
Abstract
Mainly through vector transmission, domestic cats and dogs are infected by several Bartonella spp. and represent a large reservoir for human infections. This study investigated the relationship of prevalences of Bartonella infection in shelter dogs and cats and various ectoparasite species infesting them (fleas, ticks, and lice). Moreover, relationships between Bartonella infection and animal gender and age and presence of ectoparasites were analyzed. Blood samples were collected from 120 dogs and 103 cats. There were 386 ticks and 36 fleas harvested on these dogs, and 141 fleas, 4 ticks, and 2 lice harvested on these cats. Isolation/detection of Bartonella sp. was performed by culture, polymerase chain reaction (PCR), and partial sequencing. Bartonella was isolated from 21 (20.4%) cats and detected by PCR from 20 (19.4%) cats, 2 (1.7%) dogs, 55 (39%) fleas collected from cats, 28 (10%) ticks DNA samples, and 1 (2.8%) flea collected from dogs. When combining culture and PCR data, 27 cats and 55 fleas collected on cats were positive for Bartonella henselae or Bartonella clarridgeiae, but none were coinfected. Approximately half of the B. henselae isolates from 21 cats were B. henselae type I. Moreover, B. henselae, Bartonella phoceensis, Bartonella queenslandensis, Bartonella rattimassiliensis, Bartonella elizabethae DNA was detected in ticks collected from dogs and one flea was B. clarridgeiae PCR positive. This is the first report of such a wide variety of Bartonella spp. detected in Rhipicephalus sanguineus. Further studies are required to understand the relative importance of these ectoparasites to transmit Bartonella spp. in dogs and cats.
Introduction
B. henselae and B. clarridgeiae seroprevalence or bacteremia have been reported in domestic cats in Asian countries, including Japan, Taiwan, Indonesia, Philippines, Korea, Singapore, and Thailand (reviewed in Saisongkorh et al. 2009). Stray and shelter cats usually have a higher Bartonella bacteremia prevalence than pet cats (Chomel et al. 1995, Heller et al. 1997, Boulouis et al. 2005). Ectoparasites, such as fleas, ticks, and mites, are commonly found on impounded cats and dogs and can harbor Bartonella spp. (Billeter et al. 2008). Lice, fleas, and sandflies have been reported as vectors of five Bartonella spp.—B. quintana, B. bacilliformis, B. henselae, B. grahamii, and B. taylorii (reviewed in Billeter et al. 2008). Various mites (Kim et al. 2005), keds, and biting flies (reviewed in Chomel et al. 2009a), and possibly ticks (reviewed in Billeter et al. 2008), are now considered as potential or suspected vectors for Bartonella transmission.
The objectives of this study were to investigate the prevalence of Bartonella infection in impounded dogs and cats and in the ectoparasites infesting them and to determine if animal gender, animal age, and presence of ectoparasites are associated with the infection.
Materials and Methods
Sample collection and blood culture
Between August 2008 and February 2009, blood samples and ectoparasites were collected from 120 impounded dogs from two animal shelters and 103 impounded cats from one animal shelter in central Taiwan. Within 7 days after the animals were sent to the shelters, ∼2 mL of whole blood from each dog and cat were taken from the jugular or saphenous veins into plastic EDTA tubes and frozen at −80°C before tested. Ectoparasites on these animals were collected with tweezers in 70% ethanol tubes and identified by morphological keys (Teng and Jiang 1991, Walker et al. 2000).
Direct culture and pre-enrichment culture were used to isolate Bartonella spp. from each dog blood (DB) and cat blood (CB) sample. Direct culture was performed as previously described (Chang et al. 2006). For pre-enrichment culture, blood samples were incubated in liquid Bartonella–Alphaproteobacteria growth medium (Maggi et al. 2005). Following a 7-day incubation period, 100 μL of the sample from the liquid medium were then subcultured on chocolate agar plates and examined weekly for at least 4 weeks.
DNA extraction, polymerase chain reaction, and sequencing
Two Bartonella suspect isolates obtained from each chocolate agar plate were randomly selected for DNA extraction as previously described (Maillard et al. 2004). DNA of each whole blood sample was extracted using the DNeasy tissue kit (Qiagen) following the manufacturer's instructions. When the number of ticks for each stage, fleas, or lice on an animal was five or less, every tick, flea, or louse on this animal was collected and subjected to DNA extraction. If the number of adult ticks, nymph ticks, and fleas on sampled animals was more than five, five adult ticks, five nymph ticks, and five fleas attached to that animal were randomly selected for molecular detection. Each adult tick was cut into equal halves. Half of each tick was transferred to a clean microtube for DNA extraction. Whole tick nymphs, pooled tick larvae (10 or less), whole flea, and pooled lice (10 or less) were used for DNA extraction. When >50 tick larvae were collected for an animal, only five pools of 10 larvae were selected for molecular detection. DNA extraction was performed as described earlier.
Polymerase chain reaction (PCR) of each individual DNA sample from isolated colonies, animal blood, and ectoparasites was performed with Bartonella-specific primers of the citrate synthase (gltA) gene (Norman et al. 1995), modified as reported previously (Lin et al. 2008). Previously, B. henselae, B. clarridgeiae, Bartonella rattimassiliensis, B. tribocorum, B. grahamii, B. elizabethae, Bartonella phoceensis, and B. rochalimae-like strains have been cultivated in our laboratory. However, we only used B. henselae as the positive control for checking PCR results. The cultivation room and PCR room are physically separated ones with separate sets of equipment to prevent possible cross-contamination in our laboratory. To prevent molecular contamination, pre-PCR activities (PCR preparation, DNA extraction, PCR amplification) and post-PCR activities (electrophoresis and gel expression) were also performed in separate rooms with separate sets of equipment. B. henselae Houston-1 (ATCC 49882) and double-distilled water were used as positive and negative controls, respectively.
The gltA PCR-positive fragments of DB and dog ticks (DT) were cloned using pGEM®-T vector system (Promega). Sequencing was performed with ABI 3730 XL DNA Analyzer (Mission Biotech Co. Ltd.).
16S rRNA typing of B. henselae
Genotyping of B. henselae was performed by amplifying the 16S rRNA gene (Bergmans et al. 1996), modified as reported previously (Chang et al. 2006). The strains Houston-1 and U-4 (a type II feline strain isolated at the University of California, Davis) were used as the reference strains of B. henselae type I and II, respectively.
Phylogenetic analysis and additional experiments
DNA samples having the same sequence information were assigned a group number. The highest similarity value of each group was determined by comparing with fragments deposited in GenBank. By MEGA version 4, phylogenetic analysis was performed using neighbor-joining method by Kimura 2-parameter model, and bootstrap support was calculated using 1000 bootstrap data.
When sequences have a similarity value lower than 95% with known Bartonella sp., two sets of primers for the 16S-23S ITS region and three sets of primers for the rpoB, ftsZ and 16S rDNA genes, as previously described, were used to analyze the DNA extracts (Heller et al. 1997, Renesto et al. 2001, Zeaiter et al. 2002, Maillard et al. 2004).
Statistical analysis
By StatXact version 8.0, chi-square test was used for data analysis. Two-tailed Fisher's exact test was used when expected numbers of observations were less than five. A p-value of <0.05 was considered to be statistically significant.
Results
Sample collection and blood culture
Of the 120 dogs, 84 (70%) were adults (≥1 year old), including 43 females and 41 males, and 36 (30%) were juveniles (<1 year old), including 19 females and 17 males. Of the 103 cats, 72 (70%) were adults (≥1 year old), including 31 females and 41 males, and 31 (30%) were juveniles (<1 year old), including 12 females and 19 males.
There was no isolation from DB samples. For cats, the blood of 21 (20.4%) animals yielded Bartonella suspect colonies by direct culture, but no isolation was obtained by pre-enrichment culture.
Ectoparasites were collected on 67 (55.8%) of the 120 dogs. Of these 67 dogs, 21 (31.3%) were adult females, 26 (38.8%) adult males, 10 (14.9%) juvenile females, and 10 (14.9%) juvenile males. For morphological identification and DNA extraction, 386 ticks (187 adults, 73 nymphs, and 126 larvae) and 36 adult fleas were selected. Of these 386 ticks, 151 (80.7%) adults were identified as Rhipicephalus sanguineus, 36 (19.3%) adults were Rhipicephalus haemaphysaloides, and all nymphs and larvae were identified at the genus level as Rhipicephalus spp. The 36 fleas were all Ctenocephalides felis felis.
Ectoparasites were collected on 46 (44.7%) of the 103 cats. Of these 46 cats, 8 (17.4%) were adult females, 20 (43.5%) were adult males, 5 (10.9%) were juvenile females, and 13 (28.3%) were juvenile males. A total of 141 adult fleas, 4 ticks (3 adults and 1 larva), and 2 nymph lice were selected for morphological identification and DNA extraction. All the fleas were C. felis felis. The four ticks were identified as R. sanguineus (three adults) or Rhipicephalus spp. (one larva). The two lice were Felicola subrostratus.
DNA extraction, PCR, and sequencing
Dogs
DNA was individually extracted from all 120 DB samples, 187 adult ticks, 73 nymph ticks, 21 pooled (range: 1–10 larvae) larva tick samples, and 36 fleas. Bartonella was detected in 2 (1.7%) DB samples, 12 (6.4%) adult ticks, including 11 R. sanguineus and 1 R. haemaphysaloides (sample DT 38-2), 13 (17.8%) nymph ticks, 3 (14.3%) pooled larva tick samples, and 1 (2.8%) flea (Table 1).
Number of pooled DNA samples.
PCR, polymerase chain reaction.
Cats
DNA was extracted from 40 colonies from 21 CB (2 colonies for 19 cats and 1 colony for 2 cats), all 103 CB samples, 141 fleas, 3 adult ticks, 1 larva tick, and 1 pooled (2 lice) louse sample. Bartonella was detected in 40 (100%) isolates, 20 (19.4%) CB samples, and 55 (39%) fleas, but not in the tick and louse DNA samples (Table 1).
16S rRNA typing of B. henselae
Of the 40 isolates, no B. henselae coinfection was detected in 21 cats, with 10 (47.6%) and 11 (52.4%) cats, respectively, infected with B. henselae type I and type II.
Sequencing and phylogenetic analyses
Dogs
Sequences could be properly interpreted for the gltA gene from each PCR-positive sample, including 2 DB DNA samples, 12 adult ticks, 13 nymph ticks, 3 pooled larva tick samples, and 1 flea. Bartonella strains used as references for comparison of the gltA gene are shown in Figure 1. Based on the gltA gene, sequences of 2 DB samples had a 100% similarity to Bartonella sp. SL-1/EU368000. For the 28 DT DNA samples, 30 sequences were obtained after cloning (c1 and c2). The highest similarity value of each sequence is shown in Table 2. For these tick samples, Bartonella sp. SL-1, B. henselae, B. elizabethae, and B. phoceensis were detected from adults, B. henselae, B. phoceensis, Bartonella queenslandensis, B. rattimassiliensis, and B. elizabethae were detected from nymphs, and B. henselae and B. rattimassiliensis were detected from larvae. The DNA sample of the flea collected from a dog had a 98% similarity to B. clarridgeiae (Table 2).
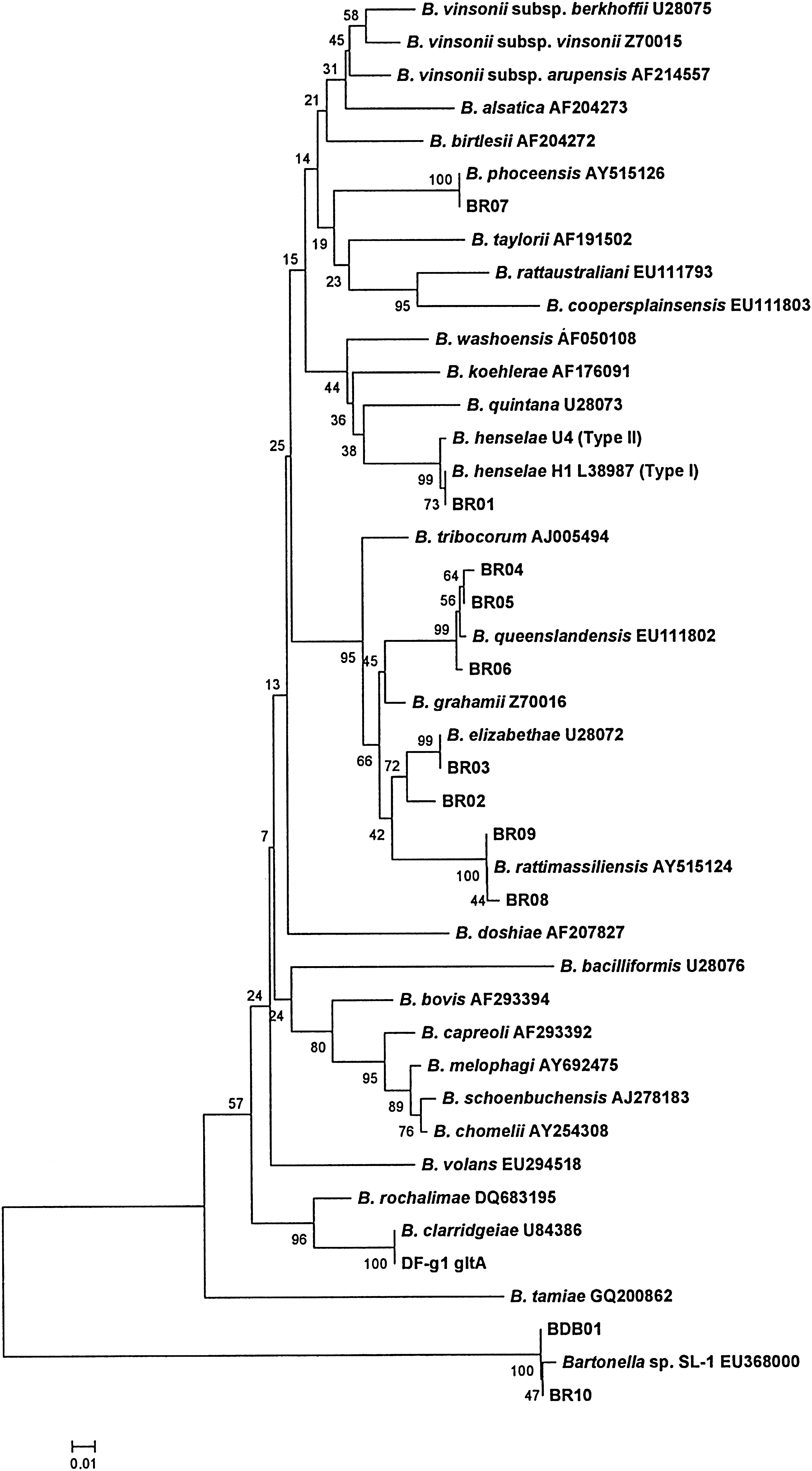
The phylogenetic tree was based on 224 base pairs (bp) of gltA gene for Bartonella strains using MEGA version 4. The tree was obtained using the neighbor-joining method by Kimura-2 parameter model, and bootstrap support was calculated by using 1000 bootstrap data.
No other similar sequences except Bartonella sp. SL-1 (EU368000) were found when compared with the fragments in GenBank based on sequenced gltA gene.
The phylogenetic tree derived from gltA dataset also showed the sequences to be closely related to B. henselae, B. phoceensis, B. queenslandensis, B. rattimassiliensis, B. elizabethae, and B. clarridgeiae and an independent clade including Bartonella sp. SL-1 (Fig. 1).
Cats
For the gltA gene, sequences could be interpreted for 40 (100%) CB isolate DNA samples, but only 7 (35%) of the 20 PCR-positive samples for the CB DNA extracts and 48 (87.3%) of the 55 PCR-positive samples for the flea DNA samples. Sequences of all 40 CB isolate DNA samples, 6 CB DNA samples, and 17 cat fleas DNA samples had a 100% similarity to B. henselae. One CB DNA sample and 31 cat flea DNA samples had a 100% similarity to B. clarridgeiae.
The phylogenetic tree derived from gltA dataset also supported these sequences to be closely related to B. henselae and B. clarridgeiae (Fig. 2). The gltA sequences of CB samples had the same B. henselae genotype classification as the PCR results using 16S rRNA genotyping.

The phylogenetic tree based on 230 base pairs (bp) of gltA for Bartonella strains drawn using MEGA version 4. The tree was obtained using the neighbor-joining method by Kimura-2 parameter model, and bootstrap support was calculated by using 1000 bootstrap data.
Statistical analysis
Cat blood
Sex and age were not identified as specific risk factors for bacteremia in cats by either culture or PCR results [male: 30% (18/60) vs. female: 21% (9/43); p = 0.37; and juvenile: 38.7% (12/31) vs. adult: 20.8% (15/72); p = 0.086].
Flea
Fleas collected from cats were more likely to harbor Bartonella spp. than those collected from dogs [cat: 39% (55/141) vs. dog: 2.8% (1/36); p < 0.0001]. The percentage of Bartonella PCR-positive fleas collected from female cats (34.4%, 11/32) was not statistically different from that from male cats (40.4%, 44/109; p = 0.68). Fleas collected from juvenile cats (39%, 33/65) were more likely to harbor Bartonella spp. than those from adult cats [28.9% (22/76); p = 0.0097]. The percentage of Bartonella PCR-positive male fleas was not statistically different from that in female fleas [male: 29.7% (44/104) vs. female: 42.3% (11/37); p = 0.24].
Tick
Ticks collected from male dogs were more likely to harbor Bartonella spp. than those from female dogs [male: 14.4% (24/167) vs. female: 3.6% (4/114); p = 0.0038]. However, this significant crude association between Bartonella infection status of ticks collected from dogs and dog gender was confounded by the life stage of ticks. After control for the life stage of ticks, dog gender was no longer significantly associated with Bartonella infection of ticks. The percentage of positive ticks collected from adult dogs was not statistically different from that from juvenile dogs [adult: 9.4% (17/181) vs. juvenile: 11% (11/100); p = 0.68]. The percentage of Bartonella PCR-positive nymph ticks was significantly higher than adult ticks [nymph: 17.8% (13/73), adult: 6.4% (12/187); p = 0.0088], but not for larva ticks [14.3% (3/21); p = 0.37]. The percentage of male ticks that were Bartonella PCR positive was not statistically different from that in female ticks [male: 7.3% (7/96) vs. female: 5.5% (5/91); p = 0.77].
Cat and flea
Bacteremic cats were more likely than nonbacteremic cats to be infested with fleas [positive cat: 65.4% (17/26) vs. negative cat: 37.7% (29/77); p = 0.022; odds ratio = 3.1; 95% confidence interval = 1.1, 9.0]. Fleas collected from Bartonella-positive cats were more likely to be Bartonella PCR positive than the fleas collected from Bartonella-negative cats [positive cat: 63.3% (38/60) vs. negative cat: 21% (17/81); p < 0.0001; odds ratio = 6.5; 95% confidence interval = 2.89, 14.8].
Additional experiments
Two DB DNA samples of group BDB01 and four DT DNA samples of group BR10 were further analyzed with additional Bartonella genes. Only one DT DNA sample, DT 3-1, was PCR positive using primers aiming at the ITS pair 1 region (annealing at 60°C), and the obtained sequence had a 95% similarity to Mesorhizobium spp.
Discussion
Similar to a study conducted in northern Taiwan (Chang et al. 2006), our results are also within the prevalence range of Bartonella infection in cats reported from other Asian countries (Boulouis et al. 2005, Chang et al. 2006, Inoue et al. 2009). However, a recent study conducted in Korea reported a much higher Bartonella prevalence in both sheltered (41.8%) and pet cats (33.3%) (Kim et al. 2009). B. henselae prevalence is usually higher than B. clarridgeiae prevalence in cat populations (Boulouis et al. 2005) and was also confirmed in Taiwan both by Chang et al. (2006) and in our study.
As reported by Gurfield et al. (2001) in France, no gender difference was identified for Bartonella prevalence in the present study. However, because of the inconsistent results previously obtained (Zangwill et al. 1993, Sander et al. 1997, Gurfield et al. 2001, Inoue et al. 2009), future studies on larger cat populations are necessary to demonstrate a possible correlation between Bartonella infection and cat gender.
Bacteremia prevalence is usually higher in young cats, especially impounded cats, than in adults, as cats are more likely to acquire B. henselae infection during the first year of life (Chomel et al. 1995). Therefore, it could explain why more fleas on juvenile cats were more likely to be PCR positive for Bartonella than those on adults in this study. Transmission of infection among cats requires the presence of fleas, and bacteremic cats are more likely than nonbacteremic cats to be infested with fleas (Chomel et al. 1995). Similar results were detected in our study, even though there was no significant association between cat age and bacteremia status in animals.
B. henselae isolates from cats in many Asian countries mainly belong to genotype I, including Japan, Taiwan, Philippines, and Thailand, with the highest prevalence of coinfection with B. henselae type I and II (9.2%) in the world detected in Taiwan (Chomel et al. 1999, Boulouis et al. 2005, Chang et al. 2006, Inoue et al. 2009). In the present study, almost half of B. henselae isolates were B. henselae type II and none of the cats were coinfected. These different observations may relate to the origin of the cat populations tested.
Cat fleas (C. felis) are the main vector of B. henselae transmission and B. henselae detection from cat fleas have been reported in many countries (reviewed in Billeter et al. 2008). B. clarridgeiae has been also detected in cat fleas (C. felis) infesting cats or dogs in France, Thailand, New Zealand, Hungary, Spain, and Germany (reviewed in Billeter et al. 2008, Just et al. 2008). In our study, most of the B. henselae PCR-positive fleas were collected from B. henselae PCR-positive cats. However, many fleas collected from B. henselae-positive cats were PCR positive for B. henselae, B. clarridgeiae, or both. The high prevalence of fleas PCR positive for B. clarridgeiae and the very low prevalence of this bacterium in cats raise the hypothesis that B. clarridgeiae may still be highly adapted to its flea host and not completely to its vertebrate host.
In further molecular detection, a segment close to Mesorhizobium sp. was detected in one DT using ITS pair 1 primers. However, primers designed to amplify the Bartonella ITS region could result in nonspecific amplification of Mesorhizobium sp., which may lead to a false-positive diagnostic test result (Maggi and Breitschwerdt 2005). The Bartonella sp. SL-1 gltA sequence, along with a segment of ITS region, was detected in sucking louse, Neohaematopinus sciuri, collected from a dead eastern grey squirrel (Sciurus carolinensis) in Greenville Zoo of South Carolina (Nelder et al. 2009). This sequence of ITS region has been previously reported in one unidentified tick on a sheep in Peru and the bacterium was named as genotype Bartonella sp. T7498 (Parola et al. 2002). Because Bartonella sp. SL-1 is far away from other Bartonella spp. and no other fragment of Bartonella could be detected in our study, further studies will be required to determine if this strain truly belongs to the Bartonella genus.
R. sanguineus and R. haemaphysaloides, which are both three-host ticks, are the main tick species found on dogs in Taiwan (Tsai 2002). Although domestic dogs are the preferred hosts for all stages of R. sanguineus, they can also parasitize other animals, such as rodents, birds, reptiles, herbivores, carnivores, and humans (Marquardt et al. 2000). Similarly, R. haemaphysaloides ticks have been collected on other animals, and rodents are the primary hosts in its larva and nymph stages (Walker et al. 2000). In this study, we confirmed the presence of several Bartonella spp. in R. sanguineus ticks in Taiwan, including B. henselae, B. elizabethae, B. phoceensis, B. rattimassiliensis, and B. queenslandensis. The unknown strain close to Bartonella sp. SL-1 was harbored by both R. sanguineus and R. haemaphysaloides. This is the first report of such a wide variety of Bartonella spp. carried by R. sanguineus. These findings raised important issues for possible Bartonella transmission within ticks, between ticks and dogs, and possibly to humans. However, ticks may not be efficient vectors for several Bartonella spp., which are not adapted to its dog host. The lower Bartonella prevalence in adult ticks compared with nymphs and larvae in this study may also indicate the possibility of very limited transstadial transmission in ticks. In this study, two adult ticks harbored simultaneously B. henselae and B. elizabethae, and B. elizabethae and B. phoceensis, respectively. Because three-host ticks change hosts between different life stages, coinfection with different pathogens may result from blood feeding on different hosts.
In this study, the proportion of ectoparasites containing host blood was not recorded and analyzed in this study. Therefore, the association of presence of host blood and detection of Bartonella DNA in ectoparasites could not be evaluated. It is of major importance to evaluate that these arthropods could play the biological role for Bartonella transmission in future studies.
In this study, for Bartonella identification, additional PCRs (ITS region, rpoB, ftsZ and 16S rDNA genes) were performed only for those samples with PCR positivity for the gltA gene and their gltA sequences with a similarity value lower than 95% comparing with the known Bartonella sp. There were only six samples fitting this criteria for further rpoB and ftsZ PCR testing. Unfortunately, the results of rpoB and ftsZ PCRs were all negative in these six samples tested. It could be of concern that false-positive results occurred by PCR of the gltA gene. However, we considered that, under the well-controlled laboratory conditions, obtaining false-positive results were very unlikely, although it cannot be completely prevented. As a matter of fact, for those positive samples, they were checked again by PCR. Only samples showing positive results consecutively were determined to be positive ones. It was considered that the main reason for those samples that are PCR positive but not yielding interpretable sequences could be due to very low levels of DNA for sequencing analysis. This phenomenon is especially observed when using PCR products directly from animal blood or vectors.
In this study, some Bartonella spp. reported in small mammals in Taiwan (Lin et al. 2008, Hsieh et al. 2009) were detected from DT at different stages. These findings raise the possibility that dogs may be infected by Bartonella spp. through ticks that became Bartonella infected when feeding on previous small mammal hosts. Therefore, it will be important to determine the role of ticks in the transmission of these pathogens between animals and possibly to humans.
Footnotes
Acknowledgments
The authors thank the Taichung City Animal Protection and Health Inspection Center and Hsin Chu County Animal Disease Control Center in Taiwan for allowing sample collection. This study was supported by a grant (NSC95-2313-B-005-028-MY2) from the National Science Council, Executive Yuan, Taiwan.
Disclosure Statement
No competing financial interests exist.