after insecticide application (Gürtler et al. 2004), as a result of the movement of insects between habitats (Schofield 1985). Although T. cruzi infection in peridomestic T. infestans is not as frequent as in intradomestic insects, the peridomestic structures may be important sources of vector specimens.
Knowledge about a house colonization process is essential for vector control programs. In a few studied places, human houses remain free of T. infestans even in the proximity of infested goat corrals and chicken coops (Catalá et al. 1997, 2004, Gorla et al. 2010, Chagas Provincial Program of La Rioja, unpublished data). Studies showed that within an endemic region, each domicile could have a particular sociocultural pattern, economic level, and family structure, thus resulting in a particular way of life and a different risk to T. cruzi infection (Catalá et al. 2004). Other factors intrinsically related to T. infestans populations could also be involved in producing regional variables on the way the vectors invade the domiciles. The above-mentioned studies suggest that the movement of T. infestans between domestic or peridomestic structures could have spatial and temporal variations that might lead to regional differences. The quantitative study of phenotypic traits could be informative about the vectors' population structure and their relative mobility in space and time.
The high phenotypic plasticity of triatomines is well known, as shown by the high intraspecific morphological variability observed with different techniques (Dujardin et al. 1999, 2009, Jaramillo et al. 2002, Gumiel et al. 2003, Schachter-Broide et al. 2004, Vargas et al. 2006, Abrahan et al. 2008, Hernández et al. 2008). Morphometric characteristics of T. infestans populations are good indicators of the origin of insects re-colonizing human habitations (Dujardin et al. 1999, 2007). Similarly, it has been demonstrated that Rhodnius prolixus from palm trees were the cause of reinfestation of nearby domiciles in the state of Barinas, Venezuela (Feliciangeli et al. 2007). This conclusion, derived from the morphometric study of wings, was confirmed afterward by genetic studies on the same specimens (Fitzpatrick et al. 2008). In a similar fashion, Hernández et al. (2008) showed a strong association between macroenvironmental conditions, chromosomal cytotype, and antennal phenotypes of T. infestans, based on the study of populations collected in five of the seven countries in which the species is endemic. Abrahan et al. (2008) demonstrated heterogeneity in the phenotypes of spatially close T. infestans populations who are living in different habitats.
As part of a larger project aiming at analyzing the movement between intradomestic and peridomestic T. infestans populations in the arid Chaco region of Argentina, this article compares the phenotypic structure of T. infestans populations who are living in chicken coops and goat corrals. The nutritional status of adults was also examined as a proxy estimate of the dispersal rate. This article provides the first combined morphometric analysis of the head of adults and fifth instar nymphs in triatomines and the first combined analysis of head and wing in T. infestans.
Materials and Methods
Study area
The work was carried out in the Independencia department (7120 km2), located on the western side of the Los Llanos (La Rioja, Argentina). The area is located at the southern tip of the Gran Chaco (arid Chaco, Fig. 1). The main peridomestic structures found near the domicile are goat corrals and chicken coops. The insects were collected in these two types of peridomestic structures of rural houses in the localities of Patquía Viejo, Pozo de la Pampa, La Aguadita, Salinas de Bustos, and La Torre. The first two localities are 570 meters above sea level (masl), 80 km apart from Salinas de Bustos and La Torre, which are 1128 and 1182 masl, respectively. La Aguadita is located 60 km away from Patquía Viejo at an altitude of 900 masl (Table 1 and Fig. 1). The whole area remained untreated with insecticides during the 15 years before the present study. The localities were selected due to their abundant vector populations in the base line sampling (up to 43 insects per goat corral and up to 109 in chicken coops after 30 min of active search in each ecotope) and the support to the study provided by the house owners.

Map of the study area, showing the collection sites. The inset shows the location of the studied area (black box). Eastern zone (E): Patquía Viejo, Pozo de la Pampa, and La Aguadita; western zone (W): Salinas de Bustos and La Torre.
Number of T. infestans examined according to site of capture.
FChi, females in chicken coops; FGo, females in goat corrals; masl, meter above sea level; MChi, males in chicken coops; MGo, males in goat corrals.
The studied area shows similar house types and peridomestic buildings as well as cultural practices and strategies related to animal breeding. The movement of people or the exchange of animals occurs mainly between nearby localities, as the lack of transportation means and large distances between houses prevents movement between more distant localities.
In general, chicken coops are low rectangular structures, with a rudimentary roof. The most common building materials are wood on bricks, wire, leather, cardboard, and plastic (Fig. 2a). The goat corrals are generally made with tree branches horizontally intertwined and a covered area in which calves are kept (Fig. 2b).

Chicken coops (
The distance between chicken coops and goat corrals is variable (30–80 m). The average size of a goat herd was 100–130 animals per corral, including calves, at the time of the insect collection. Each sampled chicken coop had on average 8–10 chickens. The studied chicken coops did not include chicken nests, because they are unstable habitats that do not persist through time.
Environmental data
Environmental variables, especially temperature, are related with the distribution of T. infestans (Gorla 2002) and phenetic characteristics of individuals (Catalá et al. 2007).
Since no meteorological station exists within the study area (nearest one is 85 km away), Worldclim data at 30 arc seconds (
Insects
The insects were collected during September–October 2006, just before an official vector control intervention of the Provincial Program of Chagas (the first in 15 years). Twenty-one goat corrals and 12 chicken coops were manually examined by actively searching during 15 min (two persons) using an irritant dislodging agent (tetramethrin 0.2%). Thirteen goat corrals and 6 chicken coops were negative for adults and fifth instar nymphs. Table 1 summarizes the number and origin of insects. One hundred forty-three adults (62 females and 81 males) and 114 fifth instar nymphs (54 females and 60 males) were collected in chicken coops and goat corrals. All specimens were kept alive at low temperature (<10°C) until reaching the laboratory in which individual data were recorded and dissection was performed.
Nymph species were identified according to Brewer et al. (1981, 1983). The sex of fifth instar nymphs was recognized by examination of urosternites VIII and IX, as suggested by Brewer et al. (1981).
Reproductive and nutritional variables
The nutritional status of T. infestans adults was estimated as the body weight or body length ratio (W/L index), as suggested by Schofield (1980). The nutritional status was used as a proxy estimate of dispersal rate, as the percentage of individuals with W/L <8 mg/mm indicates high probability of flight initiation (Lehane and Schofield 1982, Lehane et al. 1992). In females with eggs, the number of chorionated eggs present in the oviducts was examined to improve the estimation of the actual nutritional status (blood stored in the promesenteron), as each egg weighs (2.3 mg), which can seriously distort the index W/L and does not allow comparison with males.
The insects were weighed on an analytical balance with an accuracy of 0.1 mg and measured from clypeus to abdominal tip using a caliper.
Statistical analysis was performed using univariate methods to make comparisons between populations with T. infestans collected from different habitats and populations of the same sex who were collected from the same habitat. Analysis of variance or Kruskal–Wallis test was used, after testing for the assumptions of parametric statistics using Statistica (StatSoft, Inc., 2005).
Geometric morphometrics of the head and wings
Each collected insect was dissected and preserved in 70% alcohol for later analysis. Heads were cut off by the collar and mounted on a pin attached to a wire rack. The right wings were placed between slide and cover objects, covered with 70% alcohol, and photographed using a camera stabilizing support. The photographs were taken with a Kodak C613 digital camera (6.2 MP) and with the aid of a stereomicroscope in the case of the heads. Ten landmarks type I were selected on the right wing of adults and 10 type II on the ventral surface of head of adults and nymphs V. For heads, the average of the landmarks on both sides (five landmarks) was used to reduce intraindividual variation and minimize digitization errors (Fig. 3).

Points of reference (landmarks) measured as coordinates of heads (
Shape variables were obtained through the generalized procrustes analysis superimposition algorithm and the subsequent projection of the procrustes residuals into a Euclidean space (Rohlf 1999). Both nonuniform (“partial warps,” strictly speaking) and uniform components (Rohlf 1990) were used as shape variables. The uniform component describes global variation such as stretching and compression, and the nonuniform component corresponds to local variation (Zelditch et al. 2004). These two components describe the differences in shape as deviations from an average configuration of landmarks.
When possible, the problem of sample size lower than the number of shape variables was circumvented by using a restricted representation of shape, that is, a set of principal components (relative warps) derived from the shape variables. These relative warps were used as input into a discriminant analysis. The significance of the discriminant analysis was evaluated by the values of Wilks, and Mahalanobis distance, and these values were tested using a nonparametric test based on permutations (1000 runs). Bonferroni correction was used for multivariate comparisons.
For comparing overall wing or head size among groups, the isometric estimator known as “centroid size” (CS), derived from coordinates data, was used: It is defined as the square root of the sum of the squared distances between the center of the configuration of landmarks and each individual landmark (Bookstein 1991).
The variables of size and shape of wings and heads were used for comparisons of habitat (chicken coops vs. goat corrals), sex, and zone. The statistical significance of CS differences was assessed by using a nonparametric test based on permutations (1000 runs). Heads were compared for both adults and fifth instar nymphs at the same landmarks.
The relationship between CS and shape discrimination among groups (allometry) was estimated by the regression on size variation of the discriminant axes derived from shape.
The two-block partial least squares analysis was used to assess the covariation between the shapes of wings and heads.
The linear correlation between the sizes of wings and heads was estimated with the Pearson correlation coefficient (r).
Software used in the morphometric analysis
The COO (version 39) enabled the digitization of landmarks, VAR (version 1.4) was used for nonparametric ANOVA of size, COV (version 90) for the two-block partial least squares analysis, MOG (version 82) for performing the Procrustes superimposition and generating partial warps and CS for each individual, and PAD (version 84) for discriminant analysis and nonparametric estimation of the statistical significance of each Mahalanobis distance. The modules COO, MOG, COV, PAD, and VAR were developed by J.P.D. and are available at no cost at
The Statistica package (StatSoft, Inc., 2005) was used for other univariate comparisons (parametric ANOVA and Kruskal–Wallis).
Analysis of the antennal phenotype
The antennae of adults were extracted and processed with sodium hydroxide (4%) and neutralized with glacial acetic acid (5%). This procedure allowed cuticle diaphanization and made possible the identification and counting of sensilla using a stereomicroscope at 400 × equipped with a draw chamber. The antennae were mounted “in toto” on slides, using glycerin. For each specimen, the ventral side of the three distal segments of the antennae (pedicel, flagellum 1, and flagellum 2) was drawn, identifying and counting the sensilla according to Catalá and Schofield (1994): bristles (BR), thin-walled trichoid (TH), thick-walled trichoid (TK, except in the pedicel), and basiconica (BA, except in the pedicel). The number of sensilla per segment (antennal phenotype) allowed for comparisons across zones and between habitats in both sexes.
Antennal variables were compared using analysis of variance when data met the assumptions of parametric statistics. Alternatively, nonparametric equivalent comparisons were used (Kruskal–Wallis). The analysis included a multivariate discriminant analysis using sensilla that are not related to sexual differentiation (seven variables) in the comparison of zones. For the comparisons of the two habitats within each zone, the sensilla that showed significant differences in univariate analysis were used. The significance of the discriminant analysis was evaluated by the values of Wilks, and Mahalanobis distances, using a nonparametric test based on permutations (1000 runs). Bonferroni correction was used for multivariate comparisons (PAD software).
Results
Characterization of the study area based on environmental variables
The cluster analysis using either the 15 variables of Table 2 or the 9 temperature-related variables grouped the five studied locations into two zones, called East (E), which included Patquía Viejo, Pozo de la Pampa, and La Aguadita, and West (W), which included Salinas de Bustos and La Torre (Fig. 4).
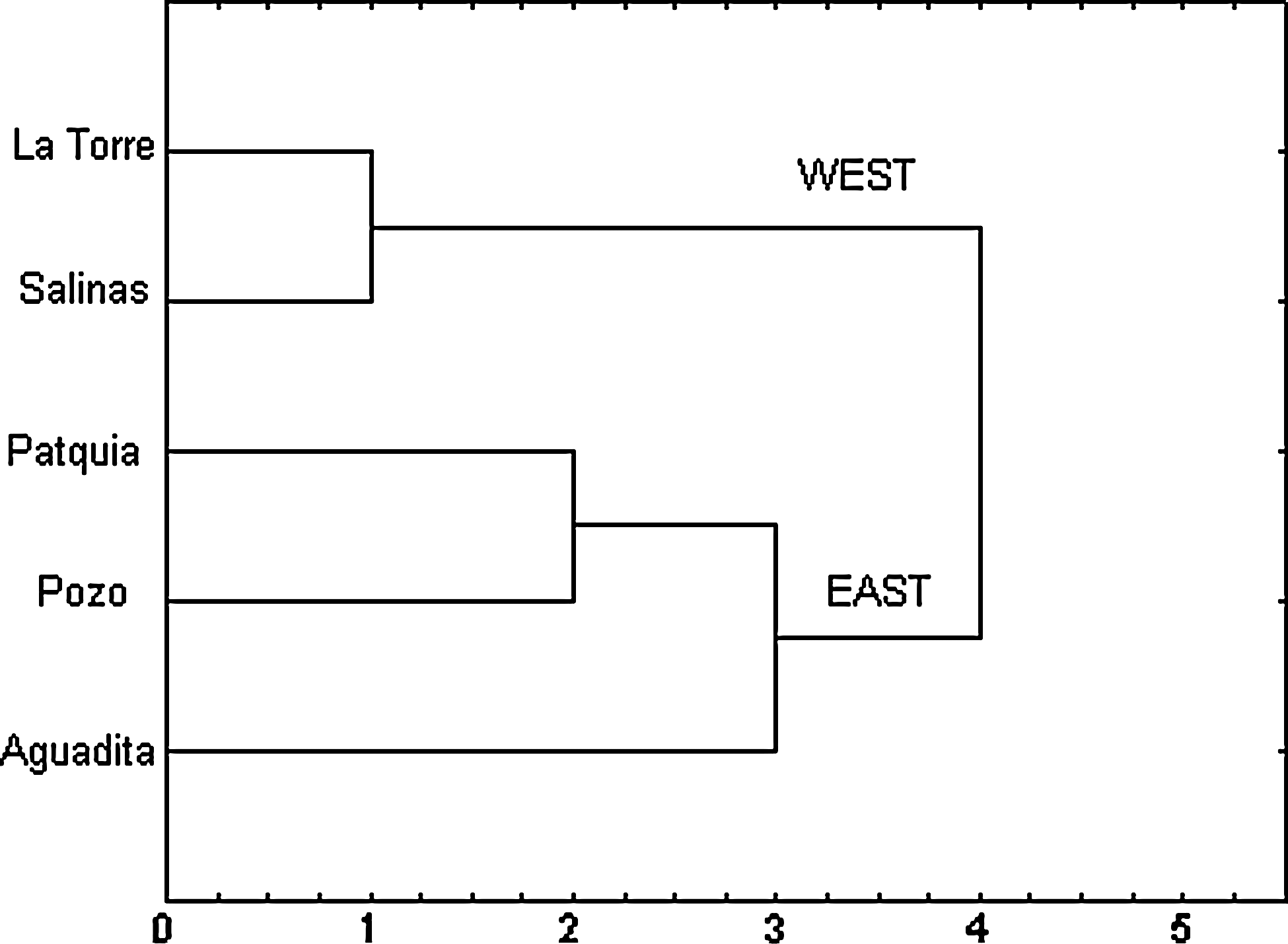
UPGMA dendrogram of localities, grouped by the similarity of temperature variables extracted from the Worldclim database.
Differentiation of two geographical zones using geometric morphometrics and antennal phenotypes
The first approach was to analyze all the insects collected in the study area as a whole (five localities), mainly because the number of insects collected in the habitat samples was lower than the number of specimens required for the morphometric analysis. However, a preliminary morphometric analysis of wings and heads (Abrahan and Hernández, unpublished data) showed more similarity between the individuals from Patquia Viejo, Pozo de la Pampa, and La Aguadita than those from La Torre and Salinas de Bustos. Since the sample size did not allow a morphometric analysis by locality, these were grouped in the two zones identified by the analysis of the environmental variables. In the later paragraphs, analyses are discussed first by zone, grouping the insects captured in both habitats, and then by habitat within each zone.
Comparison of morphotypes between zones
Both heads (adults and nymphs) and wings (adults) were larger in the western zone (Table 3). Shape variables differentiated the nymphs (both sexes) and adult males from both zones but not the females (Table 3).
Antennal phenotypes (number of significant variables) are shown in parenthesis.
A, adults; CS, centroid size; M, male; F, females; Multiv., multivariate analysis; N, nymphs; NA, not applicable; NS, not statistically significant; S, statistically significant; SH, shape; Univ., univariate analysis; W/L, body weight/body length ratio.
p < 0.01.
p < 0.05.
Antennal phenotypes
The univariate analysis of antennal phenotypes showed significant differences (p < 0.05) between zones for both sexes. In females, the TH of the pedicel showed a higher average in the eastern zone, whereas the TH of flagellum 1 and the BA of flagellum 2 showed a higher average in the western zone. In males, the number of TH and BA of flagellum 1 showed higher values in the western zone. Sexual dimorphism was observed in both zones, mainly in the number of TH of pedicel, more abundant in males.
Discriminant analysis for both sexes using six variables (TH and BA from flagellum 1 and BR, TH, TK, and BA from flagellum 2) showed a significant separation of insects from the two zones (p < 0.0006).
Habitat morphotypes within each zone
Eastern zone
Size: The head size (CS) of nymphs (both sexes) and adult males was significantly higher (p < 0.01) in insects collected in chicken coops than in those collected in goat corral (Table 4). Adult females showed no statistically significant differences. The same effect was found in wing size of adults (p < 0.01). Sexual dimorphism of the size of heads and wings in adults from chicken coops was found (p < 0.001) but not in adults from goat corrals and in the heads of nymphs of both habitats.
Differences statistically significant between habitats (p < 0.01) for comparisons within the same sex and zone.
M, males; F, females; Q25, quartiles 25; Q75, quartiles 75.
Shape: The nymphs of both sexes collected in chicken coops and goat corrals differed in the head shape (p < 0.05; five components, 96% of total shape variance).
In adults, differences in head shape were found only in males (p < 0.05; 2 components, 95% of total shape variance) (Table 3).
The wing shape showed no statistically significant differences either between habitats or between sexes (six components, 83% of total shape variance) (Table 3).
Allometry: The contribution of size to head shape differentiation was 9% (nymphs) and 0% (adults). This allometric effect was 0% for the wings.
Heads and wings correlation: A positive linear correlation between head size and wing size was found among the specimens collected in the goat corrals but not in those collected in the chicken coops (Fig. 5). The wing size showed higher variability at intermediate values of head size in both sexes. There was no statistically significant covariation between shape of head and shape of wing, irrespective of the sex or the habitat.
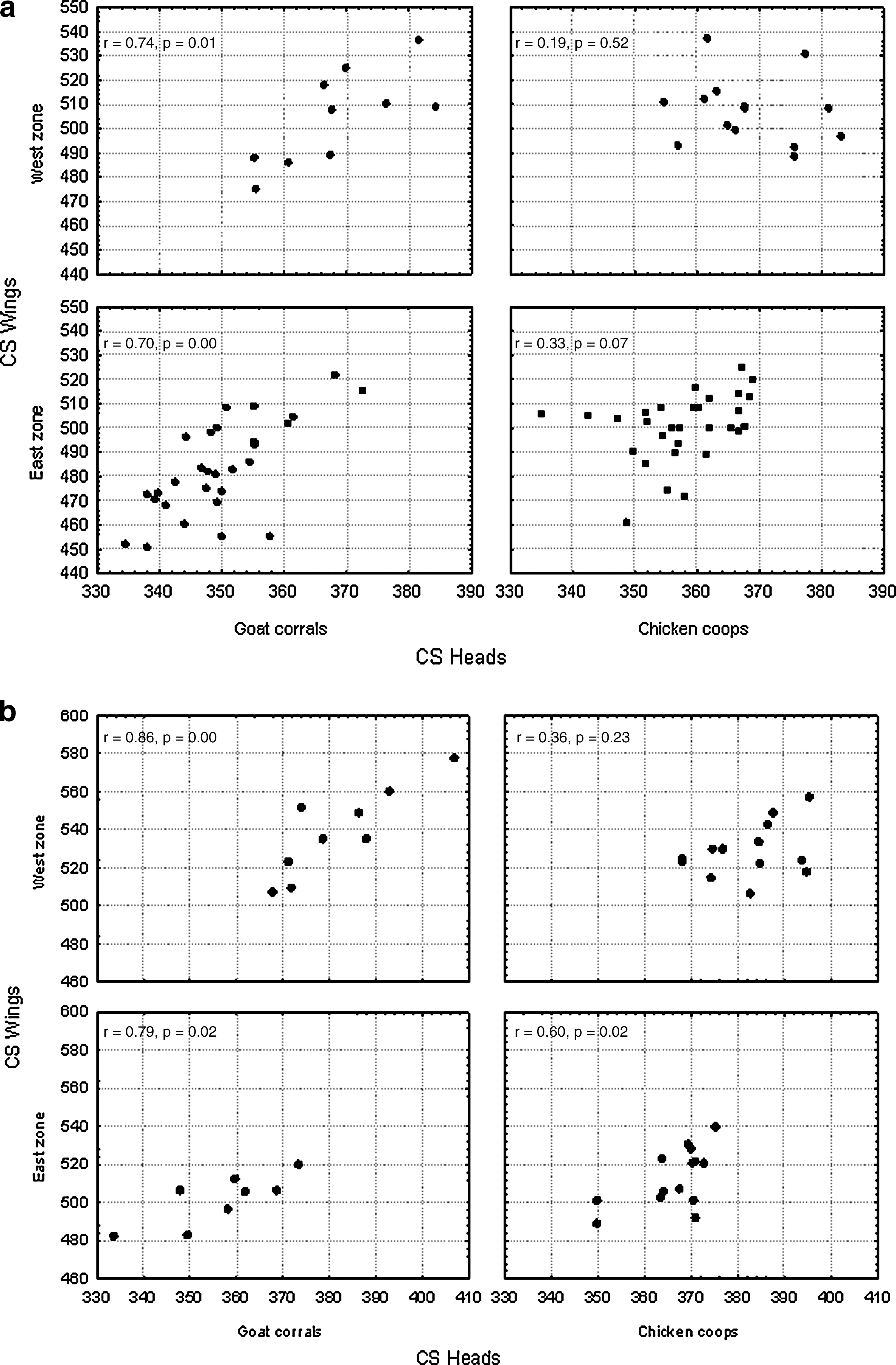
Regression of head and wing size (CS) for the two habitats. (
Antennal phenotypes: Variation in the number of TH of flagellum 2 was observed in both sexes, with higher average values in insects from chicken coops (33.65) than in insects from goat corrals (30.59), (p = 0.004). The discriminant analysis for both sexes showed significant discrimination between specimens of the two habitats (p = 0.0064) using five variables of the flagellum (F1TH, F1TK, F2TH, F2TK, and F2BA).
Nutritional status of adults: The nutritional status estimated as W/L ratio was not different among habitats (females: median = 15.36 and 15.20 mg/mm; males: 11.07 and 11.39 mg/mm, respectively, for goat corrals and chicken coops) (Table 3). Male specimens collected from goat corrals had minimum values of W/L between 5 and 6 mg/mm, whereas those collected from chicken coops had the minimum value of 9 mg/mm. In addition, in males from goat corrals, a bimodality in the frequency distribution was observed (Shapiro-Wilks test: w = 0.92, p = 0.02), indicating the existence of two groups with different nutritional status (data not shown). In females, the minimum values of W/L were >7 mg/mm in chicken coops and >6 mg/mm in goat corrals. In goat corrals, 14% of females and 17% of males showed values less than 8 mg/mm. In chicken coops, there were no insects with W/L below this value.
Western zone
Size: The average values of head CS were higher in insects collected in chicken coops than in those collected in goat corrals (nymphs and adults). This difference was significant only in male nymphs (Tables 3 and 4). Sexual dimorphism was significant in adults of chicken coops and in the two habitats for nymphs.
Wing size was not different between individuals of different habitats. Sexual dimorphism was significant (p < 0.004) in the two habitats.
Shape: The nymphs, either males or females, showed no differences between habitats with regard to the head shape (five components, 94% of shape variance). In adults, females from chicken coops and goat corrals differed in head shape, after removing the size influence (p < 0.05, 5 components, 96% of shape variance).
The wing shape showed no statistically significant differences between habitats for either sex (six components, 86% conformation).
Allometry: The contribution of size to head shape differentiation was 18%, whereas in wings it was 4%.
Heads and wings correlation: A positive relationship between head size and wing size was found in insects of goat corrals but not in those of chicken coops (Fig. 5). This feature was observed in both males and females. There was no statistically significant covariation between wing shape and head shape; this was verified in each sex and in each habitat.
Antennal phenotypes: High similarity was found in the antennal phenotypes of adults in both habitats. In females, the mean value of F1TH was higher in insects of goat corrals (88.85) than in those of chicken coops (77.54, p = 0.01). In males, F2TH were more abundant in insects of chicken coops (35.05, p < 0.001) than in those of goat corrals (30.58). Multivariate analysis confirmed the absence of significant differences between the two habitats (Table 3).
Nutritional status of adults: Nutritional status estimated as W/L ratio was higher in T. infestans males collected in goat corrals than in the males collected in chicken coops. In females, the difference was not statistically significant (female: median = 15.58 and 13.45 mg/mm, males: 12.95 and 10.21 mg/mm, respectively, for goat corrals and chicken coops) (Table 3). The frequency distribution showed 0% of individuals with W/L below 8 mg/mm (both sexes in the two habitats).
Discussion
Chicken coops and goat corrals are the main peridomestic habitats for T. infestans in the region of Los Llanos (La Rioja) (Porcasi et al. 2007, Gorla et al. 2010, Chagas Provincial Program of La Rioja, unpublished data). The persistence of the peridomestic populations of the vector is a serious problem due to the frequently high densities of their populations and proximity to human habitations. This is the first study of the relationship between the phenetic characteristics of T. infestans living in these habitats and their potential role to invade and colonize other habitats in the arid Chaco region.
Differences in morphotypes among close geographical zones
In this study, two zones 80 km apart exhibited differences in the morphotypes of T. infestans. The wing, antennal, and head phenotypes of the eastern and western zones had different characteristics, suggesting environmental effects, low or no exchange of individuals between zones, or both.
Nymphs and adults of the western zone had larger body size than those of the eastern zone. Insect body size is strongly influenced by environmental conditions. The difference in body size of the eastern and western specimens was expected, due to the difference in altitude between zones (and therefore in temperature) (Vanvoorhies 1996). It is well known that the negative relationship between environmental temperature and body size of an insect responds to a complex pattern that involves individual phenotypic variation and intraspecific genetic diversity in geographical terms (David et al. 1994, Vanvoorhies 1996, Dujardin et al. 2009).
There are evidences that body sizes of T. infestans specimens from the Bolivian Andes are larger than those of the Gran Chaco region, a difference that has been associated with environmental characteristics, especially temperature variation (Bergman's rule) related to altitude (Catalá et al. 2007) and with genetic drift and founder effects (Dujardin et al. 1998). This rule also applies here when comparing T. infestans from localities over 1000 masl with those below that altitude. The size of insects from La Aguadita (900 masl) was similar to the size of those from localities at 500–600 masl and differed significantly from those in localities over 1000 masl, suggesting that a genetic drift effect on the insects size could not be excluded.
The results of this study show that the shape of wings and heads differ between the two studied zones. Since shape variables of T. infestans are frequently driven by genetic variations (Dujardin et al. 2009), shape differences between eastern and western zones would suggest the existence of genetic isolation between them. The spatial structure of T. infestans populations at the macrogeographic level is similar to the finding in other geographical zones of Argentina, at a similar geographical scale, (Pérez de Rosas et al. 2007) and Perú (Pizarro et al. 2008), using molecular techniques. Low or no exchange of individuals between relatively close areas is a positive situation for vector control interventions, because this spatial structure makes less likely that an insecticide-treated zone would be recolonized from eventually nontreated nearby areas, thus promoting higher spraying efficiency.
Population structure and habitats
Within each zone, T. infestans showed different degrees of population structure in goat corrals and chicken coops. In the western zone, morphometric variables were spatially homogeneous, indicating low population structuring and suggesting a relatively high individual exchange between habitats. On the other hand, adults showed good nutritional status in the two western habitats (0% of individuals with very low nutritional status, W/L <8 mg/mm), indicating a low probability of dispersal (Lehane and Schofield 1982, Lehane et al. 1992, Gurevitz et al. 2006). It would be expected that low population structuring would be associated with low nutritional status, as starved individuals would disperse to locate eventual blood sources. However, individuals in the western zone show good nutritional status associated with low spatial structure indicated by the morphometric homogeneity, and individuals in the eastern zone show low nutritional status associated with high spatial structure. The explanation to this apparent contradiction could lie in the temporal window reflected by each measurement type. The nutritional status is an instantaneous indication of the physiological status of an individual (at sampling time), whereas the morphometric variables reflect past conditions that prevailed during the preadult development. In the scenario just described, the low nutritional status of some adults in the eastern zone would strongly suggest a future short-term movement of individuals and changes in the population structure. In contrast, the high nutritional status of the western individuals associated with low population structure suggests a recent dispersal event that allowed the insects the procuration of a better nutritional status.
Food availability is one of the main modifiers of body size in insects (Jirakanjanakit et al. 2007, Caro Riaño et al. 2009). Most of the T. infestans specimens from both zones collected in chicken coops had bigger CS of wings and heads than the specimens collected in goat corrals. This would indicate a difference in the feeding behavior or in the nutritional value of the host blood. The blood of mammals and birds differs in quality for triatomines (Guarneri et al. 1998, 2000). According to López et al. (1999) and Ceballos et al. (2005), the chicken coops seem to be the habitats in which T. infestans achieves the best nutritional status. Nutritional status data showed no starved individuals (with W/L <8 mg/mm) in chicken coops. In the eastern zone, 31% of adult insects collected in goat corrals were starving. These results suggest that a high number of individuals would disperse from the goat corrals. A particular consideration should be made with regard to chicken incubation sites (not included here) that are abandoned after the incubation period, usually leaving a large population of T. infestans without food access.
The integrated analysis of the size and shape of heads and wings demonstrates the lack of correlation between head and wing CS in the insects from chicken coops, contrasting with the opposite relationship in the insects from goat corrals, thus indicating a habitat effect. The correlation could not be explained by a higher size heterogeneity among corrals, because no such pattern was apparent. These results could suggest a trade-off between head and wing development during the insect growth. The head is an essential organ for each stage of the insect life. It is then supposed to receive priority in achieving a harmonious development, compared with the wings, whose mission is limited to one developmental stage only (to disperse adults). It is possible that in case of food scarcity, the wings would receive relatively less resource for their development and maintain a relatively smaller size. The lack of shape covariation between heads and wings supported the hypothesis of different canalization forces for the two organs (Caro Riaño et al. 2009). No statistically significant association between body length and size of wings or heads was detected.
This article provides the first combined morphometric analysis of the head of adults and fifth instar nymphs of T. infestans. In our hypothesis, in the absence of individual exchange between habitats, the population structure (interpreted as different morphotypes) should be observed in nymphs and adults. In the eastern zone, both nymphs and adult males showed significant differences between habitats, a difference that was not observed in adult females. This finding would confirm a higher mobility of adult females and a relatively more important structuring in male populations. The similarity of morphotypes found in chicken coops and goat corrals in the west, in both nymphs and adults, suggests the existence of higher flow of individuals between habitats in the zone.
Recent studies showed that T. infestans is a species with weak morphological canalization (Dujardin et al. 2009), which is also supported by the results of this work. The studied populations have a different regional structure in the habitats they occupy. Temporal variation of this structure and the dispersal pattern of the species are aspects that deserve additional studies. A better understanding of the process driving the movement of individuals of this species and its variations over time would help in the improvement of vector control activities in the Gran Chaco region of South America.
Footnotes
Acknowledgments
To the Provincial Chagas Program of La Rioja and Elmer Rodríguez that contributed in the field work. To Natalia Folguera and Ivana Amelotti for their support during the laboratory work. This study received financial support from FONCYT PICT 2006 878 and CONICET (Argentina). M.L.H., D.E.G., and S.S.C. are supported by the Consejo Nacional de Investigaciones Científicas y Técnicas (Argentina).
Disclosure Statement
No competing financial interests exist.