
Abstract
Infection by Anaplasma and Ehrlichia in feral raccoons (Procyon lotor) in Hokkaido, Japan, was examined by molecular methods. A polymerase chain reaction (PCR) screen for Anaplasmataceae, based on 16S rRNA, showed that 38 (5.4%) of 699 raccoons examined were positive. These 38 positive samples were examined for Anaplasma phagocytophilum, Anaplasma bovis, Ehrlichia chaffeensis, and Ehrlichia canis infection by species-specific nested PCR. Nested PCR results indicated that 36 of the 38 samples were positive for A. bovis. All 38 samples were PCR negative for A. phagocytophilum, E. chaffeensis, and E. canis. This is the first report of the detection of A. bovis in the peripheral blood of raccoons. A total of 124 raccoons were infested with ticks, including Ixodes ovatus, Ixodes persulcatus, and Haemaphysalis spp. The rate of A. bovis infection in raccoons infested with Haemaphysalis spp. (46.7%, 7/15) was significantly higher than that in raccoons without Haemaphysalis spp. infestation (3.7%, 4/109, p < 0.001). No significant differences were observed in A. bovis infection rates between raccoons infested with I. ovatus or I. persulcatus and those not so infested. A total of four ticks (two males and two nymphs) and one larval pools from four raccoons showed positive for A. bovis–specific nested PCR. This results support the correlation between the A. bovis infection of raccoons and Haemaphysalis infestation. In conclusion, raccoons could be possible reservoir animals for A. bovis, and A. bovis infection in raccoons may be related to infestation with Haemaphysalis spp.
Introduction
Anaplasma and Ehrlichia, both originally found in the United States (McQuiston et al. 1999, Rikihisa 1991), might have been introduced into Japan by raccoons. DNA fragments of A. phagocytophilum and Anaplasma bovis have recently been detected from Sika deer (Cervus nippon), cattle, and several tick species in Japan (Ohashi et al. 2005, Kawahara et al. 2006, Ooshiro et al. 2008, Jilintai et al. 2009, Wuritu et al. 2009, Yoshimoto et al. 2010). E. chaffeensis DNA has also been detected from Sika deer in Japan (Kawahara et al. 2009). We therefore attempted to detect A. phagocytophilum, A. bovis, E. chaffeensis, and E. canis pathogens of both human and veterinary importance, by molecular analysis of peripheral blood samples obtained from feral raccoons in Japan. The objective of this study was also to clarify the epidemiologic role of raccoons for these pathogens in Japan.
Materials and Methods
Raccoon blood and tick samples
A total of 699 raccoons were captured between May and October 2007 and between March and October 2008, as part of raccoon population control programs implemented by the Hokkaido Government and the Ministry of the Environment in west-central Hokkaido, Japan (Fig. 1). Raccoons were anesthetized with an intramuscular injection of ketamine hydrochloride. Blood samples from the heart were collected in ethylenediaminetetraacetic acid tubes and centrifuged at 1000 g for 10 min; plasma was removed and the remaining blood components were frozen at −20°C for polymerase chain reaction (PCR) assays.

Shading shows the location of the study area.
Raccoons were also examined for tick infestation. When tick infestation was observed, as many ticks were collected as possible and stored in 70% ethanol for later morphological identification.
DNA extraction and PCR
DNA was extracted from blood or tick samples using a QIAamp DNA Mini Kit (Qiagen GmbH, Hilden, Germany). DNA samples were stored at −20°C in 200 μL of Tris-EDTA buffer until further use.
Screening PCR was performed using the group-specific primer pair, EHR16SD and EHR16SR, which amplifies the 16S rRNA gene of the family Anaplasmataceae (Parola et al. 2000).
Samples that are positive for the screening PCR were examined by species-specific nested PCR. We used the primer pair EC9 and EC12A for the first amplification of A. phagocytophilum and A. bovis DNA. The resulting PCR products were then used as templates for the species-specific second amplification (Kawahara et al. 2006) with primers AB1f and AB1r for A. bovis (Kawahara et al. 2006), and newly designed primers AP-f1 and AP-r1 for A. phagocytophilum. The second PCR for A. phagocytophilum was performed under the following conditions: 40 cycles of denaturation (95°C, 60 s), annealing (58°C, 60 s), and extension (72°C, 60 s). For E. chaffeensis, primers NS16SCH1F and NS16SCH1R were used for the first amplification and NS16SCH2F and NS16SCH2R for the second (Kawahara et al. 2009). For E. canis, primers fD1 and EHR16SR were used for the first PCR, and E. canis–specific primers CANIS and GA1UR were used for the second PCR (Warner and Dawson 1996). To prevent contamination in nested PCR, we have performed each reaction setup in separate chamber using exclusive pipets and tips. Nucleotide sequences of all primers used here are listed in Table 1. Specificity and sensitivity of newly designed A.phagocytophilum–specific nested PCR were examined by using DNA of A. phagocytophilum, A. bovis, Anaplasma marginale, Anaplasma central, Anaplasma platys, E. canis, Ehrlichia muris, Wolbachia pipientis, Neorickettsia risticii, and R. rickettsia DNA. The sensitivity of the nested PCR was also examined by using diluted DNA extracted from A. phagocytophilum infected culture cells.
PCR, polymerase chain reaction.
In the nested PCR, DNA samples extracted from the A. phagocytophilum strain Webster, E. chaffeensis strain Arkansas, and E. canis strain Israel were used as positive controls, and distilled water was used as the negative control. No positive controls were used for the A. bovis–specific PCR to prevent cross contamination.
Sequencing and phylogenetic analysis
To determine nearly full-length sequences of the 16S rRNA genes amplified by species-specific nested PCR, additional PCR amplifications were performed using primer sets fD1 and EHR16SR, and EHR16SD and Rp2, respectively (Weisburg et al. 1991). When a strong band was detected after PCR, products were purified using the Qiaquick PCR purification kit (QIAGEN GmbH). Direct sequencing of the PCR products and analysis of sequences obtained were performed as described previously (Inokuma et al. 2007b, 2008). Homology searches based on the sequences of the PCR products were performed using BLAST (National Center for Biotechnology Information). Phylogenetic relationships of the obtained sequences to other sequences registered in GenBank were determined using the neighbor-joining method.
Statistical analysis
Chi-squared tests were performed to compare rates of species-specific nested PCR amplification in raccoons infested with each tick species against those from noninfested raccoons. StatMate IV Version 4.01 was used for the analysis, where a p-value of <0.05 was considered significant.
Results
In the initial screening PCR, 38 (5.4%) of 699 raccoons examined were positive for Anaplasmataceae. Species-specific nested PCRs on these 38 positive samples showed that 36 were positive for A. bovis, whereas all were PCR negative for A. phagocytophilum, E. chaffeensis, and E. canis. Randomly selected 7 positive amplicons among 36 positives for A. bovis–specific nested PCR were analyzed for nucleotide sequences to confirm the results. All seven sequences were identical and showed 99.8% nucleotide identity (510/511 bp) to the 16s rRNA gene of A. bovis detected from South Africa (U03775). To confirm the result of rest of 2 among 38 positive in the initial screening PCR, the 2 amplicons of the screening PCR were analyzed by direct sequence method; however, they cannot be determined.
In the specificity test for the newly designed A. phagocytophilum–specific nested PCR, only A. phagocytophilum DNA was positive. The sensitivity test of the nested PCR revealed that it can detect DNA extracted from one infected cell in 1 μL (data not shown).
A total of 13 samples were successfully sequenced approximately 1400 bp of the 16S rRNA gene, excluding the primer region. These sequences have been deposited in GenBank under the accession numbers GU937011 to GU937023. The 13 sequences were identical and showed 99.7% nucleotide identity to the 16s rRNA gene of A. bovis detected from South Africa (U03775). This sequence was also 99.5% identical to an A. bovis 16s rRNA sequence detected from Sika deer in Shimane, Japan (AB211163). In the 16S rRNA gene-based phylogenetic tree, these 13 sequences clustered in the same clade as A. bovis (Fig. 2).
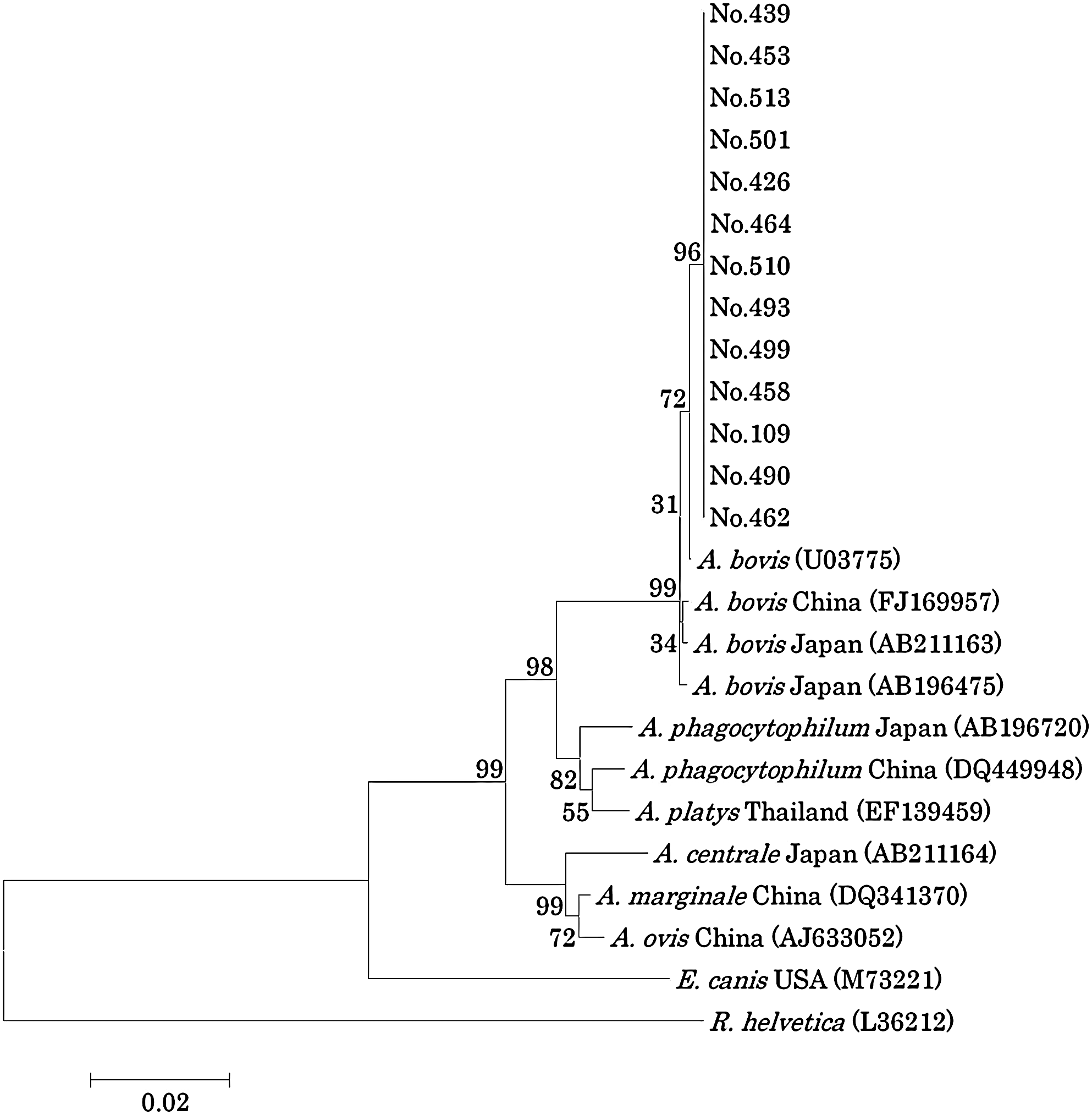
Phylogenetic relationships among various Anaplasma spp. based on the nucleotide sequences of the 16S rRNA gene. Clustal W software and the neighbor-joining method were used to construct the phylogenetic tree. Scale bar represents 2% divergence. Numbers at the nodes are percentages of bootstrap re-samplings supporting the topology shown. Numbers 109, 426, 439, 453, 458, 462, 464, 490, 493, 499, 501, 510, and 513 are identification numbers of raccoons from which DNA was isolated.
A total of 672 ticks were collected from 124 of the 699 raccoons. Ixodes ovatus was the predominant tick species, followed by Haemaphysalis spp. and Ixodes persulcatus (Table 2). It was difficult to identify the Haemaphysalis specimens at the species level because most of ticks were fully engorged or semi-engorged. The rate of A. bovis infection in raccoons infested with Haemaphysalis spp. (46.7%, 7/15) was significantly higher than that in raccoons without Haemaphysalis spp. infestation (3.7%, 4/109, p < 0.001). No significant differences were observed in A. bovis infection rates between raccoons infested with I. ovatus or I. persulcatus and those not so infested (Table 3).
Number of raccoons species-specific nested PCR positive for Anaplasma bovis.
p < 0.001.
Number of raccoons species-specific nested PCR negative for A. bovis.
A total of 57 Haemaphysalis spp. (4 males, 18 nymphs, and 35 larvae) were collected from 7 raccoons among 36 positives for A. bovis–specific nested PCR. DNA was extracted from each adult and nymphal tick. Five larvae were pooled into one tube and DNA was also extracted from each tube. A total of 29 tick samples (4 males, 18 nymphs, and 7 larval pools) were analyzed to detect A. bovis by the species-specific nested PCR. As a result, 5 among 29 samples from 4 raccoons were positive (2/4 males, 2/18 nymphs, and 1/7 pools of larvae) (Table 4).
N, nymph; M, male; L, larval pool.
Discussion
In a screening PCR based on 16S rRNA for Anaplasmataceae, 38 of 699 raccoons tested were positive. These 38 positive samples were examined for A. phagocytophilum, A. bovis, E. chaffeensis, and E. canis infection by species-specific nested PCR. These samples included 36 that were positive for A. bovis and two that were negative in all the nested PCRs. It is possible that those two raccoons were infected with other species in the Anaplasmataceae family. This is the first report of the detection of A. bovis from the peripheral blood of raccoons; these results suggest that raccoons could be possible reservoir animals for A. bovis.
Although the known vectors of A. bovis are Hyalomma spp., Rhipicephalus appendiculatus, and Ammblyomma variegatum (Donatien and Lestoquard 1936, Matson 1967, Rioche 1967), those particular ticks have not been detected in Japan. A. bovis DNA has been detected from Haemaphysalis longicornis and Haemaphysalis megaspinosa collected from Honshu and Hokkaido, respectively (Kawahara et al. 2006, Yoshimoto et al. 2010), and these ticks are possible vectors of A. bovis in Japan. In our study, A. bovis infection rates in raccoons infested with Haemaphysalis spp. were significantly higher than those in raccoons without Haemaphysalis spp. infestation. No significant differences in A. bovis infection rates were observed between raccoons infested with I. ovatus or I. persulcatus and those not infested. A. bovis infection in raccoons may be related to infestation with Haemaphysalis spp. To confirm the correlation between the positive raccoons for A. bovis–specific nested PCR and the infestation with Haemaphysalis spp., Haemaphysalis ticks collected from seven raccoons that showed positive for A. bovis–specific nested PCR. As a result, four ticks (two males and two nymphs) and one larval pools from four raccoons showed positive for A. bovis–specific nested PCR. This results support the correlation between the A. bovis infection of raccoons and Haemaphysalis infestation. Negative ticks might not intake enough amount of blood to be infected from the host animals. Further studies are required to confirm the role of Haemaphysalis spp. in A. bovis infection.
A. bovis causes an economically devastating disease in livestock; its principal symptoms include fever, anorexia, diarrhea, and, infrequently, involvement of the central nervous system (Matson 1967). Because raccoons frequently come near areas where humans and domestic animals live, it is possible that A. bovis infection is spread widely among domestic animals by raccoons.
All 38 samples subjected to nested PCR were negative for A. phagocytophilum, E. chaffeensis, and E. canis. A. phagocytophilum causes granulocytic anaplasmosis in humans, dogs, and horses, and pasture fever in ruminants (Chen et al. 1994, Rikihisa 2006, Inokuma 2007). Recently, A. phagocytophilum DNA has been detected from I. persulcatus, I. ovatus, Sika deer, and cattle in Japan, including Hokkaido (Ohashi et al. 2005, Kawahara et al. 2006, Ooshiro et al. 2008, Jilintai et al. 2009, Wuritu et al. 2009, Yoshimoto et al. 2010). E. chaffeensis is an agent that causes monocytotropic ehrlichiosis in humans and dogs (Paddock and Childs 2003, Inokuma 2007), and its DNA was recently detected from Sika deer in Japan (Kawahara et al. 2009). Although raccoons are some of the important reservoir animals of A. phagocytophilum and E. chaffeensis in the United States (Comer et al. 2000, Levin et al. 2002, Dugan et al. 2005, Yabsley et al. 2008), neither of these pathogens were detected from raccoons in this study.
E. canis is an important pathogen that causes canine ehrlichiosis (Inokuma et al. 2003). Although there have been reports of raccoons infected with E. canis and seropositive for E. canis in the United States and Japan, respectively (Dugan et al. 2005, Inokuma et al. 2007a), E. canis DNA was not detected from raccoons in this study. It is possible that E. canis was not introduced into Japan by raccoons.
This study suggests that raccoons could be possible reservoir animals for A. bovis and that they play an important role in the maintenance of A. bovis in nature. More epidemiologic studies are required to confirm the epidemiologic role of raccoons in A. bovis infection.
Footnotes
Acknowledgments
We wish to thank the Nature Preservation Division of Hokkaido Government, Ministry of the Environment and Hokkaido Forest Management Corporation for providing opportunities to collect samples. We are also grateful to Dr. Tsubota, Laboratory of Wildlife Biology (Graduate School of Veterinary Medicine, Hokkaido University), for laboratory support. This study was supported in part by Grant H21-Shinkou-Ippan-014 for Research on Emerging and Re-Emerging Infection, and Grants-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (No. 21380192).
Disclosure Statement
No competing financial interests exist.