
Abstract
This report builds on recent serological evidence for the presence of hantavirus in northern Colombia by providing sequence-specific and phylogenetic data of hantavirus infections in wild rodents. From August 2007 to August 2008, 354 rodent specimens representing four families were collected in the northwestern Antioquia region of Colombia. Antibodies reactive to Sin Nombre virus and Maciel virus antigens by IgG enzyme-linked immunosorbent assay were found in 15 of 109 (14%) Cherries cane rats (Zygodontomys cherriei), the only sigmodontinae rodents captured. Lung tissue samples from 11 of the 15 seropositive rodents were RT–polymerase chain reaction positive for hantavirus RNA, using primers for the S and M genome segments. Eight of these amplicons were sequenced and phylogenetic analyses indicated RNA of a hantavirus closely related to Calabazo virus, previously found in Panama. This is the first report of the genetic characterization of a hantavirus in rodents in Colombia.
Introduction
In the spring of 1993, a previously unknown hantavirus was discovered in the Four Corners region of the United States. The virus was associated with an outbreak of human respiratory disease with a case fatality ratio of 52% (CDC 1994, Khan et al. 1996). This new agent was named Sin Nombre virus (SNV), and deer mice (Peromyscus maniculatus) were identified as its principal rodent reservoir host (Childs et al. 1994).
Since then, many pathogenic hantaviruses and their rodent reservoirs identified have been reported from Panama to the southern cone of South America: Choclo virus (Oryzomys fulvescens) (Salazar-Bravo et al. 2004), Caño Delgadito virus (Sigmodon alstoni) (Fulhorst et al. 1997), Castelo dos Sonhos virus (unidentified reservoir) (Johnson et al. 1999), Araraquara virus (Bolomys lasiurus) (Johnson et al. 1999), Juquitiba virus (Oryzomys nigripes) (Suzuki et al. 2004), Laguna Negra virus (Calomys laucha) (Johnson et al. 1997), Orán virus (Oligoryzomys chacoensis) (Calderon et al. 1999), Bermejo virus (Oligoryzomys chacoensis) (Padula et al. 2002), Central Plata virus (Oligoryzomys flavescens) (Delfraro et al. 2003), Lechiguanas virus (Oligoryzomys flavescens) (Levis et al. 1997), and Andes virus (Oligoryzomys longicaudatus) (Levis et al. 1998, Delfraro et al. 2003). Likewise, nonpathogenic hantaviruses have also been discovered: Calabazo virus (Zygodontomys brevicauda cherriei) (Salazar-Bravo et al. 2004), Maporal virus (Oecomys bicolor and Oligoryzomys fulvescens) (Milazzo et al. 2002, Fulhorst et al. 2004), Anajatuba virus (Oligoryzomys fornesi), Rio Mearim virus (Holochilus sciureus) (Rosa et al. 2005), Rio Mamore virus (Oryzomys microtis) (Bharadwaj et al. 1997), Maciel virus (MCLV) (Necromys benefactus), and Pergamino virus (Akodon azarae) (Levis et al. 1998).
From 1993 to the present, there have been several outbreaks of HPS in Argentina (Lopez et al. 1996), Bolivia (Johnson et al. 1997), Brazil (da Silva et al. 1997), Chile (Toro et al. 1998), Paraguay (Williams et al. 1997), Uruguay (Padula et al. 2000), and Panama (Vincent et al. 2000). Only serological evidence of hantavirus infection in both rodents and humans has been reported in Colombia (Mattar and Parra 2004, Aleman et al. 2006). Consequently, our goal was to obtain genetic evidence of a hantavirus in rodents captured at three municipalities of the Uraba Gulf (Antioquia, Colombia), close to the Panama border. We found a 4% (15/354) seroprevalence of rodent infection (by enzyme-linked immunosorbent assay [ELISA]) and the first genetic evidence for the presence of a hantavirus in Colombia.
Materials and Methods
Study sites
Rodents were captured in or near the municipalities of Apartado, Turbo, and Necocli. These towns are located on the Atlantic coast in the Uraba Gulf near the southwestern most corner of the Caribbean Sea, between 330 and 430 km northwest of Medellin, which is the capital of the Department (Province) of Antioquia, Colombia (Fig. 1). Within the trapping area, the climate is hot and humid, with an average temperature of 28°C and relative humidity of 85%. The economy of the region is mainly agricultural, with bananas as the main product, followed by plantain, cassava, corn, and rice. Other commercial activities in the area are fisheries, livestock breeding and production, forestry, and tourism.
Maps of Colombia (left) and the Department of Antioquia (right), highlighting the three municipalities where the rodent captures were performed.
Rodent sampling
Between August 2007 and August 2008, we performed monthly week-long live-trapping with an additional week of trapping in March 2009. We used 50 Sherman traps (HB Sherman Trap Company) and 16 Tomahawk traps (Tomahawk Trap Company) for the capture of living animals.
Traps were set at dusk in both urban (mainly grain and food storage structures) and rural (corn, rice, and cassava plantations and gallery forest near crops) areas and baited with a mixture of peanut butter, oat flakes, and vanilla essence. The traps were checked the next morning and the captured rodents were collected and processed. Trapping and sampling of rodents were carried out following standard safety procedures (Mills 1998). Briefly, captured rodents were anesthetized intramuscularly with a 9:2 mixture of Ketamine (Imalgene®; Merial) and Xylazine (Rompun®; Bayer) using 0.1 mL per 50 g of body weight. The anesthetized rodents were bled by cardiac puncture or from the retrobulbar plexus using heparinized capillary tubes. Blood samples were collected in glass tubes without anticoagulant, and sera were separated and transported under refrigeration. Rodents were euthanized by an overdose of sodium pentobarbital (Euthanex®; Invet) and body measurements were recorded for identification. Lung, liver, kidney, and spleen tissues were placed in plastic vials and immediately stored in liquid nitrogen. At the end of the field work, serum samples were transported to the laboratory and kept at −20°C and the tissue samples were frozen at −80°C. This study was conducted with the approval of the animal research ethics committee of the Universidad de Antioquia and also of Corpouraba, the environmental authority of the region.
Identification of rodents
Rodents were identified based on morphometric measurements, the characterization of discrete features of skulls (presence of crests, processes and face appearance, teeth size, etc.) and by comparison of skins with others previously classified and available at the Mastozoology Laboratory of the Universidad de Antioquia. Samples (embalmed skin and skulls, and some formalized tissues) of all the rodents captured were deposited into the Biology Museum of the University, available for further characterization. Currently, genetic identification of these rodent species is also underway, to settle possible discrepancies with previous descriptions in the literature regarding the classification of the captured specimens (not shown).
Serological tests
Blood samples from rodents were tested by ELISA for IgG antibodies reactive with nucleocapsid antigen of SNV (Schountz et al. 2007) and antibodies reactive to whole inactivated MCLV (Instituto Nacional de Enfermedades Virales Humanas—INEVH, Pergamino, Argentina). For testing with MCLV antigens, we followed the basis of a previous protocol (Ksiazek et al. 1995), with modifications. Briefly, the wells of a 96-well plate (Thermo Fisher Scientific) were prepared by coating (i) the upper half with 100 μL of a clarified homogenate of Vero cells infected with the MCLV and (ii) the lower half with 100 μL of the homogenate of uninfected Vero cells. The plate was incubated overnight at 4°C and the wells washed five times with washing buffer (phosphate-buffered saline with 0.1% Tween-20). Serum samples, one positive and four negative controls (negative controls were obtained from Balb/c mice; the positive control was obtained from a pool of wild rodents acquired from the INEVH), were diluted (1:100, 1:400, 1:1600, and 1:6400) in master solution (phosphate-buffered saline [pH 7.4], 5% skim milk, and 0.5% Tween 20), and 100 μL of each of these dilutions was incubated with both positive and negative antigens at 37°C for 1 h. After five additional washes, 100 μL of the detecting conjugated antibody (a mixture of anti-mouse and anti-Peromyscus leucopus, white-footed mouse; KPL) was added and incubated for 1 h at 37°C. The plate was then washed five times, and 100 μL of tetra methyl benzidine (TMB) substrate was added to each well. After 20 min of incubation at room temperature, 100 μL of the stop solution (1 M sulfuric acid) was added and the optical densities were read with two wavelength filters within the range of 450 and 630 nm.
RT–polymerase chain reaction and sequence analysis
Total viral RNA was extracted from seropositive rodent lung tissue samples using TRIzol (Invitrogen). A 513-nt fragment of M genome segment and a 183-nt fragment of the S genome segment were amplified by nested RT–polymerase chain reaction (PCR) following the protocol previously described by Levis et al. (1998) for sequence analysis purposes (sequences for the M segment primers for Argentinean hantaviruses were provided by Dr. Levis, INEVH, Pergamino, Argentina, and would be available upon request; sequences for the S segment primers have not been published and were generously provided by Institute Carlos III, Madrid, Spain; for research purposes please make specific requests to Dr. Antonio Tenorio).
DNA products were purified and sequenced by a commercial facility (Macrogene). The resulting sequences were aligned with hantavirus sequences available in GenBank using Clustal X (Thompson et al. 1997). The nucleotide and amino acid sequences of the nuclear (N) protein and glycoprotein precursor genes obtained were compared with homologous sequences of other New World and Old World hantaviruses (Table 1). Aligned sequences were used to determine sequence identities and phylogenetic relationships. Identities were calculated by subtracting uncorrected (p) genetic distances from 1.0. Genetic distances were obtained with the showdist command in the Paup* program (Swofford 2002). Phylogenetic analysis was done using the Bayesian method implemented in MrBayes v3.1.2 software package (Ronquist and Huelsenbeck 2003). The substitution model used was the general time-reversible with gamma distribution of among-site rate variation. Analyses were run until good convergence of the two Markov chains Monte Carlo searches was reached, as assessed by an average standard deviation in partition frequency lower than 0.01.
Accession numbers for the sequences used to design the phylogenetic tree.
Results
Rodent capture
Three hundred fifty-four rodents were collected from domestic areas associated with urban and rural settings in the municipalities of Apartado (8 specimens), Turbo (109 specimens), and Necocli (237 specimens) (Table 2). Necocli was the region with the greatest habitat diversity and provided the widest range of species.
From 104 Zygodontomys cherriei from Necocli, 15 were positive for hantavirus IgG detection by ELISA and 11 were positive by RT–polymerase chain reaction.
Of the rodents captured, 219 included individuals from the subfamily: (i) Murinae: Rattus rattus (124 specimens), Rattus norvegicus (24 specimens), and Mus musculus (71 specimens); (ii) Sigmodontinae—Zygodontomys cherriei, Cherries cane rat (Voss 1991, Weksler 2006) (109 specimens); (iii) Echimyidae—Proechimys semispinosus, Spiny rat (Patton 1987) (22 specimens); and (iv) Heteromyidae—Heteromys anomalus, Spiny pocket mouse (Anderson 1999) (4 specimens).
Serological analysis
Of the 354 blood samples, 15 (4%) had antibody to both SNV and MCLV antigens by ELISA. All antibody-positive rodents captured at the municipality of Necocli belonged to the species classified as Zygodontomys cherriei (15/109, 14%).
Molecular and sequence analyses
Lung tissue was removed from the 15 seropositive rodents and examined for viral genetic material using a nested RT–PCR approach. Eleven of these samples were found to be positive, being able to support the amplification of 183 nt and 513 nt bands using S and M segment primers, respectively (Fig. 2).
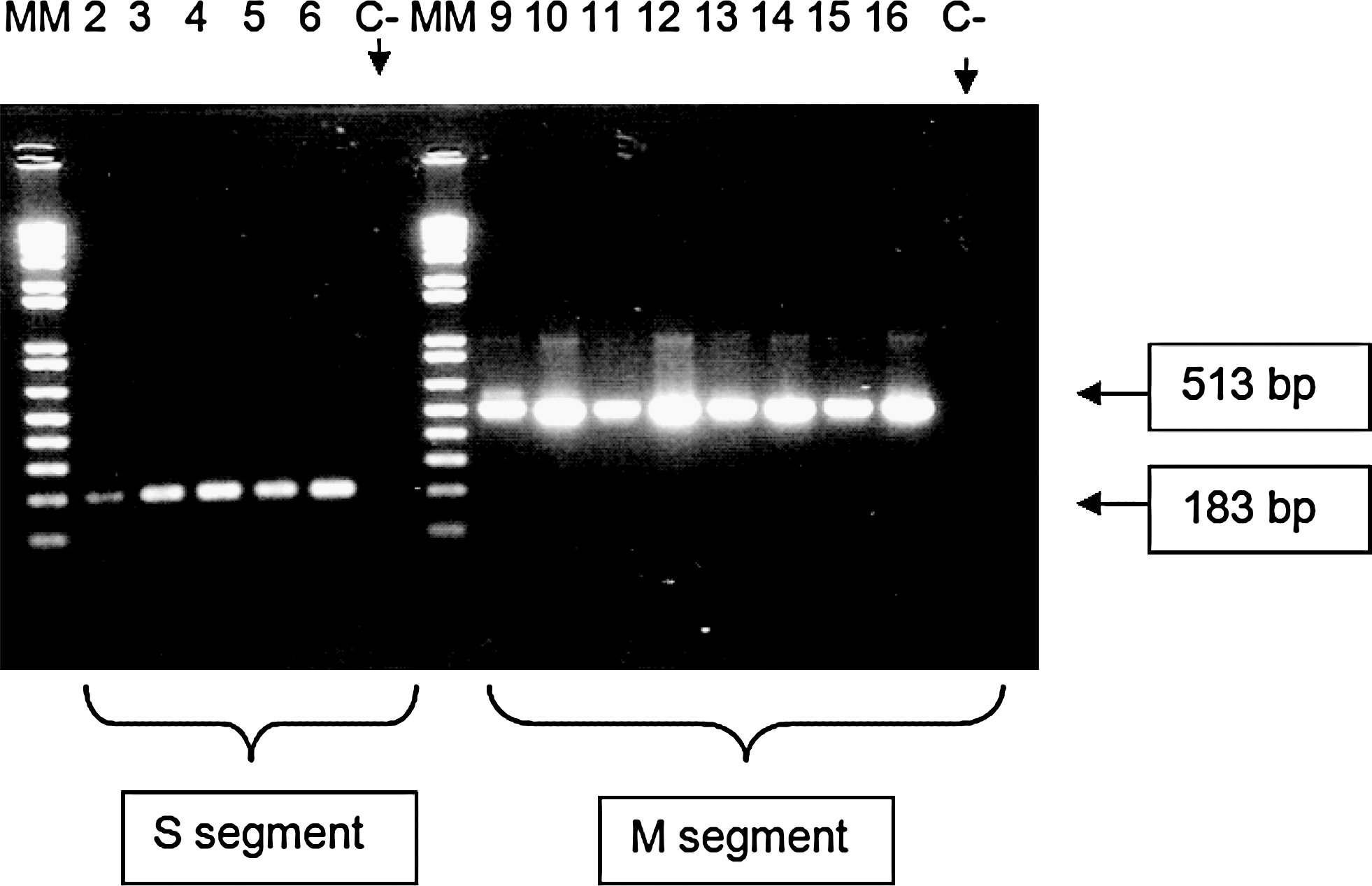
RT–polymerase chain reaction electrophoresis. Rodent DNA lung samples run on an agarose gel after RT–polymerase chain reaction. Samples in lanes 2–6 on the left side were amplified by S segment primers (unpublished sequences generously provided by Dr. Antonio Tenorio, Institute Carlos III, Spain). The same samples are seen on the right side in lanes 9–13 but amplified by primers for the M segment. Lanes 6 and 13 correspond to rodent lung sample 232, which was positive for hantavirus antibody by enzyme-linked immunosorbent assay at a dilution of 1:100 but negative at 1:400.
Sequencing generated a 476-nt fragment from positions 2726–3193 of the M segment, enabling comparisons with six seropositive rodents from Necocli. Over this region, there were only between 0 and 2 nucleotide differences among them (99%–100% identities). They were, therefore, considered to be of the same species. We have provisionally termed this Necocli M segment sequence. Identities with other American hantavirus sequences obtained from M segments published in GenBank ranged between 70% and 80% in this genomic region.
Strong sequence identities were noted between the Necocli M segment sequence and Calabazo virus in a 202-nt fragment of the G2 encoding region obtained from GenBank. To examine this relationship in more detail, we sequenced a 376-nt fragment of the S segment of one of our Necocli samples (Necocli 230) and compared it with Calabazo and other homologous hantavirus sequences (Table 3). Identities of 85% and 100% at the nucleotide and amino acid levels, respectively, were found between Necocli and Calabazo viruses.
NECV, Necocli; CALV, Calabazo; ANDV, Andes; ARAV, Araraquara; LECV, Lechiguanas; MCLV, Maciel; JUQV, Juquitiba; MPLV, Maporal; CHLV, Choclo; LNV, Laguna Negra; RMV, Rio Mamore; CATV, Catacamas; RIOSV, Rio Segundo; SNV, Sin Nombre; CDV, Cano Delgadito; PUUV, Puumala; DOBV, Dobrava; SEOUV, Seoul; HTNV, Hantaan viruses.
Phylogenetic analyses of the concatenated S and M fragments (totaling 578 nucleotides), using Old World hantaviruses as an outgroup, segregated North American and South American hantaviruses in separate monophyletic groups. Caño Delgadito virus was an exception as it did not cluster with any other virus (Fig. 3). Calabazo and Choclo viruses isolated in Panama clustered in the South American group. The sequences from Necocli and Calabazo viruses were monophyletic with a posterior probability value of 1.00. Both viruses also formed a clade with Maporal virus with a 0.91 posterior probability.
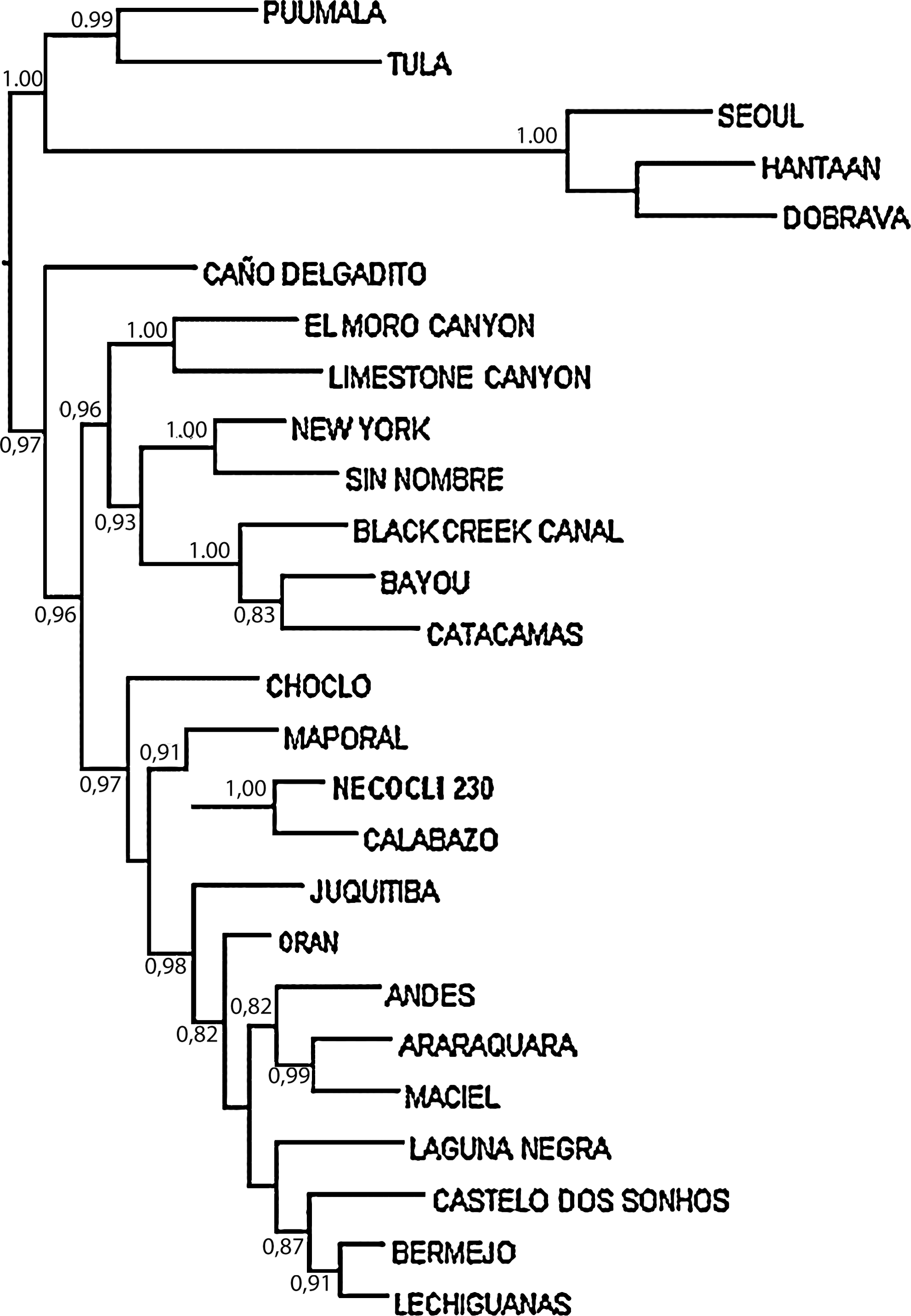
Majority-rule consensus tree generated in the Bayesian phylogenetic analysis performed with the program MrBayes v3.1.2., using the GTR+Γ substitution model. The analysis was run for 500,000 generations. Sequence analyzed is comprised of nucleotide positions 313–688 and 2792–2993 in the S and M segments, respectively. The numbers near the nodes are posterior probabilities for the corresponding clade. Only probabilities higher than 80% are shown. The name of the sequence obtained in this study is shown in bold. The tree was rooted with Old World hantavirus sequences (Hantaan, Seoul, Dobrava, Puumala, and Tula viruses) and was drawn with FigTree v1.2.2.
Discussion
Since 1993, when SNV was first identified as a pathogenic hantavirus causing HPS in North America, similar viruses have been identified in other rodents of the Sigmodontinae subfamily of South and Central America. Some of these are also associated with HPS (Mills and Childs 1998). More than 10 years later, the first serological study of hantaviral circulation in human populations in Colombia showed 13.5% prevalence in samples from healthy rural workers (Mattar and Parra 2004). This study now provides the first genetic evidence of a hantavirus in Colombia.
In 2006, the first serological evidence of hantavirus infections in rodents from the Colombian Atlantic coast was published by Aleman 2006 and detailed a 2.1% seroprevalence in specimens of Oligoryzomys spp., Heteromys spp., and Proechimys spp. Our study provides additional information on the potential hantavirus reservoirs in a neighboring northern area of Colombia. The differences between the frequencies of seropositivities in both studies (2% vs. 4%) could be due, at least in part, to the higher number of Sigmodontinae rodents captured in the latest work (39% of the total in our case compared with 20% in Aleman's results). Differences between these studies may also be found in reactivity of rodents other than those of the Sigmodontinae subfamily in previous reports. However, in addition to the serologic evidence, our study provides the first genetic evidence of hantaviral infection in rodents from Colombia. Accordingly, sequence analysis shows the S and M segments of this hantavirus to be very closely related to Calabazo virus. Taken all together, the serological and molecular evidence presented here suggests that Cherries cane rats (Zygodontomys cherriei) are the most likely reservoir for this Colombian hantavirus. Identification of Z. cherriei was determined from a review by Weksler (2006). This species was known in the past as Zygodontomys bravicauda subspecies cherrei.
Fourteen of 15 positive Cherries cane rats captured were male, suggesting a greater gender predisposition that could be explained by territorial fights (Calisher et al. 2007). As mentioned previously, there is evidence that males might be more susceptible to hantaviruses than females, possibly because of deficiencies in the innate immune response mediated by hormonal influx (Hannah et al. 2008).
The phylogenetic tree (Fig. 3) and the relatively low nucleotide identity (<85%) of the sequences of the virus found in rodents from Necocli with other members of the genus suggest that this might be a new hantavirus for Colombia. The closest relative (with 100% identity at the amino acid level) is Calabazo, a hantavirus found in what appears to be the same rodent species in the Azuero Peninsula of Panama (Vincent et al. 2000, Salazar-Bravo et al. 2004), about 414 miles west of Necocli. The close relationship between these two viruses could indicate that Necocli might just be a strain of Calabazo, but still it will be the first hantavirus identified in the Colombian territory. As Calabazo and Necocli viruses cluster with Maporal and other South American hantaviruses it is likely that their most recent common ancestor was a South American hantavirus; a more detailed analysis, including sequences from the three genomic segments, is warranted. Although Calabazo virus has not been associated with human infection or disease, we cannot make inferences about the susceptibility of humans to Necocli virus. However, we speculate that it is likely that antibodies found by Mattar and Parra (2004) in farmers of the Cordoba Province of Colombia, <90 miles east of Necocli, were induced by Necocli virus or a closely related hantavirus.
The greatest number of rodent captures in the municipality of Necocli can be explained by the wide variety of habitats in this location. It was for this reason that we focused the last six visits of our work in this area.
Hantaviruses seem ubiquitous throughout South America. In Venezuela, there are reports of 1.7% seroprevalence among 1380 healthy individuals distributed around the country including one case of HPS in 1999 (Rivas et al. 2003) (although not confirmed to be associated with hantavirus). Other studies have demonstrated the circulation of Caño Delgadito and Maporal, although etiologic evidence of these viruses being associated with disease has not been shown (Fulhorst et al. 1997, Milazzo et al. 2002). In Peru, the HTN-007 circulates in Small-eared Colilargo (or small-eared pygmy rice rat, Oligoryzomys microtis) (Powers et al. 1999), and until now, there are no reports of hantaviruses from Ecuador. This could be misunderstood as indicating that hantaviruses do not pose the same extent of public health problems in northern South America, as in the south. More likely this is due to underreporting of cases due to insufficient epidemiological surveillance or inadequate diagnostic tools. Finally, the manifestations of hantavirus disease could be clinically confused with other compatible syndromes more commonly expected in Colombia, such as dengue, leptospirosis, or rickettsial diseases (Padmanabha et al. 2009); we are currently studying this possibility too. A different scenario occurs in certain countries in more southern areas of South America (Argentina, Uruguay, Paraguay, and Chile) and the center of South America (Brazil and Bolivia), where active surveillance programs are regularly carried out, and some of the ecoepidemiology of the reservoirs existing in each zone has been defined, so that it is easier to detect the emergence of a new hantavirus.
In the near future, what remains to be done with the Necocli virus is a more complete characterization, trying to determine its geographical distribution and its capacity to infect humans and produce disease. Additionally, we consider it important to perform similar studies in other regions of Colombia (central savanna, Amazonian rainforest, etc.), to establish a regular network of cooperation and collaboration.
Footnotes
Acknowledgments
The authors thank Dr. Tony Schountz, University of Northern Colorado, Greeley, CO, for his generosity in providing the SNV antigen used in our ELISA tests. The authors also thank Dr. Sergio Solari (professor of the Instituto de Biologia) and Daisy Gomez (biologist), both from the Mastozoology group at the Universidad de Antioquia, for their assistance in identifying the rodents. The authors are indebted to Ms. Sair Arboleda for drawing the maps in ![]() . On a more personal manner, the authors express profound gratitude to Dr. Maria Salvato (professor at the Institute of Human Virology, University of Maryland at Baltimore, MD), Dr. Charles Calisher (emeritus professor, Colorado State University, Fort Collins, CO), and Dr. Roger Hewson (Scientific Program Lead, Virology and Emerging Diseases, Health Protection Agency–Microbiology Services, Porton Down, Salisbury, United Kingdom), for their valuable insights, and editorial advice in the preparation of this manuscript. Finally, the authors are indebted to Colciencias for its financial support through grant no. 111534319203 (“Search for serological and genetic evidence of emergent and reemergent agents in humans and urban and rural rodents in Antioquia, Colombia”).
. On a more personal manner, the authors express profound gratitude to Dr. Maria Salvato (professor at the Institute of Human Virology, University of Maryland at Baltimore, MD), Dr. Charles Calisher (emeritus professor, Colorado State University, Fort Collins, CO), and Dr. Roger Hewson (Scientific Program Lead, Virology and Emerging Diseases, Health Protection Agency–Microbiology Services, Porton Down, Salisbury, United Kingdom), for their valuable insights, and editorial advice in the preparation of this manuscript. Finally, the authors are indebted to Colciencias for its financial support through grant no. 111534319203 (“Search for serological and genetic evidence of emergent and reemergent agents in humans and urban and rural rodents in Antioquia, Colombia”).
Disclosure Statement
The authors declare that no competing financial interests exist.