
Abstract
The hard tick Ixodes ricinus is the principal vector of tick-borne encephalitis virus (TBEV) in Slovenia; but until now, there was no information about the prevalence of TBEV infection in Slovenian ticks. We conducted a 2-year survey in 2005 and 2006, during which we were collecting I. ricinus ticks monthly in eight different locations of Slovenia. A total of 4777 I. ricinus ticks were collected: 1515 in year 2005 and 3262 in year 2006. The collected ticks were pooled into groups from which total RNA was extracted. Viral RNA was detected using real-time RT–polymerase chain reaction (PCR). Ticks infected with TBEV were found in six of eight locations. Viral RNA was detected in 8 of the 230 pools of ticks collected in 2005 and in 14 of the 442 pools collected in 2006. Prevalence of TBEV infection in Slovenian ticks was determined as 0.47%: 0.54% in 2005 and 0.43% in 2006. The detected infection rate in ticks significantly correlates with the TBEV incidence rates in selected areas. Using the method of sequencing, we have confirmed that the TBEV in ticks is genetically related to the TBEV in Slovenian patients.
Introduction
TBE virus (TBEV) is a member of the family Flaviviridae, genus Flavivirus (Kuno et al. 1998, Gubler et al. 2007). There are three main lineages of TBEV: European, Far Eastern, and Siberian subtypes (Lindquist and Vapalahti 2008). Ixodes ricinus is the primary vector of the European subtype of TBEV, whereas the other two subtypes are transmitted by Ixodes persulcatus.
TBE distribution in Europe is very focal, that is, the virus circulates in the forest ecosystems between the tick and vertebrate populations (Korenberg 1994, Randolph et al. 1999). Previous studies have shown that the prevalence of TBEV infection in unfed I. ricinus ticks in Europe ranges substantially from 0.1% to 5% (Danielova et al. 2002, Suss et al. 2006). Of all tick species present in Slovenia, I. ricinus is the most widespread and frequent (Tovornik 1988). Although I. ricinus is a recognized vector of TBEV in Slovenia, no information exists about the prevalence of TBEV infection in ticks. In addition, genetic relationship between TBEV in ticks and TBEV in Slovenian patients has not been confirmed.
The aim of this study was therefore to establish the prevalence of TBEV infection in I. ricinus ticks and to determine how the gathered information correlates with the epidemiological data on TBE in Slovenia. For this purpose we conducted a 2-year survey in 2005 and 2006, during which we were collecting I. ricinus ticks monthly in eight different locations in Slovenia. Using molecular techniques we characterized the virus from ticks and compared it with the TBEV sequences obtained from Slovenian patients.
Materials and Methods
Tick sampling
Questing I. ricinus ticks (adults and nymphs) were collected in eight different locations over a period of 2 years (between April and November in 2005 and between February and December in 2006) at approximately the same time of month. Two researchers collected ticks by dragging a 1-m2 white cotton cloth over the lower vegetation at peripheral and inner parts of forests, including suburban parts and picnic areas at eight different locations in Slovenia. Locations were selected according to the endemic foci of the TBE (Table 1). We also selected locations that had the same prevalence of infection, but they differed in altitude, for example, Rakovnik (322 m) and Osolnik (817 m), or Kamnik (590 m) and Štefanja gora (685 m). Collected ticks were pooled into groups, which consisted of 5 adults (males and females separately) and 10 nymphs. The adults were first cut in half. One half was used for pooling, and the remaining half was stored in RNAlater® (Ambion, the RNA Company, Austin, TX) at −20°C. Prior to pooling, the ticks were washed in 70% alcohol for 5 min and then rinsed in sterile water.
Level of endemicity was based on TBE incidence in the region where tick sampling was conducted and it was determined according to 2000–2009 TBE incidence data.
Average incidence of TBE in the municipality where tick-sampling was conducted, in the years of sampling 2005–2007.
TBE, tick-borne encephalitis.
Molecular detection
According to the manufacturer's instructions, total RNA was extracted from each pool by following the modified guanidinium thiocyanate–phenol–chloroform procedure (Chomczynski and Sacchi 1987) with Trizol® Reagent (Invitrogen Life Technologies™, Carlsbad, CA). The pooled ticks were homogenized in 300 μL phosphate-buffered saline. From each pool, 100 μL was then used for RNA extraction. The resulting pellets were dried, dissolved in 30 μL of RNAsecure™ (Ambion), and stored at −20°C.
We detected viral RNA by real-time RT–polymerase chain reaction (PCR) following the protocol published by Schwaiger and Cassinotti (2003). In short, the 3′ end of TBEV genome was amplified and detected using specific TaqMan probe labeled with FAM. RNA from positive pools was afterward amplified with RT-nested PCR to prepare amplicons for sequencing. We amplified a 252-bp-long segment of the highly conserved NS5 protein of the TBEV genome following the method previously described (Puchhammer-Stockl et al. 1995, Saksida et al. 2005). In addition, NS5 amplicons of TBEV from Slovenian patients obtained in a previous study were also sequenced (Saksida et al. 2005). Sequencing was done on both strands of DNA using commercial reagents “BigDye® terminator cycle sequencing ready reaction kit” (ABI Prism, PE Applied Biosystems, Foster City, CA). The sequences were analyzed with computer programs SeqMan and EditSeq of the Lasergene 1999 software package (Dnastar, Madison, WI). Sequence alignments were made by MegAlign program based on ClustalW algorithm.
Genetic relationships of TBEV from ticks and patients were established with a software package TREECON (TreeCon for Windows v1.3b; University of Konstanzu, Germany). Phylogenetic tree was constructed by neighbor-joining method. The topology of the tree was obtained with Kimura 80 model and support for the nodes was calculated with 1000 bootstrap replicates.
Statistical analysis
Prevalence of TBEV was analyzed according to independent variables such as sampling site, years, and tick stage by chi-square or Fisher's exact tests. Significance was set at p < 0.05. Pearson's chi-square test was calculated to estimate the significance of differences between prevalence of infection in ticks in different regions of Slovenia and incidence of TBE in corresponding regions. The statistics were performed using SPSS 17.0 (SPSS, Chicago, IL).
Results
Altogether we tested 4785 I. ricinus ticks: 1517 collected in the first and 3268 collected in the second year in eight different parts of Slovenia. In 2005, 55 pools were formed from 188 male ticks, 49 pools from 145 females, and 126 pools from 1184 nymphs. In 2006, 95 pools were formed from 431 male ticks, 97 pools from 429 females, and 250 pools from 2408 nymphs.
Ticks infected with TBEV were found in six of eight locations (Table 1). No TBEV-infected ticks were found on locations 1 and 3.
Viral RNA was detected in 8 of the 230 pools of ticks collected in 2005 and in 14 of the 442 pools collected in 2006 (Table 2).
Bolded numbers represent pools and individuals amongst which TBEV was detected, and infection rates higher than zero.
Program PooledInfRate version 3.0 (a Microsoft® Excel Add-In, developed by Brad Biggerstaff; CDC, Fort Collins, CO) was used to compute the infection rates from pooled data. The overall prevalence of TBEV infection in ticks in Slovenia was calculated as 0.47%: 0.54% in 2005 and 0.43% in 2006, but the difference in the overall infection rate between years was not statistically significant (p = 0.262885). When comparing the infection rates in specific sites between years, only location 7 showed statistically significant differences (p = 0.002087) (Table 2). The highest infection rate in 2005, namely 1.08%, was found in males, followed by 0.71% in females and 0.43% in nymphs. In 2006, the infection rates were 0.70% for both males and females and 0.34% for nymphs. The differences between infection rates in adults and nymphs were statistically significant (p = 0.0002339). The TBEV infection rates of ticks collected in different locations in 2005 and 2006 are presented in Table 2. Sampling site 2 had a significantly higher infection rate than the average in Slovenia (p = 0.002804), and locations 1 and 3 had a significantly lower infection rate (p < 0.00001), because no TBEV was detected in ticks in this site.
We sequenced eight selected amplicons obtained with RT-PCR in 2005, seven distinctive amplicons from 2006, and six amplicons from the previous study (Saksida et al. 2005). Amplicons for sequencing were selected with respect to different tick sampling sites and different tick stages within one sampling site. Primer sequences were removed from generated sequences. Therefore, 212-bp-long sequences of NS5 protein were compared for identity in BLAST database, revealing the highest similarity with European strains of TBEV.
Phylogenetic analysis demonstrated three groups of TBEV sequences from ticks and patients according to the geographic part of Slovenia. The sequences from ticks were identical within a geographic region, but differed from the sequences in other locations. The sequences of TBEV from ticks matched with the TBEV detected in Slovenian patients. The percentage of identity among TBEV sequences included in the alignment is shown in Figure 1a. Among three groups, Slovenian TBEV sequences from the first group were most similar to Isosaari strain (AJ298323). The second group was closely related to Neudoerfl (U27495) and TBE263 (U27491). The third group formed a separate cluster (Fig. 1b).
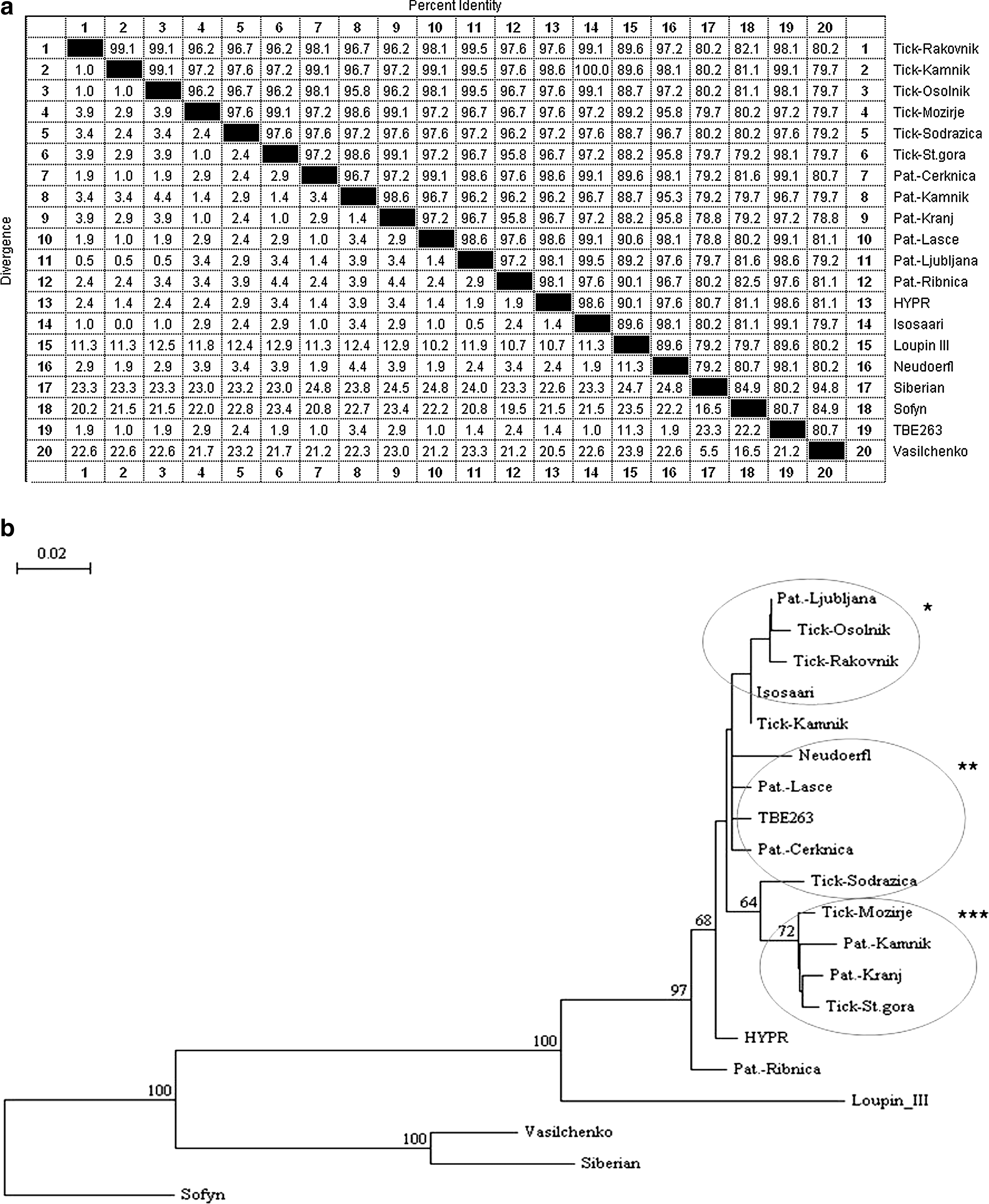
Phylogenetic relationships of the tick-borne encephalitis virus (TBEV) isolates from the ticks collected in Slovenia, from Slovenian patients, and from the referent isolates of the virus inferred from multiple sequence alignment of 212 bp (primer sequences were removed).
Unique sequences of TBEV from ticks and patients determined in this study can be accessed under accession numbers: EU057638, EU057639, EU057640, EU057641, EU057642, and EU057643.
Discussion
This is the first study in Slovenia describing the presence of TBEV directly in unfed ticks collected from the environment.
I. ricinus is the most abundant and clinically significant tick species in Slovenia. This tick is recognized as a vector of many pathogens such as borreliae, Anaplasma phagocytophillum, babesiae, and TBEV in Slovenia and other European countries (Strle and Stantic-Pavlinic 1996, Petrovec et al. 1997, Duh et al. 2001). Studies performed around Europe show that 0.1%–5% of ticks are infected with TBEV (Randolph 2001, Oehme et al. 2002, Suss 2003). In our study, we determined that the overall prevalence of TBEV in ticks in Slovenia is 0.47%. However, significant variation in the prevalence of infection depending on the location of sampling and year of sampling was noted.
Overall prevalence was higher in 2005 (0.54%) than in 2006 (0.47%), even though the incidence in humans was higher in 2006 (426 confirmed cases) than in 2005 (375 confirmed cases). The data were obtained from the Laboratory for Diagnosis of Zoonoses, the reference center for microbiological TBE diagnosis in Slovenia. The difference in prevalence rate between 2005 and 2006 can be, in our opinion, explained with much higher number of ticks sampled in 2006 in the locations where the prevalence of TBE in humans is very low (locations 1 and 3; Table 1). Comparing the overall prevalence of TBEV in ticks in Slovenia with other countries in Europe, low prevalence of TBEV in ticks was determined. At the first glance, this is somewhat unlikely because the incidence of TBE in Slovenia is one of the highest in Europe (Donoso Mantke et al. 2008). Therefore, it is necessary to analyze the data in a more detailed way. Several explanations could resolve this inconsistency. One explanation for low prevalence of TBEV in ticks in Slovenia is the occurrence of microfoci, which arises because the feeding of larval and nymphal ticks depends on small mammals that often live in relatively small areas. Subsequently, the circulation of the virus is dependent on those limited areas (Rogers 1988). Even though our selected locations represent some of the areas with the highest number of TBE patients, it might be that the microfoci areas and our sample sites, which were only 100 m2, did not overlap.
The other possible explanation is the prevalence in specific sites, which also show significant variation. In all the sites with high or medium incidence of TBE in humans (Table 1), TBEV was detected in ticks at least in 1 year (Table 2). In both locations with low incidence in patients, we found no positive ticks. Also looking at specific sites, especially in the second year of our analysis when the TBE incidence in humans was one of the highest in the past years, we can see a very good correlation. The three locations (2, 4, 7; Table 1) with the highest prevalence of TBEV detected in ticks are also the locations where there was the highest incidence of TBE in patients comparing all selected locations. Statistical analysis confirmed significant correlation between prevalence of TBE in ticks in specific regions with the incidence of TBE (p = 0.026).
The differences were also noted in the prevalence of infection between the adults and the nymphs. The overall infection rate in the adults was 0.76% and in the nymphs 0.37%. The reason for higher prevalence of TBEV in the adults than the nymphs could be that the ticks were collected from vegetation and were therefore unfed. They acquired the infection via feeding in the previous stages and transtadially sustained it to the next one. As the adults fed twice and the nymphs only once before, they had a greater chance to get infected (Nuttall and Labuda 1994). But, it has been implied before that the majority of ticks get infected in the larval stage, which may indicate that the infection rate should be the same, presuming the infection with the virus does not influence longevity. Another explanation might be that the viral load is bigger in adult ticks and therefore easier to detect, because viral multiplication occurs at the times of blood meals, which is double in case of adults.
Using the method of sequencing, we have confirmed that the TBEV in ticks is genetically related to the TBEV in Slovenian patients and also that they are geographically related (Figs. 1 and 2). They cluster within the European TBEV group together with TBE263 (U27491), Isosaari (AJ298323), and Neudoerfl (U27495) isolates. Clustering of Slovenian strains of TBEV together with the central European and Finnish viruses is not surprising as Slovenia is also a central European country (Fig. 1b). Although a short segment of NS5 protein was analyzed, a notable diversity of the TBEV was demonstrated. However, for proper phylogenetic analysis, sequencing of larger part of the TBEV genome from the patients and ticks in Slovenia is needed.
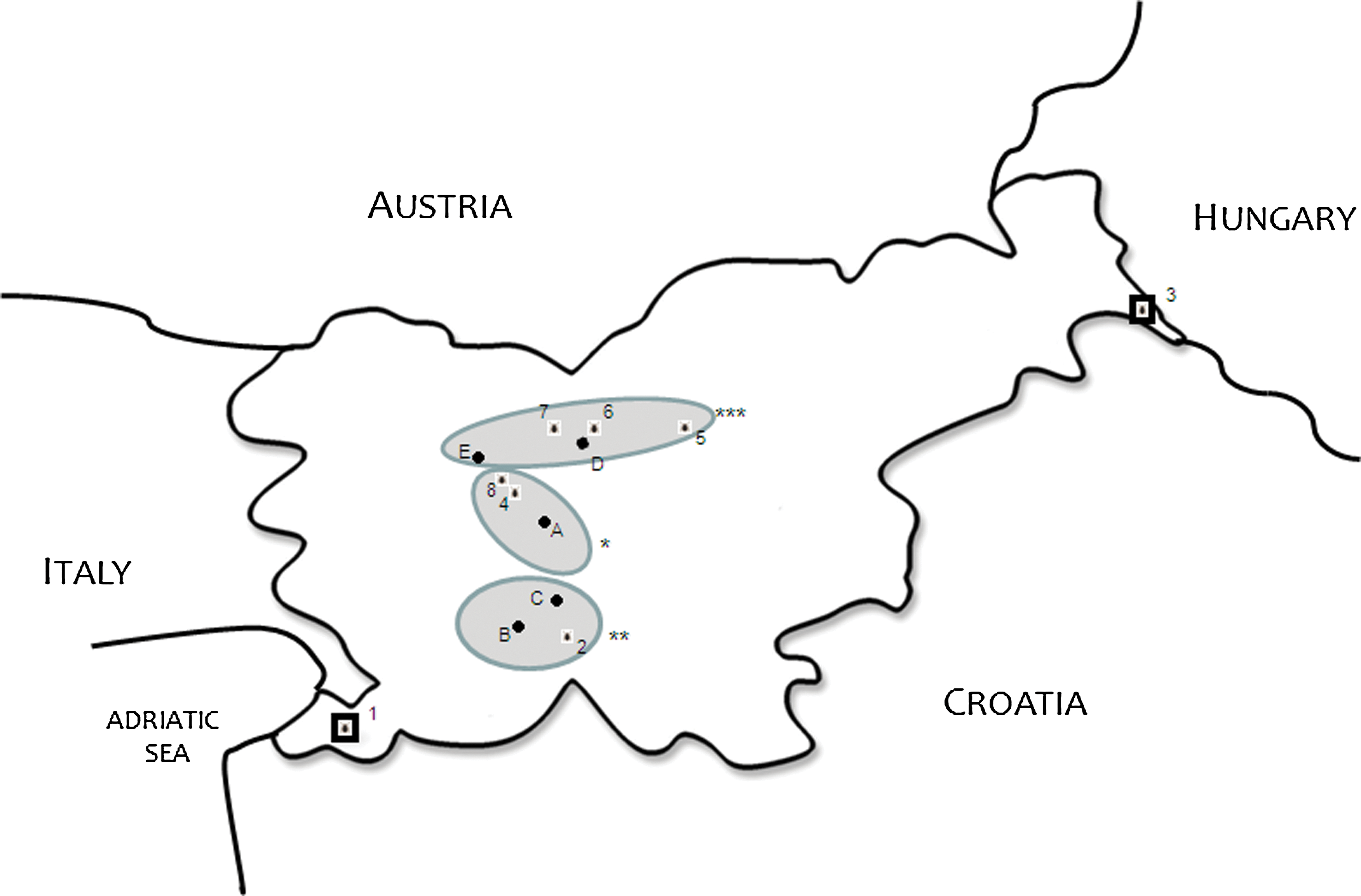
Geographic distribution of sampling sites and analyzed Slovenian patients. (*Central Slovenia (A patient Ljubljana; 3 sampling site Rakovnik; 4 sampling site Osoinik); **South-Western Slovenia (B patient Cerknica; C patient Lašče; 2 sampling site Sodražica); ***Northern Slovenia (D patient Kamnik; E patient Kranj; 7 sampling site Štefanja gora; 8: sampling site Kamniška Bistrica); 1 sampling site Črni kai; 8 sampling site Murska šuma).
In conclusion, the determined prevalence of TBEV infection in I. ricinus ticks collected in Slovenia is 0.47%. The adults and nymphs are infected with TBEV that is genetically related to TBEV detected in Slovenian patients. Infected ticks were found only in those locations of Slovenia where the present prevalence of TBE is determined as medium or high.
Footnotes
Acknowledgments
The authors thank Katja Strašek Smrdel for her help in accessing and interpreting data. This study was partially funded by EU grant GOCE-2003-010284 EDEN. This article is catalogued by the EDEN Steering Committee as EDEN0147 (
Disclosure Statement
No competing financial interests exist. The contents of this article are the sole responsibility of the authors and do not necessarily reflect the views of the European Commission.