
Abstract
Ossabaw Island, Georgia, is the only reported endemic focus of Vesicular Stomatitis New Jersey Virus (VSNJV) in the United States. Based on recent negative serologic results of white-tailed deer and feral swine and the failure to isolate VSNJV from Lutzomyia shannoni, it appears that VSNJV is no longer present at this site. This apparent disappearance does not appear to be related to a change in L. shannoni habitat, specifically to the density of tree holes in the maritime and mixed hardwood forests. We believe that the disappearance of VSNJV from Ossabaw Island is directly related to a reduction in the feral swine population and a subsequent increase in the utilization of white-tailed deer by the known vector, L. shannoni.
Introduction
Based on previously published serologic data from white-tailed deer sampled on Ossabaw Island, the prevalence of VSNJV neutralizing antibodies decreased from 60% to 62% in 1956 (Karstad 1956) and 1983 (Stallknecht et al. 1986), respectively, to 38% and 23% in 1988 and 1989 (Fletcher et al. 1991), respectively. This observed decrease in antibody prevalence may reflect a sampling bias or an actual decrease in VSNJV transmission at this site. With regard to sampling bias, host age and location (forest type) have been shown to influence VSNJV antibody prevalence in white-tailed deer and feral swine on Ossabaw Island (Stallknecht et al. 1985, 1987, Stallknecht and Erickson 1986, Fletcher et al. 1991). A decrease in antibody prevalence also may be related to changing transmission patterns as a result of population changes in either the biological vector, L. shannoni, or the vertebrate amplifying hosts. With regard to the latter, there has been a notable decrease in the feral swine population related to increased hunter harvests during the 1990s and specific removal efforts; since 2001, >15,000 feral swine have been removed as part of island management practices (Georgia Department of Natural Resources [GADNR], personal communication).
The objectives of this study were to determine the current status of VSNJV antibody prevalence in the white-tailed deer and feral swine populations and to compare these results with historic data from both species. In an attempt to explain decreasing VSNJV antibody prevalence, results from a complete survey and qualitative analysis of the vector habitat (quantification and characterization of tree holes within the three forest types), an attempt to isolate VSNJV from the sand flies, and an analysis of blood-meal sources of L. shannoni are presented.
Materials and Methods
Serum collection
Blood samples were obtained from hunter-killed white-tailed deer and feral swine that were harvested during hunts operated by the GADNR as well as from feral swine removed during population control efforts. Age and location of the animal were obtained for each blood sample taken. For swine, age was classified as juvenile (<14 month of age) and adult (>14 month of age) based on tooth replacement. White-tailed deer were categorized into seven age classes (0.5, 1.5, 2.5, 3.5, 4.5, 5.5, and 6.5+ year) utilizing the methods of Severinghaus (1949). Blood was stored at 4°C; serum was separated by centrifugation and stored at −20°C before testing.
Serology
Serum samples from hunter-killed white-tailed deer and feral swine were tested for VSNJV neutralizing antibodies by virus neutralization as described (Jenney and Snyder 1981), using heat inactivated serum (56°C for 30 min.) and 103 median tissue culture doses (TCID50/mL) of an Ossabaw Island VSNJV isolate. To remain consistent with previously reported results, white-tailed deer samples were considered positive at antibody titers of 1:8 or greater (Stallknecht et al. 1986, Fletcher et al. 1991); and feral swine samples were considered positive at titers of 1:32 or greater (Stallknecht et al. 1985, 1986, 1987).
Sand fly collection
Phlebotomine sand flies, L. shannoni, were collected with funnel traps (Comer and Corn 1991) from 18 May to 7 September 2006. CDC (Center for Disease Control and Prevention) light traps augmented with dry ice (Corn et al. 1990) were added to the study on 8 July 2006. Traps were set during the late afternoon to early evening, usually between 4 and 8 PM, and retrieved by 8 AM the next morning. Light traps were hung from trees ∼1–2 m above the ground on the southern end of the island in the oak (Quercus spp.) forest.
On collection, funnel trap containers and nets from light traps were placed at 4°C for 15 min; and flies were removed and sorted on a chill table. Males and females were sorted into pools of up to 22 sand flies per pool, according to trap site, trap type, day of collection, and sex, Females that had obtained a blood meal were singly placed in a sample tube. Tubes were stored and transported in liquid nitrogen.
Virus isolation
Sand flies were transferred into tubes containing 0.1 mm glass beads and 500 μL of minimal essential media supplemented with 3% fetal bovine serum and 3% antibiotic/antimycotic (Sigma Chemical Co.). Samples were macerated using a Biospec Mini Bead Beater-8 at high speed, centrifuged at 9200× g (10,000 rpm) for10 min; and 100 μL of the supernatant was inoculated onto confluent African green monkey kidney Middle America Research Unit (Vero MARU) (ATCC) cells on 12-well plates. Cells were incubated at 37°C and observed for 72 h for cytopathic effect.
Sand fly blood-meal analysis
Blood-fed flies were placed in 300 μL Dulbecco's phosphate buffer solution (Sigma Chemical Co.) and macerated as described above. Total nucleic acids were extracted using the QiaAmp viral RNA mini kit (Qiagen Sciences) according to the manufacturer's suggested protocol. Attempts to amplify mammalian, avian, reptilian, and amphibian DNA using polymerase chain reaction (PCR) were conducted. Briefly, 2.5 μL of each sample was added to 0.5 mL thin-walled PCR tubes (USA Scientific Inc.) containing 47.5 μL of premix consisting of 5 μL 10× buffer, 1.5 mM MgCl2, 0.25 mM of each of the deoxyribonucleoside triphosphates, 0.15 units taq polymerase (Promega Corp.), 35.85 μL molecular grade biological water, and 0.5 μM each of the forward and reverse group-specific primers.
The group-specific primers were designed to target the 16s rDNA gene. The sequences of primers are as follows: mammalian f-5′ CCTGTTTACCAAAAACATCAC 3′, r-5′ AYTGTCGATAKGRACTCTWRARTAG 3′; avian f-5′ MMCAAGTATTGAAGGTGA 3′, r-5′ CTGATCCAACATCGAGGTCGT 3′; amphibian f-5′CTGTTTACCAAAAACATC 3′; reptile f-5′ CCRACTGTTTACCAAAAACATA 3′; ectotherm universal (used with reptile and amphibian forward primers) r-5′ ATCCAACATCGAGGTCGT 3′. These primers were designed using strategically placed base-pair mismatches, particularly among the three distal-most bases, gleaned from a multiple sequence alignment comprised of orthologs from representatives of several families within each animal class as well as orthologs from several potential insect vectors, mostly dipterans and hemipterans. Researchers at the Southeastern Cooperative Wildlife Disease Study and University of Tennessee, Knoxville, validated these primers in the laboratory against several locally common representatives of each class, particularly Aves and Mammalia.
Mammalian, avian, and reptilian reactions were incubated at 94°C for 2 min 30 s, followed by 39 cycles as follows: 94°C for 45 s, 55°C for 1 min, 72°C for 1 min 15 s, and concluded with an indefinite cooling step at 4°C. Amphibian reactions followed a modified protocol with an annealing temperature of 52°C instead of 55°C. PCR products were separated by electrophoresis, stained with ethidium bromide, and visualized under ultraviolet light. Samples were compared with known avian, mammalian, amphibian, and reptilian controls. Bands from amplified gene products were extracted and purified with the QIAquick gel extraction kit (Qiagen Sciences). DNA was sequenced in forward and reverse directions at the Molecular Genetics Instrumentation Facility, University of Georgia, with an ABI 3100 automated sequencer (Applied Biosystems, Perkins Elmer Corp.). Opposing sequences were assembled and reconciled using the Sequencher software package, version 4.1.4 (Gene Codes Corp.). A nucleotide-nucleotide BLAST (blastn) search was performed to determine the most similar sequences of the target genes published in GenBank (
Tree hole survey
In temperate regions, such as Ossabaw Island, GA, it is believed that sand flies utilize tree holes as habitat for overwintering and ovipositing and as a diurnal resting shelter (Comer and Brown 1993). During 1993, a tree hole survey was conducted on Ossabaw Island to determine which of the island's three major forest types (pine, mixed hardwood, and live oak/maritime) were suitable for supporting sand fly populations (Comer et al. 1993). In 2007, this survey was repeated as previously described for each of the forest types. Fixed radius circular plots, each measuring 1000 m2, were randomly assigned to each forest type; and the number of tree holes, as previously defined (Comer et al. 1993), were counted. Differences in prevalence of tree holes within forest types were tested by analysis of variance using SAS statistical software. Differences among the same forest type over time were also analyzed using SAS analysis of variance.
Results
VSNJV serology
From November 1990 to December 2008, serum samples were obtained from 1364 white-tailed deer. Antibody prevalence to VSNJV decreased to zero during this period (Fig. 1). Based on the slope of a linear regression model that included previously published VSNJV antibody prevalence estimates for deer on Ossabaw Island from 1981 to 1989 (Fig. 1) (Stallknecht and Erickson 1986, Fletcher et al. 1991), antibody prevalence decreased to ∼1.9% per year. The decrease in antibody prevalence was apparent across all age classes (Table 1).
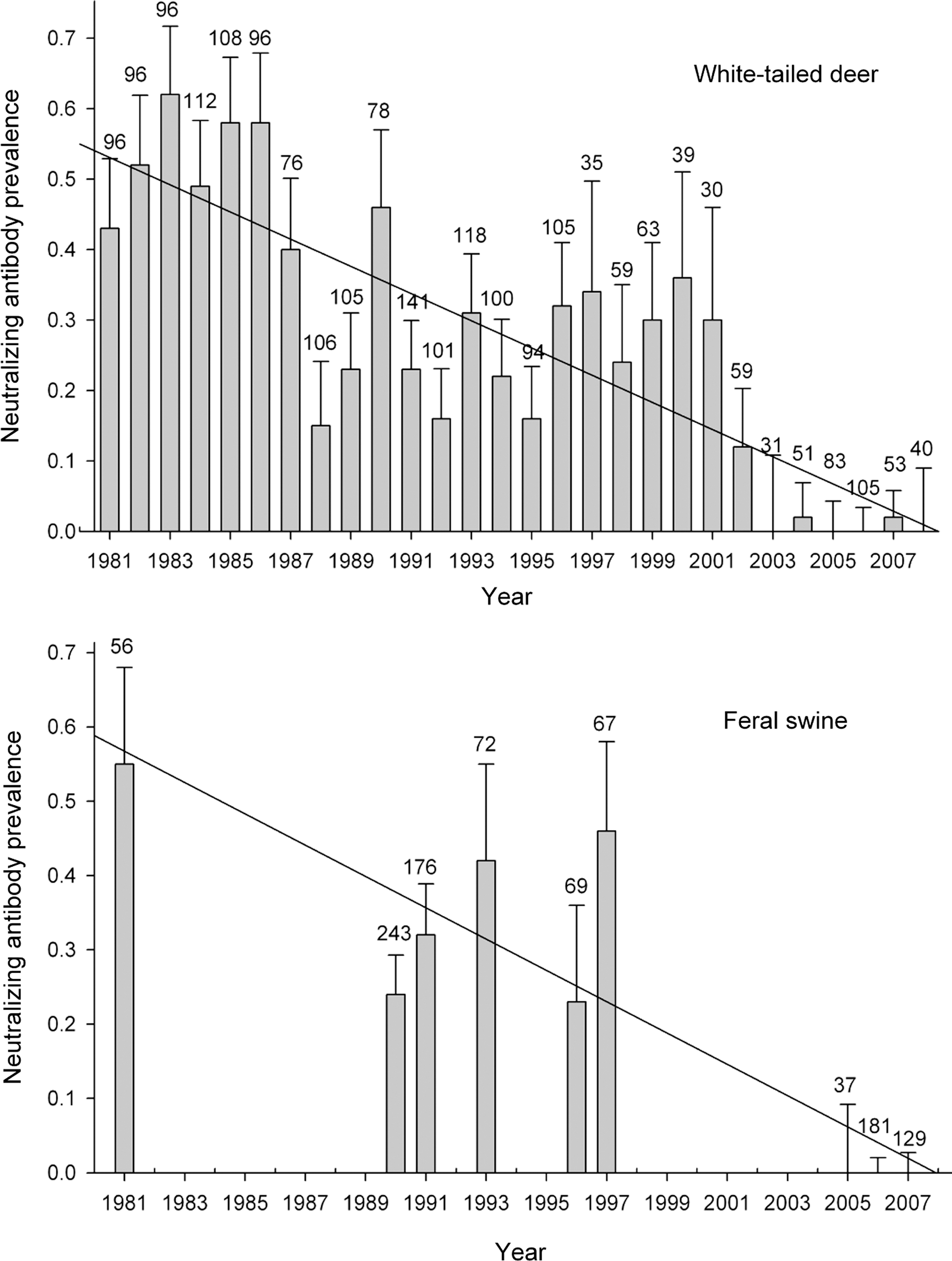
Annual antibody prevalence estimates for white-tailed deer and feral swine on Ossabaw Island Georgia from 1981 to 2008. Previously published data for white-tailed deer (1981–1984 and 1985–1989) and swine (1981, 1990, and 1991) are included. The solid line is a linear regression model based on the annual prevalence estimates; the number above each bar is the sample size.
Age data were not available for deer harvested from 1992 to 1995.
From November 2005 to February 2007, serum samples were obtained from 347 feral swine; an additional 208 serum samples were tested from hunter-killed feral swine harvested in 1993, 1996, and 1997. Antibody prevalence decreased to zero during this period (Fig. 1). Of the feral swine sampled from 2005 to 2007, 49% were juveniles and 51% were adults; ages of pigs sampled in 1993, 1996, and 1997 are unknown. Based on the linear regression model that included published prevalence estimates from 1981, 1990, and 1991, antibody prevalence decreased to ∼2.1% per year.
Virus isolation
Virus isolation was attempted from 324 sand flies pools and 56 blood-fed females (3167 sand flies); VSNJV was not isolated.
Blood-meal analysis
Of the 5139 sand flies, 56 blood-fed females were captured in the mixed hardwood areas; and 5 females were captured in the maritime forest. Of those captured in the mixed hardwood forest, 28 (55%) were obtained from a single tree over the course of the study. Using mammalian, avian, reptilian, and amphibian primers, DNA was amplified from 35 of these 56 sand flies and all products were mammalian in origin; only 2 species, white-tailed deer and feral swine, were identified as a blood-meal source. White-tailed deer accounted for 97% (34 of 35) of the species-identified blood-meals; the remaining blood-meal was of swine origin (1 of 35, 3%). The percentage of identified blood-meals from deer (97%) was higher than what was previously reported during 1994 (81%) (Comer et al. 1994). In contrast, the percentage of blood-meals from swine decreased from 16% in 1993 (Comer et al. 1994) to 3% in 2006. The majority (34/35) of the blood-meals that were identified came from sand flies captured in the mixed hardwood forest type.
Tree hole survey
In March 2007, each of the three forest types on the island was sampled to determine the mean number of tree holes per 1000 m2 plot. The numbers of plots sampled from each forest type are as follows: 35 plots in the live oak forest, 50 plots in the mixed hardwood forest, and 12 plots in the pine forest. The mean number of tree holes was calculated for each forest type and compared with data from 1993. Results are shown in Table 2. The mean number of tree holes detected was significantly different between each forest type (p = 0.0022) but was similar to previously reported data (Comer et al. 1993). For the live oak forest, the 2007 mean of 6.14 tree holes/plot was not significantly different (p = 0.1164) than the 7.93 mean reported in 1993. The mean number of tree holes in the mixed hardwood forest increased from 5.2 in 1993 to 7.58 in 2007 (p = 0.002). The mean number of tree holes/plot also increased in the pine forest (1.6 in 1993 to 3.08 in 2007), but this difference was not significant (p = 0.2370).
Previously published data (Comer et al. 1993).
SE, standard error of the mean.
Discussion
A decreasing prevalence of VSNJV serum-neutralizing antibodies in white-tailed deer was confirmed by the thorough sampling of all age classes of white-tailed deer and validated by detecting the same decreasing trend in antibody prevalence among feral swine. This decline in prevalence was not associated with a change in the age structure of the animals sampled; and for white-tailed deer, this was apparent within all age cohorts. For feral swine, sampled animals were distributed evenly between adults (51%) and juveniles (49%). Historically, prevalence estimates for adult swine on Ossabaw Island have approached 76% (Fletcher et al. 1985). Additionally, seroconversion in juvenile feral swine is known to occur in the summer months (Stallknecht et al. 1987) and can range from 12% to 60% in a given year. Antibody prevalence rates in this range would have been detected in our samples that were collected during fall. The failure to detect antibodies in white-tailed deer and feral swine from 2005 to 2008 also did not reflect a change in the location of sampled animals. With regard to potential location sampling bias, a significant relationship between forest type and antibody prevalence was reported for both white-tailed deer (Fletcher et al. 1991) and feral swine (Stallknecht et al. 1985, 1987, 1993) sampled on Ossabaw Island with the highest antibody prevalences associated with the maritime/live oak and mixed hardwood forests. Of the 274 negative white-tailed deer (2005–2008) and 339 negative feral swine (2005–2007) with known harvest locations, 220 (80%) deer and 276 (81%) feral swine were sampled from these forest types. Previous studies report an antibody prevalence of 44% (1985–1989) in deer (Fletcher et al. 1991) and a seroconversion rate of 43% in feral swine (1991) (Comer et al. 1993) from these same forest areas.
With the exception of a single deer, antibodies to VSNJV were not detected in deer (n = 281) or feral swine (n = 347) sampled during or after 2005. The single deer that tested antibody positive in 2007 was 6.5+ year old and was collected in the maritime forest. Due to the persistence of detectable VSNJV antibodies in deer and swine, it is difficult to determine the year when VSNJV disappeared from this ecosystem. However, based on results from individual age cohorts of white-tailed deer (excluding the single 2007 positive within the 6.5 year age class) (Table 3), it appears that transmission of VSNJV ended during the period 2001–2002. The failure to detect antibodies in the oldest age class of feral swine (>14 month old) during 2005 provides a similar estimate and, in this case, indicates that transmission did not occur after 2004. The failure to isolate virus from 5139 L. shannoni collected in 2006 supports our serologic data from white-tailed deer and feral swine; but due to a relatively low probability of virus isolation, these results alone do not provide conclusive evidence that VSNJV are no longer present on Ossabaw Island. Previous virus isolation rates are highly varied and range from 0.02% to 0.15% for nonblooded female sand flies (Corn et al. 1990, Comer et al. 1992) and from 0.008% to 0.017% for male sand flies (Comer et al. 1992).
Based on possibility that the deer was infected as a fawn (<6 month).
One 6.5+ year old deer tested antibody positive in 2007; this animal could have been older than 6.5 years, as this is the oldest age that can be determined by tooth eruption and wear.
The decline in antibody prevalence in white-tailed deer was not associated with an obvious change in the vector population. Sand flies are abundant on Ossabaw Island, and results from this study show that the habitat remains viable for the support of L. shannoni. In fact, when compared with the previous study (17 year ago), the only statistically significant change is an increase in the number of tree holes available as sand fly shelters (Comer et al. 1993).
There has been a clear change in host feeding behavior as was evidenced by blood-meal analysis of blood-fed female sand flies. White-tailed deer are still the primary hosts for sand flies (97%), but there has been an obvious reduction in utilization of feral swine for blood-meals (16% in 1993 to 3% in 2006) (Comer et al. 1994). We believe that this change is related to the feral swine population declines related to removal efforts established by DNR in the late 1990s. Unfortunately, accurate population estimates that would be needed to statistically demonstrate the effects of removal efforts are not available.
Feral swine have the potential to amplify VSNJV through direct or indirect contact transmission and by providing a suitable host for vector-borne transmission. They also are the only wildlife species in which naturally occurring VSNJV infection has been confirmed by virus isolation (Stallknecht et al. 1987, 1993). In experimental studies, efficient contact transmission of VSNJV between swine has been demonstrated but appears to be dependent on clinical disease (vesicular lesions) (Stallknecht et al. 2001, 2004). Mechanical transmission of VSNJV by insects to domestic swine has been recently demonstrated in an experimental setting (Smith et al. 2009). Finally, biological transmission by insect vectors feeding on swine has been demonstrated (Mead et al. 2004a); in this case, virus can be obtained by black flies by feeding on lesion-rich material or skin in the absence of a detectable viremia (Mead et al. 2004b). Virus can be further amplified within the insect vector population through co-feeding between infected and uninfected insects feeding on noninfected hosts (Mead et al. 2004b) and through transovarial transmission, which has been demonstrated with L. shannoni and VSNJV (Comer et al. 1990). In addition to providing a viable host for VSNJV amplification, feral swine were extremely abundant on Ossabaw Island; and with a high reproductive rate, a significant number of susceptible animals were annually added to the population. The extent of VSNJV annual transmission to these susceptible juveniles is evident by seroconversion rates from 1982 to 1986, which ranged from 12% to 60% (Stallknecht et al. 1985, 1987).
Based on our past and current field data from Ossabaw Island and the results of experimental studies in swine, we believe that VSNJV at this site represented a remnant of the VSNJV epidemics that affected livestock (including domestic pigs) in the southeastern United States during the 1950s and 1960s (Hanson 1952, Jenney 1967). VSNJV was first isolated in the southeastern United States from swine in 1943, when an outbreak occurred at a hog cholera vaccine plant (Shahan et al. 1946, Karstad and Hanson 1957). Genetically, the Ossabaw Island VSNJV strain is most closely related to the original isolates associated with this outbreak (Bilsel et al. 1990, Vernon et al. 1990, Rainwater-Lovett et al. 2007). Two factors could explain this prolonged persistence. The first relates to the presence of L. shannoni, a native sand fly species that is a competent vector for VSNJV. Although transovarial transmission of VSNJV in this species has been demonstrated (Comer et al. 1990), the rate of transmission is relatively low; and without periodic amplification via a vertebrate host, VSNJV would probably not have persisted. The second relates to the presence of a suitable amplifying host; and we believe that the high-density feral swine population that was present on Ossabaw Island fulfilled this need. This exotic species is highly susceptible to VSNJV, develops clinical disease (which is important for amplification via contact transmission), is utilized by L. shannoni as a host, and can provide a source of virus for biting arthropods as demonstrated with black flies (Mead et al. 2004b). The breakdown in the extent of contact between vector and amplifying hosts as a result of removal efforts directed at that host is apparent in the results of the blood-meal analysis which demonstrated reduced utilization of swine by feeding L. shannoni.
To our knowledge, this is the only record of the disappearance of an endemic VSNJV focus. If this system was dependent on feral swine, it would imply that the now potentially extinct VSNJV focus occurred as a result of anthropogenic activity, specifically the introduction and establishment of two invasive species, feral swine and VSNJV, into a very unique and pristine ecosystem (old-growth maritime forest) that supported a viable L. shannoni population. Together, we believe that these components provided a focal environment in which VSNJV was successfully maintained from an original outbreak that started in the late 1940s.
Footnotes
Acknowledgments
The authors thank the Ossabaw Island Foundation and the GADNR. Numerous sand fly traps and samples and technical assistance were provided by Ed Rowton. Additionally, the authors thank Ben Wilcox, Luis Rodriguez, Steve Pauszek, Kaitlin Rainwater-Lovett, and others who helped collect samples. This research was supported by USDA, APHIS, Veterinary Services Cooperative Agreement 08-9613-0032-CA. Additional support was through the United States Department of Agriculture (CRIS-1940-32000-04000D) and through sponsorship of SCWDS by the fish and wildlife agencies of Alabama, Arkansas, Florida, Georgia, Kansas, Kentucky, Louisiana, Maryland, Mississippi, Missouri, North Carolina, Ohio, Puerto Rico, South Carolina, Tennessee, Virginia, and West Virginia.
Disclosure Statement
No competing financial interests exist.