
Abstract
Farmed shrimps are vectors of various Vibrio species that are considered a potential health hazard. Previous study has shown that Vibrio parahaemolyticus, Vibrio vulnificus, and Vibrio alginolyticus dominated in the water and larval samples of shrimp hatchery (Randrianarivelo et al. 2010). The effects of two essential oils (EOs) of Cinnamosma fragrans, an endemic plant to Madagascar (B8: linalool-type and B143: 1,8-cineole-type), were determined on the total heterotrophic aerobic bacteria and the Vibrio concentrations in the rearing water of Penaeus monodon hatchery. The assays took place in OSO Farming's shrimp hatchery in Madagascar. EOs were directly added to the water tank. The bacterial concentrations of water tank were assessed on marine agar and thiosulfate citrate bile sucrose agar. The larvae culture corresponded to four replicates each of B8, B143, erythromycin (E), and control (oil and antibiotic free). The bacterial concentration of the rearing water in B8, B143, and antibiotic (E) tanks were significantly lower (p < 0.05) than in the control. Further, there was no significant difference (p > 0.05) between the three treatments B8, B143, and E. This study demonstrated that both EOs of C. fragrans, like antibiotic, inhibited bacterial growth in the rearing water of P. monodon larvae. The potential of C. fragrans EO to control the bacterial load in in vivo conditions of P. monodon hatchery makes it a relevant option for producers to minimize risk of Vibrio growth in the rearing water of larvae, which is the primary source of colonization of shrimp larvae.
Introduction
Therefore, alternative methods need to be developed to maintain a healthy microbial environment in the larval rearing tanks. It is recognized that rearing water is the main source for entry of pathogens in the host (Lavilla-Pitogo et al. 1998). In this regard, plant secondary metabolites, for example, essential oils (EOs), are known to display antimicrobial properties (Direkbusarakom et al. 1998, Dorman and Deans 2000, Elgayyar et al. 2001). The use of EOs of Cinnamosma fragrans, B143 and B8, which were characterized by their high content in oxygenated monoterpenes, 1,8-cineole and linalool, respectively, exhibited, like erythromycin, significant higher survival and lower bacterial concentrations of the larvae of Penaeus monodon than the control (Randrianarivelo et al. 2010). The present study aimed to show the effects of both EOs of C. fragrans on total heterotrophic bacterial and Vibrio spp. population concentrations in the rearing water of P. monodon larvae. Plant extracts have been generally used through oral administration using bioencapsulation, by direct mixture with feed ingredients, or by feeding to Artemia, which acts as a biological carrier that can then be fed to shrimps (Immanuel et al. 2004). From this point of view, use of EOs as antimicrobials in aquaculture, directly in the water tank, is innovative.
Materials and Methods
EO extraction
C. fragrans leaf samples were harvested in Mariarano for B143 sample (15°41′S: 46°43′E, altitude: 7 m, 71.6% 1,8-cineole) and in Tsaramandroso for B8 sample (16°18′S; 47°02′E, altitude: 600 m, 95.8% linalool) and were extracted as described by Randrianarivelo et al. (2009).
EO and antibiotic assays
The in vivo experiments were carried out at the OSO organic shrimp hatchery in Madagascar. The Nauplii larvae were cultured for 18 days in duplicate, and the corresponding larval stages were, respectively, Nauplii (N1, N6), Zoe (Z1, Z1Z2, Z2, Z3, Z3M1), Mysis (M1, M2, M3, M3PL1), and postlarvae (PL1, PL2, PL3, PL4, PL5, PL6, PL7, PL8) from T0 till T18. Sea water was filtered and sterilized by a ultraviolet system. Initially, a water volume of 9 L was maintained, with daily addition of 0.75 L during the first 7 days. To renew the water, the water exchange rate used was 25% by the 11th day (stage PL1), 50% by the 12th day (stage PL2), 50% by the14th day (stage PL4), and 50% by the 16th day (stage PL6). The aquaria were inoculated with 400 larvae per liter. The culture conditions maintained were 29.2°C, 32% salinity, pH 7.8, and 4.5%–5.5% of dissolved oxygen levels. Feeding was carried out according to the official “AB Label” organic standards applied by the farm protocol. The treatment (EO or antibiotic [erythromycin]) (Bouchara) was applied once a day at 7 a.m., on days 3, 5, 7, 10, 12, and 14. The antibiotic concentration was 20 μg mL−1. The oil was added at a concentration of 734 μg mL−1. It was directly mixed with water. For the assays using Tween 80 (Fisher Labosi), the oil was mixed in a solution of 0.04% (v/v) Tween 80. In the control B, a solution of only emulsifier (Tween 80) was added at 0.04% (v/v) at the same frequency as the oil assays.
The following assays were performed in duplicate and during two campaigns in 2006 and 2007 as described by Randrianarivelo et al. (2010):
Control A: the larvae culture was not treated (no EOs, no antibiotic, no Tween 80).
Control B: the larvae culture was treated with Tween 80 (no EOs, no antibiotic).
B8 T: the larvae culture was treated with B8 EO with Tween 80.
B8: the larvae culture was treated with B8 EO without Tween 80.
B143 T: the larvae culture was treated with B143 EO with Tween 80.
B143: the larvae culture was treated with B143 EO without Tween 80.
E: the larvae culture was treated with the antibiotic erythromycin.
Bacterial concentration
The culture started from Nauplii stage (day 0) till PL8 (day 18). Every day during the 18 days of P. monodon culture, water (10 mL) from each aquarium was sampled. The water samples were diluted to 10−5 in sterile saline solution. A volume of 0.1 mL from each dilution was plated in duplicate onto marine agar (MA) for total heterotrophic aerobic bacteria (Difco) and onto thiosulfate citrate bile sucrose (TCBS) agar for vibrios (Difco) and incubated at 30°C for 48 h and 24 h, respectively.
Statistical analysis
Kruskal–Wallis test was used to determine the significant difference between the tested groups at a threshold of p < 0.05. The reproducibility of the data collected in 2006 and 2007 was validated by Kruskal–Wallis test with a threshold of p < 0.05. As the analysis of data collected in duplicate for 2006 and 2007 has demonstrated that all p-values were >0.05, the data presented in this article corresponded to both years 2006 and 2007 (four replicates).
Results
The bacterial concentration of the rearing water in both the EOs and antibiotic (E) tanks were significantly lower (p < 0.05) than in the control (Figs. 1 and 2). Further, there was no significant difference (p > 0.05) between the three treatments (B8, B143, and E). At day 18, the log10 reduction between the control and the three treatments were 0.32 log10 cfu mL−1 on MA and 0.93 log10 cfu mL−1 on TCBS.
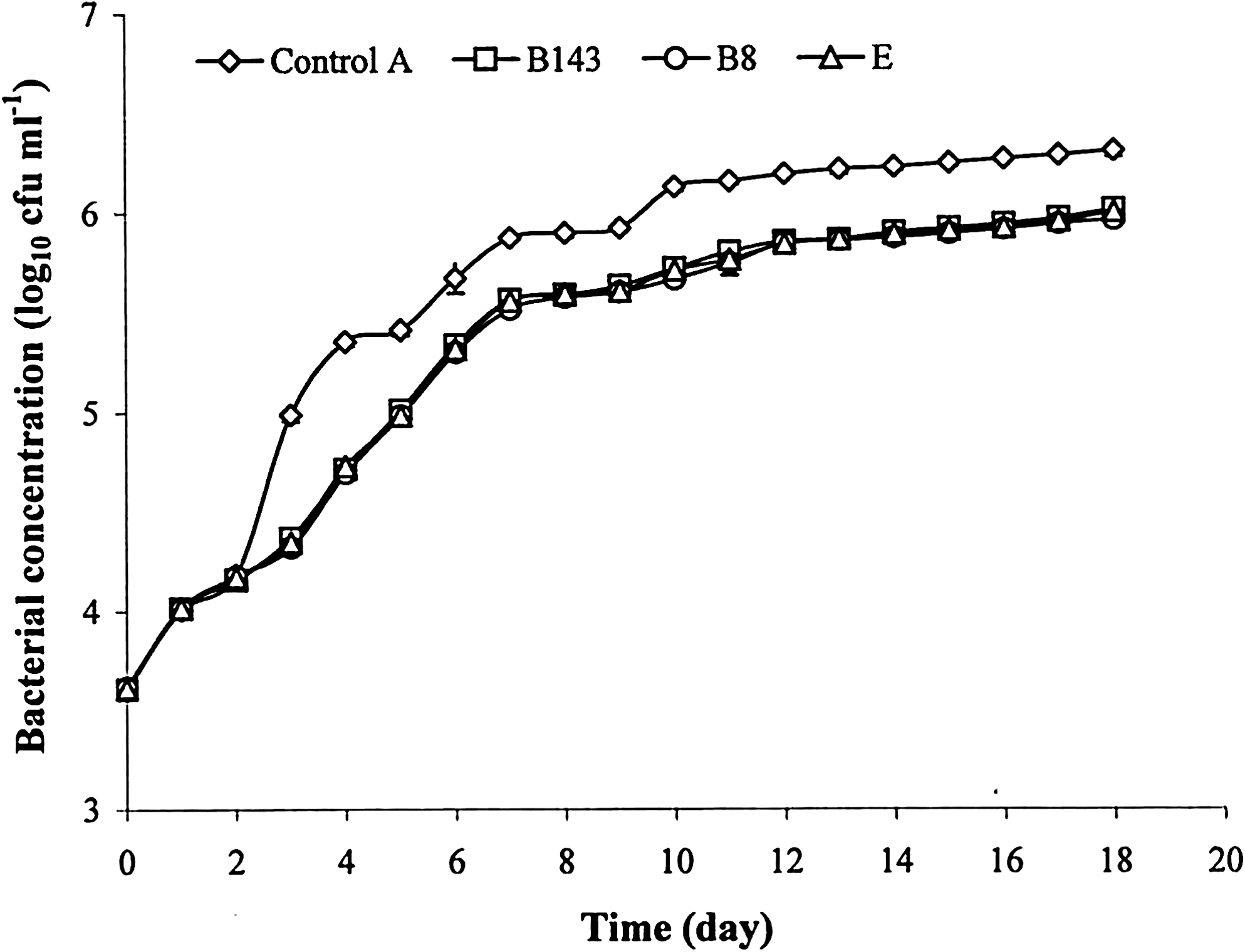
Evolution of total bacterial concentration of rearing water on MA (log10 cfu mL−1) during 18 days of culture of Penaeus monodon larvae for the control A (essential oil and antibiotic free) and for assays using the essential oils of Cinnamosma fragrans (B8 and B143) and erythromycin (E). Values correspond to the mean of four replicates (n = 4 ± SD). MA, marine agar; SD, standard deviation.
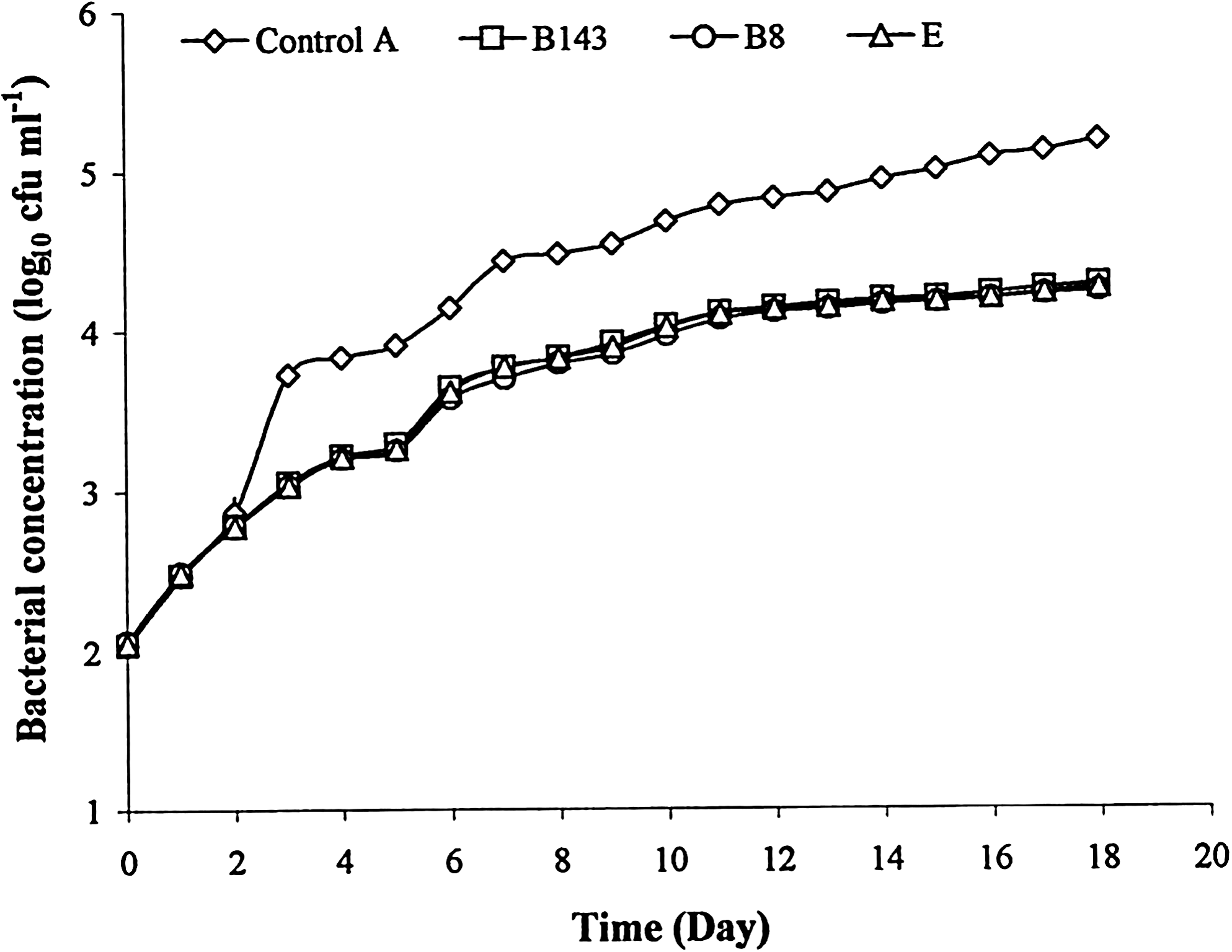
Evolution of Vibrio concentration of rearing water on TCBS (log10 cfu mL−1) during 18 days of culture of P. monodon larvae for the control A (essential oil and antibiotic free) and for assays using the essential oils of C. fragrans (B8 and B143) and erythromycin (E). Values correspond to the mean of four replicates (n = 4 ± SD). TCBS, thiosulfate citrate bile sucrose.
In all assays, use of an emulsifier (Tween 80) did not have a significant effect on the bacterial concentrations of the water (p > 0.05) (Fig. 3).

Evolution of Vibrio concentration of the rearing water of P. monodon larvae on TCBS (log10 cfu mL−1), for assays using essential oils of C. fragrans with Tween 80 (B8-T and B143-T; white symbols) and without Tween 80 (B8 and B143; black symbols). No significant difference was found between B8/B8-T and B143/B143-T (p > 0.05). Values correspond to the mean of four replicates (n = 4 ± SD).
Discussion
Total bacterial counts were consistently higher on the general heterotrophic medium (MA) than on the Vibrio-selective TCBS, indicating the presence of non–Vibrio-related microorganisms in the aquaculture system. During larval rearing, different microflora may have come into hatchery systems through live feeds such as algae and Artemia (Hameed and Balasubramanian 2000, Otta et al. 2001, Kennedy et al. 2006). Increasing Vibrio population in larvae and rearing tank water has been reported to reduce the survival rate of larvae and postlarvae (Regunathan and Wesley 2004). Previous results have shown that isolates on TCBS from water and larval samples included a variety of vibrios species, among which V. alginolyticus, V. parahaemolyticus, and V. vulnificus were dominant (Randrianarivelo et al. 2010). These species have been reported as pathogenic agents potentially transmitted to man from fish and fish products (ICMSF 2005).
In all assays, use of an emulsifier (Tween 80) did not have a significant effect on the bacterial concentration of the water (p > 0.05), which is in accordance with previous results obtained with P. monodon larvae (Randrianarivelo et al. 2010). Both EOs inhibited the bacterial growth in in vivo conditions of larval culture, like the antibiotic (Figs. 1 and 2). The log10 reduction was higher for Vibrio population than for total bacteria for the three treatments (EOs and antibiotic). This may be attributed to the antimicrobial effects of EOs as demonstrated by their low minimum inhibitory concentration against different Vibrio species (Randrianarivelo et al. 2009, 2010). Terpenes have been reported to cause disruption of bacterial membranes by leakage of intracellular constituents (Ultee et al. 1999, Di Pasqua et al. 2007). Because of their mode of action affecting several targets, no particular resistance or adaptation to EOs has been described so far in the literature (Bakkali et al. 2008).
Several studies have reported the potential to use plant extracts as an alternative to antibiotics in aquaculture and in organic farming in particular. Juveniles of Penaeus indicus fed with enriched diets (from terrestrial plants: Ricinus communis, Phyllanthus niruri, Leucus aspera, Manihot esculenta; and sea weeds: Ulva lactuca and Sargassum wightii) had better survival and growth, with inhibition of bacterial load of V. parahaemolyticus (Immanuel et al. 2004). The green algae Tetraselmis suecica, which has been reported to inhibit in vitro growth of pathogenic Vibrio spp., reduced Vibrio count in both rearing water and larval samples when it was used as feed in larval rearing tanks of the prawn Fenneropenaeus indicus (Indian white shrimp, Penaeidae) (Regunathan and Wesley 2004). Stevia plant extracts increased the immunity and disease resistance of P. monodon shrimp (Leaño et al. 2007). Similarly, the hot-water extracts from leaves and twigs of Camphor kanekirae have also shown better immunity of Litopenaeus vannamei shrimp and disease resistance to V. alginolyticus (Yeh et al. 2008). The protection of shrimps elicited by Dendrilla nigra, a marine sponge, against the most common shrimp pathogens such as luminescent Vibrio harveyi and V. alginolyticus could be due to its antibiotic effect rather than its influence on the host defense system (Selvin and Lipton 2004).
Use of EOs of C. fragrans in water tank have been found to be effective in reducing significantly the bacterial load of both heterotrophic bacteria and Vibrio spp. in in vivo conditions of larval culture. This might be a relevant option for producers to control the microbial growth in the rearing water, and Vibrio population in particular, using natural plant extracts rather than antibiotics. C. fragrans is a common species in the western forests of Madagascar. It is not indicated as a threatened species on the International Union for Conservation of Nature Red List (IUCN 2009). Moreover, EOs are extracted from leaves, which is not a destructive method. These reasons allow us to envisage a sustainable exploitation of this resource in Madagascar.
Footnotes
Disclosure Statement
No competing financial interests exist.