
Abstract
So far, data on the natural cycle of rickettsiae of the tick-borne spotted fever group (SFG) in Central Europe are barely available. Some studies showed the occurrence of different Rickettsia species in their arthropod vectors, but it is unclear which animals might have any kind of reservoir function. This survey was therefore set up to provide information on the occurrence of SFG rickettsiae in small mammals in Germany. A total of 124 rodents and insectivores were collected over a period of 3 years in Lower Bavaria, South-Eastern Germany. Screening for Rickettsia antibodies was performed using immunofluorescence with Rickettsia conorii and R. helvetica slides, and the comparability of sera and body fluids (transudates) was investigated in these assays. Further, real-time polymerase chain reaction (PCR) was used for screening of Rickettsial DNA in rodents and insectivores. Ear versus liver tissue was compared to evaluate the more suitable tissue for detection of specific DNA. Further, a new PCR targeting the 18S ribosomal nucleic acid was established as internal control. The results indicated that transudates are a sufficient alternative to proof infection in cases where no sera are available. Rickettsial DNA, that is, Rickettsia felis and R. helvetica, was found in seven animals with the ears proving to be a proper choice for PCR. Statistical analyses revealed that the presence of ectoparasites and the body size positively correlated with the occurrence of rickettsial DNA. Overall, our study suggests that rodents and other small mammals may act as reservoir hosts for Rickettsia. However, with the course of infection and its transmission in wild animals still unknown, further investigations are needed to better understand the natural cycle of SFG rickettsiae.
Introduction
Materials and Methods
Trapping of small mammals
Rodents were collected using Sherman live traps in October 2004 and May and September 2005 at seven locations in the district of Lower Bavaria, including forests, pasture, blackberry bushes, and farmland (Fig. 1). Sampling areas were chosen according to the occurrence of Hantavirus-induced Nephropathia epidemica cases and trapped by a decoying method as described in Essbauer et al. (2006) and Mertens et al. (2009). In September and October 2008, trapping was performed at a total of 22 sites ranging from 300 up to 1400 m altitude (a.s.l) in the German part of the Bohemian Forest. Sixteen Sherman live traps were deployed in an 18 m2 plot, one trap every 3 m for three consecutive nights, and were checked twice a day. Animals were anesthetized by CO2 exposure and killed humanely according to the German Animal Protection Act, after blood was drawn by heart puncture. Trapping site, species, sex, reproductive, and physical conditions, including parasitic load, location of ectoparasites, and also ectoparasite species, were recorded for each rodent.
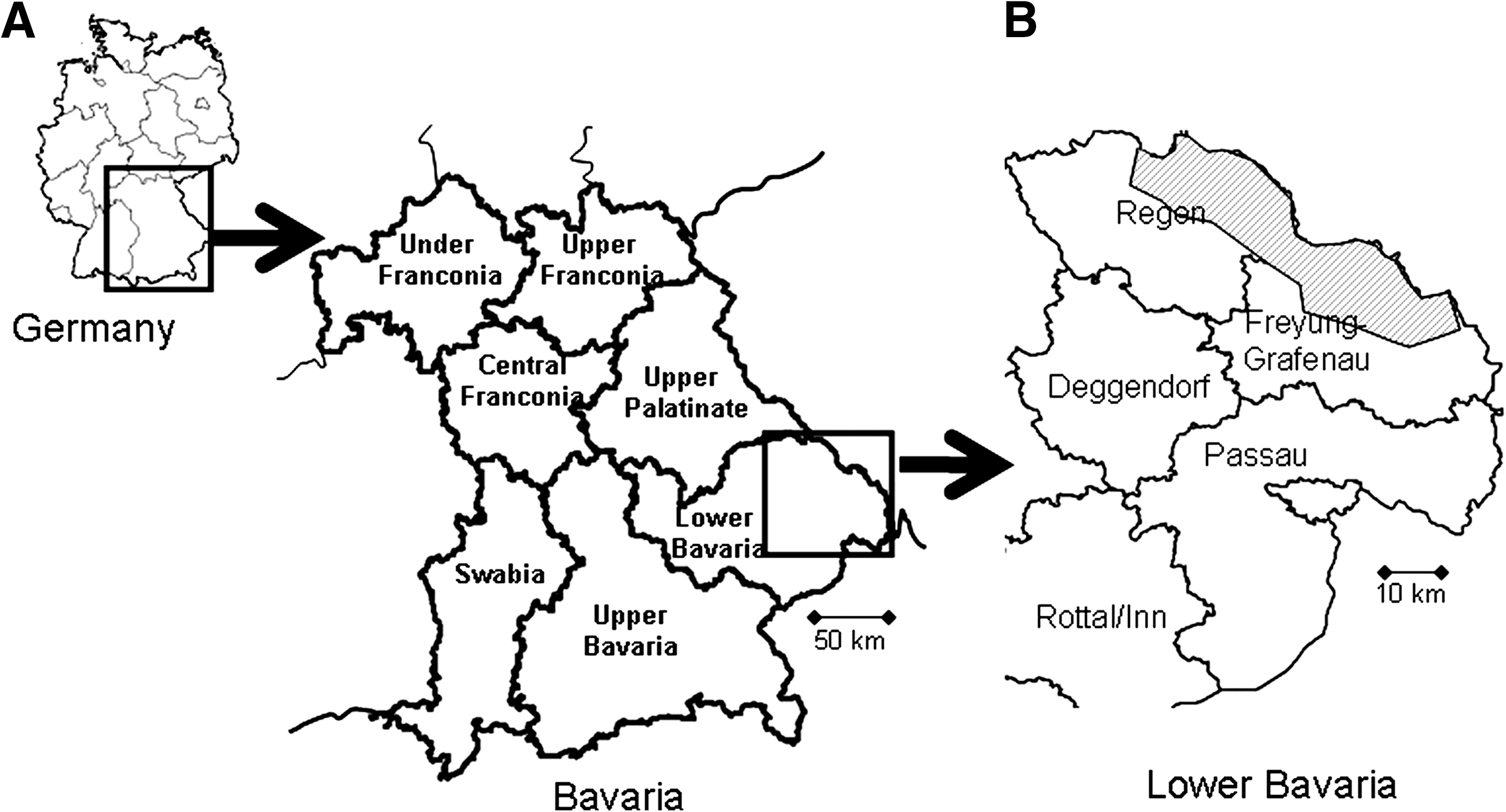
Geographic location of trapping sites in South-East Germany.
Preparation of tissue samples
Rodents were dissected under BSL-2 conditions. Tissue samples (heart, lung, liver, spleen, kidneys, and ears) were aseptically removed. Extracted viscera were put into Lysing matrix tubes A (MP Biomedicals), mixed with minimal essential medium (MEM) supplemented with 3% fetal calf serum (Biochrom AG), and homogenized with the Fast Prep 120 instrument (Biogene) with a power of 5.0 m/s for 30 s.
Serological investigations
Rodent blood was centrifuged at 285 g for 10 min at 4°C. The derived sera were stored at −40°C until use. Sera were preliminary diluted 1:20 in phosphate-buffered saline (PBS), as this dilution proved to be the most suitable with regard to specificity and sensitivity for screenings in preliminary tests. As there were blood samples missing from several mice that could not be captured alive, the heart of each animal was rinsed with 1 mL PBS, with the resulting suspension (referred to as transudate) tested additionally or instead where no serum was available.
Sera and transudates of wild mice were screened with an in-house immunofluorescence test for detection of immunoglobulin G (IgG) antibodies against R. helvetica. R. helvetica (strain AS 819, own unpublished strain) was propagated on Vero E6 cell lines for 27 days at 34°C. Infected cells were trypsinated and centrifuged at 2000 g for 5 min. The pellet was resuspended in PBS and 10 μL each spotted on 10-well antigen slides (Biomérieux). After air-drying, cells were fixed with a 50% methanol/50% acetone (1:1) solution. Further, R. conorii, which is phylogenetically distinct from R. helvetica in the SFG, was used for immunofluorescence tests. It is also crossreactive with Rickettsia from the SFG but not endemic in Germany. Slides coated with R. conorii–infected cells were obtained from a commercial supplier (Rickettsia conorii immunofluorescence assay [IFA] IgG Antibody Kit; Fuller Laboratories RCG-120).
Both Rickettsia IFA were conducted with a polyclonal rabbit anti-mouse serum IgG/fluorescein isothiocyanate as a conjugate (dilution 1:20; Dako) together with Evans blue counterstaining. As positive/negative control served the controls provided by the commercial available kit as well as formerly positive tested mouse sera and PBS, respectively, were used. Slides were read by two independent examiners using a fluorescent microscope.
Nucleic acid isolation
Nucleic acids (NA) were extracted using the QIAamp Viral RNA Mini Kit (Qiagen) following the manufacturer's instructions. This test was used because of investigations regarding RNA–viruses conducted in parallel. Moreover, it is comparable with other standard DNA extraction kits regarding the quality of the extracted NA.
For extraction of rickettsial DNA from ears, one ear per mouse in the aggregate was placed into a 1.5 mL Eppendorf tube and digested using the QIAamp DNA Mini Kit (Qiagen) as described in the instruction manual. If not analyzed immediately, the extracted NA were stored at −20°C.
Polymerase chain reactions
Rickettsial polymerase chain reactions
A real-time (RT) polymerase chain reaction (PCR) targeting citrate synthase (gltA) was performed using the LightCycler FastStart DNA Master HybProbe System (Roche) for LightCycler 1.5 following the protocol published by Wölfel et al. (2008). An uracil–DNA–glycosylase (UDG) incubation step was added to prevent the re-amplification of carryover PCR products between reactions. Briefly, 0.5 U UDG Mix (Roche) was added to each reaction, and the protocol was modified including a preincubation step for UDG digest at 40°C for 10 min.
A conventional PCR was performed, targeting the partial outer membrane protein B (ompB) by following the protocol published by Roux and Raoult (2000). The protocol was then modified with annealing steps. Temperatures were gradually reduced in two-degree steps from 60°C to 50°C (touchdown). For this, Platinum Taq Polymerase High Fidelity (Invitrogen) was used, adding 5 μL of DNA to each reaction.
Mammalia species-specific PCRs
To investigate if there was an inhibition of the PCR (internal control), we established a universal mammalian-specific 18S ribosomal RNA gene PCR ahead of Rickettsia-specific PCR amplification. For choice of sequences, different mammalian 18S rRNA sequences were obtained from the GenBank (Table 1), and aligned by ClustalW2 (Embl-EBI,
In 2004/2005, for 23.4% (n = 19/81, Hantavirus-positive animals of a previous study), and in 2008, for all animals species determination was confirmed by both PCR and subsequent sequencing of partial mitochondrial cytochrom B-(mtCytB) gene (Essbauer et al. 2006).
Analysis of PCR products
OmpB and mtCytB PCR products were loaded on a 1.5% agarose gel and analyzed by UV illumination. Amplificates were purified using the QIAquick Gel Extraction Kit (Qiagen) and sequenced by the GATC sequencing service (Konstanz) using corresponding specific primers. Sequence and phylogenetic analyses were performed using BioEdit (Version 7.0.0, Copyright ©1997–2004) (Hall 1999).
Statistical analysis
Data of trapped rodents were collected and evaluated with R 2.9.2 (R Development Core Team 2009). Serological assays were compared using concordance comparison and kappa-statistics. Descriptive statistics was performed for ompB-PCR-positive samples for body size (head-body length), body weight, and organ weights. The number of these PCR-positive samples, that is, samples that could be sequenced for Rickettsia species determination, was low and restricted to seven animals in two rodent species (Myodes glareolus and Apodemus flavicollis). Therefore, we restricted our final analysis to these seven animals in our data set comprising 106 animals of the two dominant species. We selected three predictors only, because several of our variables were intercorrelated (e.g., body size and body length). For the other variables, we restrained from a formal test and provided only descriptive statistic. In our final test model with Rickettsia-PCR positivity as dependent variable, we included in the following sequence the predictors: (1) as control variable the species, (2) occurrence of ectoparasites, (3) body size, and (4) elevation. We applied a sequential analysis of deviance table (Hastie and Pregibon 1992). That is, the reductions in the residual deviance as each term of the formula is added in turn are given in as the rows of a table, plus the residual deviances themselves. We first added the species to the model and then subsequently ectoparasites, body size, and elevation. Due to the binomial dispersion of our dependent variable Rickettsia-PCR positive, we applied the chi-squared test.
Results
In 2004/2005 a total of 81 animals were trapped in Lower Bavaria: 21 yellow-necked mice (A. flavicollis), 7 wood mice (Apodemus sylvaticus), 47 bank voles (M. glareolus), 4 brown voles (Microtus agrestis), 1 water vole (Arvicola amphibius), and 1 house mouse (Mus musculus).
In 2008, a total of 43 animals were trapped in the Bohemian Forest: 15 bank voles (M. glareolus), 23 yellow-necked mice (A. flavicollis), 4 brown voles (M. agrestis), and 1 Eurasian pygmy shrew (Sorex minutus). Table 2 summarizes trapped animals and available samples for all serological and molecular Rickettsia assays.
Rickettsia serology
Of the 2004/2005 micromammalia, 23 out of 71 (32%; no serum or transudate available for 10 animals [Table 3]; animals listed in Table 4) tested rodents showed serological evidence of a past infection with rickettsiae. Among these are 30% (n = 12/40) bank voles (M. glareolus), 21% (n = 4/19) yellow-necked mice (A. flavicollis), 66.7% (n = 4/6) wood mice (A. sylvaticus), and 50% (n = 2/4) field voles (M. agrestis). In summary, 18.3% (n = 13/71) of the samples showed a reaction in the R. conorii IF assay and 21.1% (n = 14/71) against R. helvetica. There were five animals that seroreacted in both assays (Tables 3 and 4).
For detailed data of serological reactive sera of rodents, see Table 4.
Pos, positive; RT-PCR, real-time polymerase chain reaction.
13.8% (n = 4/29*) of sera.
15.5% (n = 11/71) of transudates.
32.0% (n = 8/25*) of sera.
11.3% (n = 8/71) of transudates.
No amplification in opmB-PCR is possible.
27.3% (n = 6/22) of sera.
16.3% (n = 7/43) of transudates.
40.9% (n = 9/22) of sera.
9.3% (n = 4/43) of transudates.
The asterisk (*) in the above footnotes indicates that numbers differ as for four sera there was not sufficient serum available for both assays.
F, female; M, male; juv, juvenile; NT, not tested (no or insufficient serum); Pos, positive; Neg, negative; Undil, undiluted serum; ND, not determined.
From small mammals collected in 2008, immunofluorescence tests for antibodies against R. conorii revealed 20.9% (n = 9/43) reactive sera/transudates. For animals where both serum and transudate were available for testing (n = 21), body fluids were tested in several dilution steps. For the rest of samples (n = 22), the investigation was refined to just transudate. Serum titers ranged from 1:20 to 1:160 (DO 08/009, M. agrestis), with transudates showing positive reactions, ranging from undiluted up to 1:16. Two animals with reactive sera were tested negative for transudates and two tested positive. In comparison, reactivity in the R. helvetica immunofluorescence was found in 10 samples from 10 rodents (23.3%), with serum titers ranging from undiluted up to 1:320 (DO 08/009, M. agrestis).
To evaluate and compare the results of the R. helvetica and the R. conorii IFA, we counted negative and seroreactive samples that revealed the same reactivity in both assays (in summary, 52 sera and 65 transudates). Of 52 tested serum samples, 39 reacted concordantly in the R. helvetica as well as in the R. conorii assay. Therefore, we obtained a high concordance. Nevertheless, the kappa value was rather low (k = 0.35) and the confidence interval rather broad (0.05 to 0.66), due to the low and unequally distributed sample numbers. For transudates, 57 of 65 tested samples were concordant (k = 0.43, confidence interval 0.06 to 0.80). In conclusion, with both IF assays, comparable results for Rickettsia antibody detection seemed to be obtained. Details of the serological results are summarized in Tables 3 and 4.
PCR results
Of the 2004/2005 micromammalia, 5.2% (n = 4/76) tested samples were found positive in RT-PCR, targeting rickettsial gltA (i.e., one M. glareolus and three A. flavicollis). In 2008, 5 out of 43 (11.6%) micromammalia showed amplification, including 1 M. glareolus, 1 M. agrestis, and 3 A. flavicollis. In all PCR-positive animals, the ear was found positive with cycle thresholds (CTs) between 36 and 50. RT-PCR of DNA extracted from liver tissues (n = 46) showed that 100% for the 2008 sampling and three ear-positive animals for the 2004/2005 sampling had no positive results. Two ear DNA samples with CT >40 in the gltA-RT-PCR revealed no amplificate in the ompB-PCR. For the other seven samples Rickettsia species was determined using ompB-PCR, followed by sequencing and resulted in five R. felis and two R. helvetica. R. felis was found in four A. flavicollis (three adult males and one adult female, 2008) and in one M. glareolus (adult male, 2004). R. helvetica was found in one A. flavicollis and in one M. glareolus (both adult males, 2004/2005). Obtained R. felis or R. helvetica ompB nucleotide sequences were 100% identical among each other and to the respective type species deposited in GenBank. GenBank accession numbers are GU324464–GU324470. Results of the molecular biological screening of rodents are shown in Tables 3 –5.
n.a., no amplificate in ompB-PCR.
Ten percent of all DNA extracted from livers and ears were tested for signs of possible inhibition with the 18s rRNA Sybr Green PCR. Those 10% included animals of M. glareolus, M. agrestis, A. flavicollis, and S. minutus. As expected, all samples revealed no inhibition, proving that negative samples were not false-negative.
Body conditions of PCR-positive animals
Descriptive statistics showed that there seem to be differences in body weight, body length, spleen, kidney, gonad, and liver weights for the seven partial ompB-sequenced animals (for details, see legend of Fig. 2), but not for seroreactive ones (data not shown).

Box plots. Differences of mean body weight, mean body length, mean liver, spleen, kidney, and gonad weights to ompB-positive polymerase chain reaction (PCR) results. OmpB-PCR-positive animals (n = 7) seem to have higher mean body size (26.4 ± 5 g vs. 19.7 ± 6.1 g) and length (11.3 ± 3.6 cm vs. 8.6 ± 1.2 cm), and higher liver (1.55 ± 0.34 g vs. 1.15 ± 0.44 g), spleen (0.27 ± 0.15 g vs. 0.15 ± 0.11 g), kidney (0.57 ± 0.16 vs. 0.38 ± 0.19 g), and gonad weights (0.22 ± 0.15 g vs. 0.11 ± 0.18 g) than PCR-negative animals (n = 99).
There seemed to be no difference in Rickettsia prevalence in A. flavicollis (n = 44) and M. glareolus (n = 58). After correcting for possible effects of species (Table 6), the parasites had a significant influence on the occurrence of Rickettsia (all of the ompB-PCR-positive animals carried ectoparasites). Beyond these effects even the body size as a surrogate for old animals was still significant (Table 6), while elevation had no additional effect on the occurrence of Rickettsia. Thereby, our analysis supports hypotheses 1 and 2, but hypothesis 3 has to be rejected.
Significant p-values are shown in bold.
Discussion
In this project we used a method of live-trapping that has been widely field-tested in the last years to investigate rodent-associated pathogens (Ulrich et al. 2008, 2009). Aim of the study was to provide information on the occurrence of SFG rickettsiae in small mammals in southern Germany. To compare serum and transudates, we used two different immunofluorescence assays, one for R. conorii and one for the detection of R. helvetica. Antibody reactivity to the rickettsial antigen is group specific. A marked crossreactivity is seen in the IFA procedure between members of the SFG (e.g., R. conorii, R. rickettsii, R. felis, and R. helvetica) (Fuller Laboratories, R. conorii IFA Antibody Kit Instruction manual). Therefore, the commercial available R. conorii assay was used, although R. conorii itself is not expected to occur in Germany. However, the phylogenetic relation between R. conorii and R. helvetica is quite distant; hence, detection of both antibodies with only one assay could not be assured (Roux and Raoult 2000). We therefore conducted a specific R. helvetica assay.
Twelve of 41 reactive sera/transudates (29.3%) reacted in both assays (Table 4). There was a small difference for the rodent samples between R. helvetica and R. conorii slides for IFA for 2008. Therefore, IFA screenings are a useful tool to assess past infections; however, they are not sufficient enough to differentiate Rickettsia species, offering future research potential.
So far nothing is known on Rickettsia IFA cut-off or even possible Rickettsia-neutralizing antibodies in wildlife rodent samples. Usually, screening of wild rodent is started with undiluted sera/transudates and after a specific reactivity a titration of sera/transudates is performed. We had at least four samples in 2004/2005 and six samples in 2008 with titers ≥1:40. Further, the comparison of antibody titers showed that in 9 out of 33 reactive samples both serum and transudate reacted (Table 4), that is, also confirming a specific reaction in IFA for low titers. Samples collected in 2008 showed higher reactivity and accordance than the 2004/2005 samples, which might be due to several freeze–thaw cycles of the older samples. Other studies showed the comparability of body fluids to serum in serological investigations: for the detection of antibodies against Trichinellosis in two enzyme immunoassays, tissue fluid of slaughtered swine showed equally effective as both blood and serum (Gamble and Patrascu 1996). Muscle transudate samples of pigs were also suitable in anti-PRRSV (porcine reproductive and respiratory syndrome virus) ELISA and IFAT tests (Molina et al. 2008). Moreover, the persistence of antibodies against Microsporum canis in decaying fox carcasses implied a positive serological result from blood or body fluid (Tryland et al. 2006). In rodents the comparability of serum and transudate was also shown for Hantavirus ELISA, Western blot, and IFA tests (Essbauer et al. 2006). However, more sampling should be conducted to further test this relationship. Finally, rinsed heart fluid was an acceptable alternative where no or only small amounts of serum are available. However, the best results are obtained with serum samples of live-captured animals.
Of seven PCR-positive samples (only the ompB-PCR-positive animals were considered securely positive), only two reacted in the IFAT for the detection of IgG antibodies. In 40 rodents, we found IgG antibody reactivity but in only two of these, rickettsial DNA. Other studies examining veterinary clinic cats infested with R. felis–infected cat fleas (Ctenocephalides felis) with and without fever were not able to amplify R. felis DNA in whole blood samples but found antibodies against R. felis (Hawley et al. 2007, Bayliss et al. 2009). Kidd et al. (2008) found R. rickettsii and R. conorii DNA in naturally and experimentally infected dogs, in some of them before seroconversion. Specific IgG antibodies are known to appear after 7 days the earliest, mostly after the onset of symptoms, and show a possible past infection. Cats that were exposed to fleas infected with R. felis seroconverted by 4 months postexposure (Wedincamp and Foil 2000). Dogs experimentally infected with R. rickettsii showed an IgM antibody response between days 9 and 80 postinfection, and an IgG antibody response starting between days 22 and 28 postinfection and lasting until at least day 130 postinfection (Breitschwerdt et al. 1990). It is still unknown what amounts of rickettsiae are required for a vertebrate host to create a strong antibody response. As the time frame where bacteremia and antibody response overlap is usually quite short, and taking more than one single blood sample per mouse is not possible for our study design, more samples of wild rodents are necessary to further evaluate these findings and to possibly find more animals where both antibodies and rickettsial DNA can be detected. Our findings raise questions about the duration of a rickettsial infection and whether it is persistent in rodents or not. This again queries whether mammals can be a reservoir host.
This study is ground breaking in that we find hints that small animals acted as reservoir hosts for Rickettsia as known for other pathogens like Borrelia spp. (Piesman and Gern 2004), Bartonella spp., or tick-borne encephalitis virus. Some studies found rickettsiae in arthropods collected from small mammals (Chen et al. 2002, De Sousa et al. 2006). However, there are only very few studies regarding rickettsiae in wild animals. In the Sierra Nevada one golden-mantled ground squirrel was found PCR positive for R. rickettsii (Adjemian et al. 2008), and in California three opossums were found positive for rickettsiae (Williams et al. 1992). In blood of rodents from Peru 2 out of 24 (8.3%) were found PCR positive for R. felis (Richards, personal communication). In our study sequencing of amplified ompB products revealed Rickettsia spp. infections in seven rodents. Two of them were R. helvetica, which were already described in ticks collected from rodents (Spitalská et al. 2008) and are known to be endemic in southern Germany (Pichon et al. 2006, Wölfel et al. 2006, Silaghi et al. 2008). Five of them belong to R. felis, a species that so far seemed to involve particularly cats and several flea species in its natural cycle (Reif and Macaluso 2009).
We herein also for the first time compare the suitability of wild rodent liver versus ear tissues for the detection of rickettsial DNA. Rickettsial DNA was detected in seven samples of rodent ears, whereas in no liver sample. Williams et al. (1992) found Rickettsial DNA in spleens of opossums. In another study DNA was taken from whole blood samples with 1 positive sample out of 89 (Adjemian et al. 2008). In a Spanish investigation, pools of tissues and ears were tested by multiplex PCR for evidence of SFG rickettsiae, with no positive result (Barandika et al. 2007). As we did not collect EDTA blood, we tried to test DNA isolated from blood cell pellets in serum separating tubes of the 43 animals trapped in 2008. However, we did not get any positive PCR signal in these blood samples.
Our results suggest that ear tissue was more sensitive than liver tissue. A reason for that might be the sequestration of the Rickettsia organism in extremities of the body, with a lower temperature like ears or tailtip. The optimum growth temperature for Rickettsia in vitro was shown to be at 32°C (Pornwiroon et al. 2006), which might explain the good growth in body extremities where also the immune response may be less effective. Another reason might be a more local infection without rickettsemia, a hint for that would be that ticks were often found in the ear region.
We developed a new tool for internal control of tissue from wild mammals. Others describe the use of standard housekeeping genes like GAPDH, β-actin, β2-microglobulin, cytochrom B, and as well rRNA (Livak and Schmittgen 2001). Selvey et al. (2001) reported that 18S rRNA expression offered superior consistency to that of β-actin under most circumstances and state that its expression appears to be a consistent measure of total RNA in a sample throughout the cell cycle.
There are only very few studies that analyze data on rodents and their associated pathogens statistically as it was done for Cowpox virus infection by Essbauer et al. (2009). However, statistical analyses are useful to comprehend interrelations between infection courses of (reservoir-) hosts and ecological aspects. No formal sample size estimation was performed before the experiments, because our sampling of mice in a bad season was not evaluable and no literature about the expected effect was available. However, the sample size for the logistic model analysis is reasonable large to test the two parameters of interest and, obviously, the power was large enough to reject the hypothesis. As Rickettsia are transmitted by arthropods such as ticks, fleas, and mites, the ectoparasitic infestation of animals was of special interest. We quite often found Ixodes ticks (nymphs and larvae) on the investigated small mammals. Their preferred body spots seemed to be the ears, the chin, and the tail base, presumably because the lack of fur there (data not shown). Fleas found consisted of five different species, mostly Ctenophthalmus agyrtes. It is conceivable that there was a correlation between Rickettsia-positive animals and their parasitic load.
There seemed to be a difference between body length, weight, and several organ weights. PCR-positive animals were heavier and taller, and their organs were usually heavier, which probably is a hint for mainly adult animals being infected. The older the animal, the more probable is an exposure to Rickettsia and Rickettsia-transmitting vectors. Differences in organ weights are also interesting, regarding the question whether infection with Rickettsia has clinical effects on small mammals. Clinical symptoms have been reported in dogs, for example, after infection with R. rickettsii, but not confirmedly in cats or small mammals. Infection with R. felis occurs in humans but has not been confirmed by isolation so far (Pérez-Osorio et al. 2008).
To summarize, we found that IF tests are a useful tool to investigate past infections with rickettsiae, but are unsuitable for further species differentiation. They can be used even with transudates, if there is no serum available, although results are less secure. We found two different rickettsial species, the more common R. helvetica (Dobler and Wölfel 2009) and R. felis, which has not been described in small mammals in Europe before. Rickettsial organisms can be detected best in mammals using ear tissue. The question whether rodents or other small mammals acted as reservoir hosts still needs to be answered. Although we found seroconverted animals and rickettsial DNA in their tissues, we do not know if there is a persistent infection and excretion. However, it is conceivable that infected mammals amplify the cycle by fleas feeding on their infected blood. For this reason, further research specific to this topic, including other rickettsial species endemic in Europe, is necessary.
Footnotes
Acknowledgments
The authors gratefully acknowledge the excellent practical and technical assistance of H. Weber, R. Reiter, D. Eder, R. Terzioglu, D. Woll, and F. Goldberg. M. and D. Kiefer's support in species determination of fleas is highly appreciated. The authors also thank S. Speck for carefully reading the article, E. Seibold for help with statistical analysis, and H. Scholz for critical discussion. This project is supported by a grant from the Bavarian Ministry of Health, Vector-Borne Diseases in Climate Change Investigations (VICCI).
Disclosure Statement
No competing financial interests exist.