
Abstract
A 1-year study of the ecological cycle of Francisella tularensis was performed in an enzootic area during an inter-epizootic period. The study was based on multiple sampling of all major constituents of the disease cycle. Seroprevalence of tularemia in the European brown hare (Lepus europaeus) population was 5.1% (10/197) with low antibody titers (1/10 and 1/20), and F. tularensis ssp. holarctica was isolated from four hares. F. tularensis was not detected in the 38 common voles (Microtus arvalis), 110 yellow-necked mice (Apodemus flavicollis), or 15 stripped field mice (Apodemus agrarius) trapped during the study, or the by-catch of 8 Eurasian pygmy shrews (Sorex minutus) or 6 common shrews (Sorex araneus). A total of 1106 Ixodes ricinus and 476 Haemaphysalis concinna ticks were collected from vegetation, and 404 I. ricinus, 28 H. concinna ticks, and 15 Ctenophtalmus assimilis and 10 Nosopsyllus fasciatus fleas were combed off small mammals. One H. concinna female and one nymph collected from the vegetation was found infected with F. tularensis ssp. holarctica by TaqMan polymerase chain reaction, thus resulting a 0.42% (2/476) prevalence. F. tularensis–specific DNA was not detected in environmental water samples, and the examined 100 sheep, 50 cows, and 50 buffalos grazed at the study area were all seronegative. During inter-epizootic periods, F. tularensis ssp. holarctica seems to persist only in the European brown hare—H. concinna cycle at the studied habitat. H. concinna may not serve exclusively as an arthropod vector, but it may also harbor bacteria for 3–4 years through multiple life stages and act as an important reservoir of F. tularensis. Rodent species probably do not serve as true reservoir hosts of tularemia.
Introduction
Although F. tularensis is a potential biological warfare agent and several emergences or re-emergences of tularemia have recently been seen all over the world (Petersen and Schriefer 2005, Kaysser et al. 2008), the ecology of the disease is still only partially understood, with many open questions about reservoirs and vectors. The current knowledge is primarily based on investigations performed during disease outbreaks (Gurycová et al. 2001, Kaysser et al. 2008).
The aim of the current study was the direct and indirect detection of F. tularensis in potential animal reservoirs, domestic animals, arthropod vectors, and natural waters to obtain data about the ecological cycle of tularemia in an enzootic area during an inter-epizootic period. Grassland and woodland habitats situated in an enzootic lowland region were investigated in a-year-round study.
Materials and Methods
The study region was a 10-km2, large, lowland scene situated in southeastern Hungary (47°05′N, 20°56′E). Sodic steppes, grasslands, remnants of oak woods, marshlands, and small water courses compose the landscape. The climate is continental with cold, cloudy, humid winters (−3°C mean temperature in January 2009) and warm to hot summers (23°C mean temperature in July 2009). Retrospective data about the tularemia situation of the region were obtained from the local veterinary authority and human health service. Samples for this study were collected during a 1-year period, from December 2008 until December 2009.
European brown hares (Lepus europaeus) shot during the annual hunting events in December 2008 and 2009 were screened with both slide and tube agglutination tests (Treml et al. 2007, OIE 2008). Tests were performed according to the manufacturer's instructions with a diagnostic kit (Bioveta Inc., Ivanovice na Hané, Czech Republic) listed in the World Health Organization Guidelines (WHO 2007) utilizing inactivated bacteria. In brief, a drop of blood (∼0.04 mL) was mixed with five drops (∼0.2 mL) of antigen in the slide agglutination test, and the reaction was considered positive if flakes were created within 1–3 min (at 20°C–25°C). The tube agglutination test was performed with 0.5 mL aliquots of serial dilutions (from 1:10 to 1:160) of sera mixed with 0.5 mL of diluted (1:4) antigen. The test was considered positive if visible agglutination with clear supernatant fluid took place after 20 h of incubation at 37°C and 1 h at room temperature. According to the manufacturer's directions, agglutination at dilutions of 1:80 or higher should be considered a positive result, whereas at 1:40 it is still ambiguous. The positive control serum was provided by the manufacturer. Necropsies, histological and immunohistochemical examinations, the isolation, and identification of F. tularensis strains were carried out as described previously (Gyuranecz et al. 2010).
Small mammals were live-trapped during four sessions performed in April, July, October, and November 2009, in two study habitats: an open grassland and an oak-wood area. Sherman traps laid out in 7 × 7 grids with 10-m inter-trap intervals were used for five nights and days for live trapping. Ectoparasites (ticks and fleas) were combed off the rodents' coats, identified, and further examined individually as described below. The blood of small mammals was screened by slide agglutination test as described above. Lung, liver, spleen, and kidney tissue pools (a total of about 100 mg) from each individual were homogenized in 1000 μL Tris-EDTA (pH 8.0) (TissueLyser; Qiagen Inc., Hilden, Germany) and centrifuged, and a 100 μL supernatant from each sample was used for DNA extraction (X-tractor Gene; Corbett Robotics Pty. Ltd., Queensland, Australia) by the Total RNA Isolation Kit, Nucleospin 96 RNA (Macherey-Nagel GmbH & Co. KG, Düren, Germany), except for the DNase incubation step. DNA was eluted in 50 μL elution buffer.
A part of the 17 kDa major membrane protein (tul4) precursor gene was amplified using a real-time TaqMan polymerase chain reaction (PCR) system as described previously (Versage et al. 2003). According to the original description the detection limit of this assay is one colony forming unit (CFU), whereas the validation procedure during the adaptation of the test determined a 10 CFU detection threshold (Ct 38). A liver sample (about 50 mg) from an experimentally infected mouse (NMRI mouse, ∼20 g) served as the positive control both in DNA extraction and PCR.
Ticks were collected by dragging from the vegetation in the grassland and the oak-wood near the Sherman traps for 3–3 h in April, July, and October 2009. After identification, ticks were pooled (30 larvae, 15 nymphs, 10 or fewer males or females/pool), and homogenized in 500 μL Tris-EDTA (pH 8.0) buffer. DNA was extracted and analyzed by real-time PCR as described above. Positive samples were further characterized by the sequencing of a 400 bp large sequence of the tul4 gene amplified with a conventional PCR system as described earlier (Sjöstedt et al. 1997).
About 250-mL water samples were taken from all the existing water sources located in the study area in April, July, and October 2009. Samples were filtered on 0.2 μm membrane filters (Seitz, Bad Kreuznach, Germany). Discs of 10 mm in diameter were excised from each membrane and homogenized in 750 μL Tris-EDTA (pH 8.0) buffer. DNA was extracted and analyzed by the real-time PCR method as described above. An additional 250 mL water sample collected from the lake in April was mixed with around 100 CFU F. tularensis ssp. holarctica isolate and served as a positive control to check for potential PCR inhibition.
Blood samples were collected from sheep (jugular vein), cows (caudal vein), and buffalos (caudal vein) in May 2009. All animals belonged to extensively managed flocks of 100 sheep, and herds of 186 cattle and 92 buffalos grazed in the study area. Blood and sera were examined by slide and tube agglutination tests as described above.
Results
The study area is considered a tularemia endemic region as the infection has repeatedly been detected in 5%–7% of the local European brown hare population during the past 15 years (estimated from 3320 hares by slide agglutination test). In addition, 21 human clinical cases occurred in the surrounding towns (18,000 inhabitants) during the same period. The latest epizootics could be identified in 1997, 1999, and 2004, when seropositivities of 20.4%, 36.8%, and 12.3% were recorded in the local hare population.
A total of 197 hares were shot during the annual hunting events in December 2008 (75 hares) and 2009 (122 hares). The seroprevalence of tularemia was found to be 5.3% (4/75) in 2008 and 4.9% (6/122) in 2009 by the slide agglutination test. All animals seropositive by slide agglutination (10 animals) exhibited tube agglutination titers of at least 1/10, even when evaluated strictly by the criteria (agglutinated sediment and clear supernatant fluid) recommended in the Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (OIE 2008) (Table 1). The 1/40 cut off titer recommended by the manufacturer was identified in only two of these positive cases (one each year) and only a single animal reached the 1/80 antibody titer. The later is the positive threshold value recommended by the manufacturer of the diagnostic kit. Six of the 10 seropositive hares identified by slide agglutination test were necropsied, and tularemic lesions demonstrating in situ F. tularensis–specific immunolabeling (Gyuranecz et al. 2010) were identified in each individual. The most often affected organs were lungs and kidneys. F. tularensis ssp. holarctica was isolated from four of these six cases.
NE, not examined.
One hundred thirty-seven small mammals were trapped in the two study habitats during the year (Fig. 1). Further 40 animals, including 19 common voles, 8 yellow-necked mice (Apodemus flavicollis), 2 striped field mice (Apodemus agrarius), and a by-catch of 8 Eurasian pygmy shrews (Sorex minutus) and 3 common shrews (Sorex araneus), were trapped next to the oak wood in November. All these 177 small mammals were negative for F. tularensis infection with both slide agglutination test and real-time PCR. A total of 457 ectoparasite specimens were collected from the small mammals, comprising 404 Ixodes ricinus (362 larvae, 41 nymphs, and 1 female) and 28 Haemaphysalis concinna (27 larvae and 1 nymph) ticks and 15 Ctenophtalmus assimilis and 10 Nosopsyllus fasciatus fleas. All these ectoparasites tested negative for F. tularensis by real-time PCR.
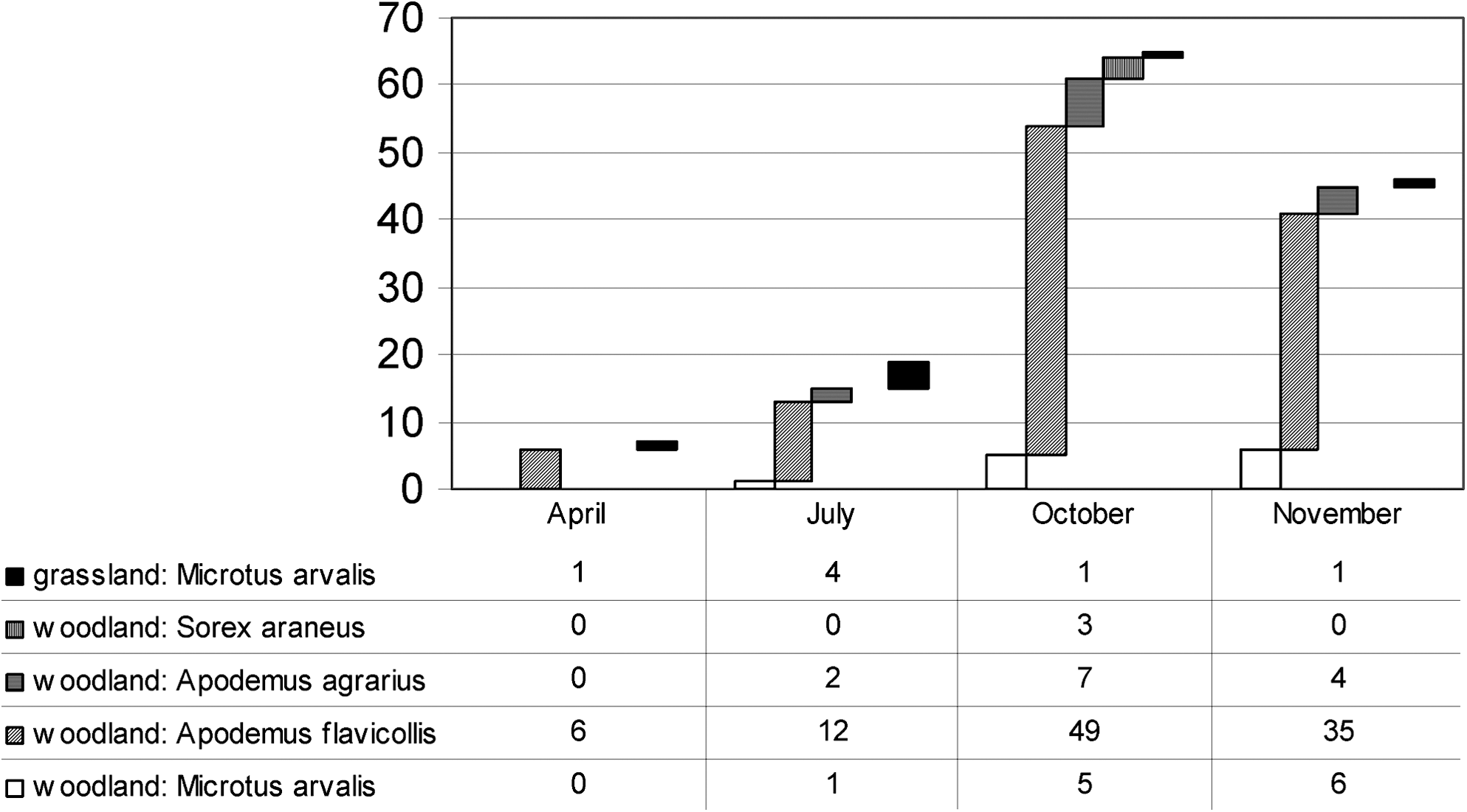
Number and distribution of small mammal species trapped in the grassland and in the oak-wood study habitat during the study year.
A total of 1582 I. ricinus and H. concinna ticks were collected from the vegetation in the grassland and in the oak-wood. Seventy percent of ticks were I. ricinus (1106) and 30% H. concinna (476). The two species exhibited different seasonal activity peaks during the year (Fig. 2). One H. concinna female pool of two individuals and one H. concinna nymph pool of 15 ticks collected at the grassland area in July were found positive for F. tularensis by real-time PCR. The 400-bp, large fragments of the tul4 gene were sequenced and submitted to GenBank (HM014008, HM014009). These findings mean that F. tularensis ssp. holarctica was carried by at least two H. concinna ticks, indicating a 0.42% (2/476) minimum prevalence for the whole region and 0.59% (2/336) for the grassland habitat.

Number and distribution of tick species and their development stages collected from the vegetation in the grassland and in the oak-wood habitat during the study year.
Water samples were taken from 6 ditches and one small lake located in the study area during April. In July and in October 2009 samples were obtained only from the lake as the ditches dried up by then. F. tularensis was not detected with real-time PCR in any of the 9 water samples. The Ct value was 35 for the positive control water sample.
Blood samples were collected from 100 sheep, 50 cows, and 50 buffalos grazed at the study area. The blood and sera of these 200 ruminants were also negative for F. tularensis antibodies with both the slide and tube agglutination (agglutination titers <1:10) tests.
Discussion
The result of our study, establishing 4.9%–5.3% prevalence of tularemia in hares, is in agreement with that of previous reports showing that the European brown hare is a common host of F. tularensis in Central Europe (Hopla 1974, Treml et al. 2007). The public health importance of the brown hare is accentuated by being one of the most abundant and most significant European game species (Strauss and Pohlmeyer 2001) and by the fact that direct contact with infected hares is a frequent mode of F. tularensis infection in humans (Mörner and Addison 2001). Hares are moderately sensitive to F. tularensis infection, they seroconvert, and potentially they can carry viable bacteria over a longer time span and thus serve as a reservoir species (Gyuranecz et al. 2010). Therefore, serological tests are useful diagnostic aids and tools for epidemiological surveys of tularemia in European brown hares. To reduce the possibility of introducing tularemia into new areas, the tube agglutination test is commonly used for screening of trapped hares before translocation for sporting purposes, reintroduction, and enhancement of existing populations. As shown in Table 1, the tube agglutination assay mainly demonstrated low antibody titers, and the diagnostic titers of 1/40 (ambiguous) and 1:80 recommended for this kit were reached only in two and a single case, respectively. Similarly, this kit also demonstrated low antibody titers (1/10 and 1/20) in 42% (29/69) of tularemia seropositive hares in a previous study (Treml et al. 2007). Since hares with lower titers showed typical pathological lesions associated with F. tularensis–specific immunolabeling, as well as clear-cut positivity by slide agglutination and were even culture positive for F. tularensis, we would recommend considering the modification of the diagnostic titer of this diagnostic kit to 1/10 for screening European brown hares before translocation. The slide agglutination test was found to be very useful and sensitive tool for the screening of tularemia in both live and dead hares as demonstrated by the present study and by our earlier long-term monitoring data (unpublished). However, since F. tularensis shares common antigens with Brucella spp., Yersinia spp., and Legionella spp., these pathogens may sometimes be responsible for false-positive results with this test (Gyuranecz et al. 2010). The frequent presence of lesions in the kidneys and lungs correlates with the results of a previous study (Gyuranecz et al. 2010) and lends further support to the hypothesis, that shedding of bacteria by urine contributes to the acquisition of tularemia by the airborne infection route in European brown hares.
According to previous studies, rodents play a key role in maintaining enzootic foci of tularemia in Eurasia (Friend 2006, Keim et al. 2007). The common vole and the European water vole (Arvicola amphibius) are the species most frequently involved in tularemia epizootics. In addition to being highly susceptible to tularemia, these species may also become chronically infected and thereby serve as disease reservoirs during periods between epizootics and they are also hosts for the immature stages of several important tick vectors (Bell and Stewart 1975, Friend 2006). The prevalence of infection observed during tularemia outbreaks was 4.5% (4/88), 5.2% (4/79), and 8.0% (2/25) by bacterial culture and real-time PCR among common voles in Slovakia, Austria (Gurycová et al. 2001), and Germany (Kaysser et al. 2008), respectively. In contrast to these results, we did not find any infected voles with either PCR or serology during the inter-epizootic period. Although the population of voles in our study region was low during the year and we managed to trap and sample only 38 individuals, the present data did not confirm the suggestion that common voles become chronically infected and thus serve as a reservoir species between outbreaks. Further, previous results demonstrating a complete lack of seroconversion among voles in spite of PCR positivity (Kaysser et al. 2008) indicate toward the dominance of fatal outcome in tularemia infections of this species. Mouse species (Mus musculus and Apodemus spp.) are highly susceptible and sensitive to tularemia; they die shortly after infection (Bandouchova et al. 2009) and are not known to be important reservoir hosts (Friend 2006). The population of Apodemus spp. at our study site was high with a normal population curve, and we did not detect any infected individuals, which is in congruence with the above finding.
As true reservoir hosts, ticks can carry F. tularensis for several years through multiple life-stages, which may help perpetuate specific enzootic foci of tularemia during inter-epizootic periods (Friend 2006, Keim et al. 2007). The seasonal activity of H. concinna and I. ricinus was similar to that observed earlier in Hungary (Széll et al. 2006, Hornok 2009). Dermacentor reticulatus is known to be the most frequent carrier of F. tularensis in Central European flood-plain forests and meadow ecosystems with infection rates of 0.5%–2.6% in Slovakia and Czech Republic (Gurycová et al. 1995, Hubálek et al. 1998, Gurycová et al. 2001) and 0.7%–5.8% in Austria (Gurycová et al. 2001), whereas the rates of infection in I. ricinus were 0.1%–0.2% (Gurycová et al. 1995) in forest habitats near the river Danube. D. reticulatus was not present at the site of our investigation, which could be explained by the arid habitat dominated by grasslands. Nevertheless, the expansion of this tick species and its preference for xerophylic vegetation was also observed in Hungary (Sréter et al. 2005, Hornok and Farkas 2009). The 0.59% infection rate found in H. concinna collected at the grassland habitat is lower than the prevalence registered in previous studies at forest habitats near the river Danube (2.8%, 1/35) (Gurycová et al. 1995). However, the sample size of the referred study was much smaller. H. concinna probably serves as the primary arthropod vector in the ecology of tularemia in our study region, and with its developmental cycle of three or more years it may also play an important role in the long-term persistence of enzootic foci of tularemia as previously described by Gurycová et al. (1995). Although the number of examined fleas was low, our results support earlier findings establishing that fleas should be considered of minor importance in the transmission and maintenance of F. tularensis (Parker and Johnson 1957, Keim et al. 2007).
There are several reports about water-borne tularemia infections in humans and the detection of F. tularensis in natural water bodies during outbreaks (Friend 2006, Kaysser et al. 2008). Although limited, the number of water samples examined in our study is representative of the water sources at our study site. The negative results could be explained by the lack of carcasses that would usually contaminate the water during epizootics, the lack of aquatic rodent species that may harbor and shed live bacteria into their environment, and the arid summer climate of the region allowing only temporary water bodies that regularly dry out every year. Alternatively, the negative PCR results could have also been obtained because the number of F. tularensis organisms in water samples was below the limit of detection (10 CFU/250 mL water).
Epizootiologic surveys of domestic animals, that is, relatively resistant species that survive the infection and develop antibodies, such as sheep or cattle, may indicate the epidemiological status of a given area (OIE 2008). Since they collect large numbers of ticks while grazing, tularemia infections and seropositive cases could be expected to occur regularly in endemic regions. Apart from the low prevalence of the disease in the area, another potential explanation for the absence of seropositive cases in our study could be the low infectious dose transmitted by ticks that may be insufficient to initiate seroconversion.
In our study we explored which species take part in the ecological cycle of tularemia in an enzootic area, serving as reservoirs and vectors of the disease between epizootics. According to our results we hypothesize that only a European brown hare–H. concinna cycle, confined to the grassland habitat, persists during inter-epizootic periods in our study region. It is potentially complemented by a hare–hare cycle as suggested by pathological findings. This means that hares may acquire infection by the air-borne route, become chronically infected, and shed live bacteria by urine into the environment. H. concinna does not only serve as an arthropod vector, but it can also carry the bacteria for several years through multiple life-stages (Gurycová et al. 1995) and may therefore be an important reservoir of F. tularensis. Various rodent species are probably not true reservoir hosts as they are all highly susceptible and sensitive to tularemia, dying shortly after infection, so positive cases can only be found during epizootics. The knowledge about the reservoirs of F. tularensis is crucial to assess potential human infection sources and to establish a systematic, long-term monitoring of tularemia activity in enzootic areas.
Footnotes
Acknowledgments
This work was funded by the OTKA-78139 grant and the NKB grant of the Faculty of Veterinary Science, Szent István University. G.F. is supported by the János Bolyai Research Scholarship of the Hungarian Academy of Sciences.
Disclosure Statement
No competing financial interests exist.